
Abstract
Recombinant adeno-associated virus (rAAV) vectors appear, more than ever, to be efficient viral vectors for in vivo gene transfer as illustrated by the approvals of 7 drugs across Europe and the United States. Nevertheless, preexisting immunity to AAV capsid in humans remains one of the major limits for a successful clinical translation. Whereas a preexisting humoral response to AAV capsid is well documented, the prevalence of preexisting capsid-specific T cell responses still needs to be studied and characterized. In this study, we investigated the prevalence of AAV-specific circulating T cells toward AAV2, 4, 5, 8, 9, and rh10 in a large cohort of healthy donors using the standard IFNγ ELISpot assay. We observed the highest prevalence of preexisting cellular immunity to AAV9 serotype followed by AAV8, AAV4, AAV2, AAVrh10, and AAV5 independently of the donors’ serological status. An in-depth analysis of T cell responses toward the 2 most prevalent serotypes 8 and 9 shows that IFNγ secretion is mainly mediated by CD8 T cells for both serotypes. A polyfunctional analysis reveals different cytokine profiles between AAV8 and AAV9. Surprisingly, no IL-2 secretion was mediated by anti-AAV9 immune cells suggesting that these cells may rather be exhausted or terminally differentiated than cytotoxic T cells. Altogether, these results suggest that preexisting immunity to AAV may vary depending on the serotype and support the necessity of using multiparametric monitoring methods to better characterize anticapsid cellular immunity and foresee its impact in rAAV-mediated clinical trials.
INTRODUCTION
Recombinant adeno-associated viral vectors (rAAV)-mediated gene therapy shows promising benefits for the treatment of genetic disorders, as illustrated by Glybera®, Luxturna®, Zolgensma®, Upstaza®, Roctavian®, Hemgenix®, and Elevidys® FDA and EMA approvals and more than 230 clinical trials ongoing ( 1 and clinicaltrials.gov). However, there are still major hurdles to overcome for their successful clinical translation such as their immunogenicity in patients. Indeed, since protocols evolved from local to systemic delivery with high doses of vector, adverse events related to the immune system activation have been reported in treated patients, resulting in some cases in regulatory holds of clinical trials. 2
Seroprevalence studies showed that human population is exposed to wild-type (WT) AAV virus early in life 3 and might present a memory immune response that can be reactivated after gene transfer. These studies were mainly conducted on human serum samples to evidence the presence of anti-AAV neutralizing antibodies 4,5 that will prevent target cell transduction after vector administration but little is known about the anti-AAV memory T cell response prevalence in humans. Importantly, these preexisting anti-AAV memory T cells might be reactivated after gene transfer and lead to gene transfer failure as previously described in a clinical trial targeting hemophilia B patients. 6,7 In this trial, the intrahepatic injection of rAAV2 led to the destruction of liver transduced cells 4–8 weeks after vector delivery due to the reactivation of preexisting IFNγ-secreting AAV-specific CD8 T cells. Since then, the liver transaminase elevation due to hepatotoxicity and the IFNγ secretion in response to AAV capsid are systematically measured in clinical trials and managed by corticosteroid treatment. 8 –10
To the best of our knowledge, in the general population, only cellular responses directed to AAV2, AAV1, AAV5, and AAV8 11 –13 have been investigated so far, which does not cover the whole diversity of rAAV serotypes now used in clinical trials. Indeed, no cellular prevalence data in humans are reported for the AAV9 capsid, which is used in a commercialized product since 2019 for the treatment of spinal muscular atrophy (SMA) in pediatric patients. 10 AAV9 serotype is also used in clinical trials targeting Pompe (NCT02240407), Canavan (NCT04998396), GM1 gangliosidosis (NCT03952637), and Parkinson (NCT04127578) diseases, as well as Duchenne muscular dystrophy (DMD) (NCT04240314, NCT03368742, and NCT03362502). The AAVrh10 has also emerged lately as a good candidate for gene delivery to the central nervous system and showed promising results for clinical applications targeting Krabbe disease, 14 metachromatic leukodystrophy 15 (and NCT01801709), GM1 gangliosidosis 16 (and NCT04273269), or amyotrophic lateral sclerosis (ALS). 8,17
In this study, we investigated the prevalence of AAV-specific circulating T cells toward AAV2, 4, 5, 8, 9, and rh10 in a large cohort of healthy donors using an IFNγ ELISpot assay that is usually used in immune monitoring in clinical studies. We observed the highest prevalence of preexisting cellular immunity against the AAV9 serotype followed by AAV8, AAV4, AAV2, AAVrh10, and AAV5. As already described, there was no correlation in our cohort between the cellular immune response and the humoral anti-capsid immunity. An in-depth analysis of T cell responses toward the two most prevalent serotypes 8 and 9 shows that IFNγ secretion is mainly mediated by CD8 T cells for both serotypes. We also performed FluoroSpot assays that showed their potential as a polyfunctional assay to characterize anti-AAV T cell responses to be used in clinical trials as we observed different cytokine profiles between AAV8 and AAV9. This observation suggests a different functional impact of preexisting capsid-specific T cells in rAAV-mediated clinical trials.
Our data suggest that preexisting anti-AAV cellular immune responses are serotype-dependent and encourage the use of multiparametric monitoring methods to better characterize capsid-specific cellular immune responses and predict their impact in rAAV-mediated clinical trials.
MATERIALS AND METHODS
All this work was performed at INSERM TaRGeT UM1089, Nantes, France, under the control of our quality management system that is approved by Lloyd’s Register Quality Assurance LRQA to meet the requirements of international Management System Standards ISO 9001:2015. It has been implemented to cover all activities in the laboratory, including research experiments and production of research-grade viral vectors.
Healthy donor blood samples
Cytapheresis samples were provided by the local Etablissement Français du Sang (EFS Nantes, Pays de la Loire, agreements N° PLER NTS 2016-25 and CPDL-PLER-2021 005) and originated from consenting healthy donors living in the Pays de la Loire area, France (22 women and 129 men, with age ranging from 19 to 72 years). All the participants provided their written informed consent. Peripheral blood mononuclear cells (PBMCs) were isolated using Ficoll density gradient centrifugation (Ficoll-Paque PLUS, GE Healthcare) and frozen in liquid nitrogen for ELISpot and FluoroSpot assays. Plasma was isolated by spinning 10 min at 2000g, room temperature, 10 mL of whole blood, and was stored at −20°C.
IFNγ ELISpot assay on healthy donor PBMCs
Anti-AAV cellular immune responses were analyzed using an IFNγ ELISpot assay. ELISpot assays were performed according to the manufacturer’s recommendations [ELISpot Plus: Human IFN-y (ALP) kit, MABTech]. Briefly, PBMCs from healthy donors were stimulated in vitro for 18–48 h, with overlapping peptides spanning the VP1 capsid protein sequence (which also covers VP2 and VP3) of each serotype (i.e., AAV2, 4, 5, 8, 9, and rh10; NCBI accession numbers AAC03780, AAC58045.1, AAD13756.1, AAN03857.1, AAS99264.1, and AAO88201.1, respectively) and divided into 3 pools (15-mers overlapping by 10 aa, Sigma-Aldrich, United States, or Mimotopes, Australia). For each serotype, 143 (AAV5), 145 (AAV2 and 4), or 146 (AAV8, 9 and rh10) peptides were synthesized. Pool 1 was made of peptides 1–46 (AAV2), 1–48 (AAV5), or 1–49 (AAV4, 8, 9, and rh10). Pool 2 was made of peptides 47–96 (AAV2), 49–96 (AAV5), or 50–98 (AAV4, 8, and 9) or 50–97 (AAVrh10). Pool 3 was made of peptides 97–145 (AAV2), 97–143 (AAV5), 99–146 (AAV4, 8, and 9), or 98–146 (AAVrh10). A negative control consisted of unstimulated cells (medium only), whereas concanavalin A (Con A, 10 µg/mL, Sigma-Aldrich, United States) stimulation was used as a positive control for cytokine secretion. Spot number was determined using an ELISpot iSpot Spectrum reader (AID, Strassberg, Germany) and a Mabtech IRIS reader and analyzed with AID ELISpot reader software V7.0 (AID, Strassberg, Germany) and Mabtech APEX v1.1 software (Mabtech, Sweden), respectively. Responses were considered positive when the number of spot-forming colonies (SFCs) per 1e6 cells was >50 and at least three-fold higher than the negative control condition.
AAV vector productions
For enzyme-linked immunosorbent assay (ELISA), single-stranded AAV serotypes 2, 8, 9, and rh10 vectors were produced by the Center for Production of Vectors (CPV-vector core from University Hospital of Nantes/French Institute of Health [INSERM], University of Nantes [https://umr1089.univ-nantes.fr/en/facilities-cores/cpv]). Briefly, vectors were produced by cotransfection of human embryonic kidney (HEK) 293 cells with the vector plasmid and helper plasmid (containing helper genes from adenovirus and the rep and cap genes according to the capsid serotype). Vectors were purified by the cesium-chloride gradient. The enriched-empty (AAV2, 9, and rh10) or enriched-full (AAV8) particle bands were collected and dialyzed. The amount of viral capsid proteins for the AAV2, AAV9, and AAVrh10 vector batches was compared with an AAV2 standard by quantitative SDS-PAGE Coomassie blue staining. An ELISA total capsid titration was performed for AAV8 (kit from Progen, Ref. #PRAAV8).
Anti-AAV IgG ELISA
Detection of anti-AAV total IgG antibodies for each serotype in the plasma of healthy human donors was conducted using an ELISA. Nunc MaxiSorp P96 plates (Sigma-Aldrich) were coated overnight at 4°C with rAAV2, rAAV8, rAAV9, or rAAVrh10 particles (INSERM UMR 1089-Center for Production of Vectors). Vectors were produced, purified, and titrated as described in the previous section. After washing and plate saturation, wells were then incubated for 1 hour at 37°C with at least 6 serial four-fold dilutions of human plasma starting at 1:10 followed by the HRP-conjugated anti-human F(ab’)2 IgG antibody (Jackson) for 1 hour at 37°C. Revelation was performed using 3,3′-5,5′-tetramethylbenzidine (TMB, OptEIA, BD Biosciences) and absorbance of duplicate samples was read at 450 nm with a correction at 570 nm on a Multiskan Go reader (Thermo Scientific). Positive threshold curves for each ELISA were determined from seronegative human sera (>20 donors) as the mean optic density (O.D.) for each dilution + 2*SD. For each donor, anti-AAV IgG titer was defined as the last plasma dilution with an O.D. remaining above the threshold curve.
IFNγ ELISpot assay on healthy donor CD4- or CD8-depleted PBMCs
CD4+ or CD8+ T cells were depleted from total PBMCs using the Stem Cell magnetic-based system (EasySepTM Human CD4/CD8 Positive Selection Kit II) according to the manufacturer’s recommendations. Cell depletion efficiency was determined by flow cytometry before (total PBMCs) and after magnetic separation (CD4- and CD8-depleted PBMCs). Briefly, CD4+ T cell and CD8+ T cell populations were analyzed using the CD3 (557749; BD Biosciences), CD19 (561031; BD Biosciences), CD4 (560251; BD Biosciences), and CD8 (93-0088-42; eBioscience) markers. CD4 and CD8 T cells were gated as CD4+ or CD8+ cells, respectively, within the CD3+CD19- population. Cells were acquired using a BD FACS LSRII flow cytometer (BD Biosciences) and analyzed with FlowJoTM software (v.10; BD Life Sciences). Anti-AAV cellular responses were assessed using the IFNγ ELISpot assay as described above.
IFNγ/IL-2/TNFα FluoroSpot assay on healthy donor PBMCs
Anti-AAV cellular immune responses were also evaluated using IFNγ/IL-2/TNFα FluoroSpot assays. FluoroSpot assays were performed according to the manufacturer’s recommendations (FluoroSpot Plus: Human IFNγ/IL-2/TNFα kit, Mabtech). Briefly, PBMCs from healthy donors were stimulated in vitro for 48 h with the same AAV8 and AAV9 peptide libraries used for IFNγ-ELISpot assays as described above. A negative control consisted of unstimulated cells (medium only), whereas concanavalin A (Con A, 10 µg/mL, Sigma-Aldrich, United States) stimulation was used as a positive control for cytokine secretion. Spot number was determined using an ELISpot iSpot Spectrum reader (AID, Strassberg, Germany) and analyzed with the AID ELISpot reader software V7.0 (AID, Germany). Responses were considered positive when the number of spot-forming colonies per million cells was > 50 and at least three-fold higher than the medium-alone negative control.
RESULTS
Cellular immune response prevalence to AAV2, 4, 5, 8, 9, and rh10 in healthy donors
The prevalence of preexisting cellular immune response to AAV serotypes 2, 4, 5, 8, 9, and rh10 was determined using an IFNγ ELISpot assay. PBMCs were isolated from whole blood collected on healthy human donors (n = 45–145 depending on serotypes) from the Pays de la Loire area, France. These cells were restimulated in vitro with an overlapping peptide library spanning the VP1 sequence of each serotype tested (i.e., AAV2, 4, 5, 8, 9, and rh10) and divided into 3 pools. Our results show that AAV9 and AAV8 have the highest prevalence: 46% (n = 66/145) and 24% (n = 35/145) of positive donors, respectively, followed by AAV4 (10%, n = 8/80), AAVrh10 (9%, n = 4/45), and AAV2 (7%, n = 9/121). Only 1 donor out of 84 was found positive for an IFNγ-positive response to AAV5 (Fig. 1A). As expected, due to their strong homology, a coprevalence analysis reveals that 26% of positive donors are also reacting to 2 or more serotypes (Fig. 1B and Table 1). It is noteworthy that all the positive donors to AAV2 are also responding to at least another serotype in contrast to AAV4, AAV8, AAV9, and AAVrh10. All of the AAV2-positive donors are positive for an anti-AAV8 cellular response. The only AAV5-positive donor shows a response to AAV8, AAV9, and AAVrh10 as well. This donor has a positive response toward the AAV pool 2 of AAV5, 8, 9, and rh10 suggesting a shared epitope (Supplementary Table S1). Regarding the AAV8, 31% of the positive donors are also responding to AAV2 and 54% are also positive to AAV9. Interestingly, 73% of AAV9-positive donors are reactive to this serotype only and 27% were found to be positive for at least another serotype. We also analyzed the cellular reactivity to AAV over ages. We did not observe a higher cellular reactivity to a given serotype in a specific age-ranged population for any serotype (Supplementary Table S2).

Cellular immune response prevalence to AAV2, 4, 5, 8, 9, and rh10 in healthy population. PBMCs were restimulated in vitro with an overlapping peptide library spanning the VP1 sequence of each serotypes tested (i.e., AAV2, 4, 5, 8, 9, and rh10) and analyzed using an IFNγ-ELISpot assay.
Coprevalence of Cellular Immune Response to AAV Serotypes 2, 4, 5, 8, 9, and rh10. Anti-AAV-Positive Donors: AAV2: n = 4; AAV4: n = 7; AAV5, n = 1; AAV8: n = 13; AAV9: n = 33; AAVrh10: n = 4
Cellular and humoral immune responses to AAV2, 8, 9, and rh10 are not correlated
We focused on AAV2 and AAV8 since these serotypes have been studied previously and on AAV9 and rh10 serotypes as they are frequently used in preclinical studies and clinical trials presently and for which there are less data available in the literature. For these 4 serotypes, we investigated the humoral response prevalence by analyzing the presence of anti-AAV total immunoglobulins G (IgGs) by ELISA in the serum of healthy donors (Fig. 2A). As already reported, we observed a high seroprevalence to AAV2 (40%, n = 96, IgG titers from 1:40 to ≥1:10 240, Supplementary Fig. S1A) and AAV8 (43%, n = 109, IgG titers from 1:20 to 1:20 480, Supplementary Fig. S1B). 5,11 AAV9 and AAVrh10 show a lower seroprevalence: 29% (n = 88, IgG titers from 1:40 to ≥1:10 240, Supplementary Fig. S1C) and 26% (n = 39, IgG titers from 1:640 to ≥1:10 240, Supplementary Fig. S1D), respectively. We analyzed the coprevalence of AAV-specific IgG in 38 healthy donors (Table 2). Eight donors (21%) were positive for the 4 serotypes. Our results show that 85% of the donors positive for anti-AAV2 IgGs are positive for AAV8 and 77% for AAV9 and AAVrh10 capsids. However, in donors positive for anti-AAV8 and anti-AAV9 IgGs, 71% to 79% are positive to AAV2 capsid. Ninety-three percent of the donors with anti-AAV8 IgGs are responsive to AAV9 and reciprocally. Interestingly, all donors with anti-AAVrh10 antibodies are responding to AAV2 and AAV8 and 90% are responsive to AAV9. The analysis of the humoral reactivity to AAV over ages suggests a higher reactivity to AAV2 and AAVrh10 over time: 23% of the 18- to 35-year-old donors (n = 31) were positive against 60% for the donors older than 55 years (n = 25) for the AAV2, and 9% of the 18- to 35-year-old donors (n = 11) were positive against 50% for the donors older than 55 years (n = 12) for the AAVrh10 serotype (Supplementary Table S3).

Humoral immune response prevalence to AAV2, 8, 9, and rh10 in a healthy population. Sera were collected from healthy human donors and analyzed by ELISA for anti-AAV IgGs.
Coprevalence of Humoral Immune Response to AAV Serotypes 2, 4, 5, 8, 9, and rh10. Anti-AAV IgG Positive Donors: AAV2: n = 13; AAV8: n = 14; AAV9: n = 14; AAVrh10: n = 10
Finally, we analyzed the correlation between the T cell responses and the antibody responses to AAV in our cohort (Fig. 2B). For AAV2, in the donors positive for anti-AAV2 IgGs (n = 39/96), we observed that 85% of the IgG-positive donors (n = 33/39) do not show an IFNγ secretion to the capsid. For AAV8, we observed that 68% of the IgG-positive donors (n = 32/47) do not show an IFNγ secretion to the capsid. For AAV9, we observed that 42% of the IgG-positive donors (n = 11/26) do not show an IFNγ secretion to the capsid. Surprisingly, none of donors that are positive for anti-AAVrh10 IgG-positive donors (n = 0/10) show an IFNγ secretion to the capsid; the donors with a cellular immune response represent 10% of the donors (n = 3/29) who do not have anti-AAVrh10 IgGs. Our results confirmed, as previously reported by others, that there is no correlation between the T cell responses and the antibody responses to AAV in our cohort.
IFNγ-secretion to AAV8 and AAV9 is predominantly mediated by CD8 T cells
Since AAV8 and AAV9 show the highest cellular prevalence (24% and 46%, respectively) and are serotypes presently preferred for high systemic dosing in patients, we wanted to further characterize the cellular immune response. We wondered whether CD8 or CD4 T cells were mediating IFNγ secretion. We performed additional IFNγ ELISpot assays in a representative cohort of positive donors (n = 8 per serotype) using total PBMCs, CD4- or CD8-depleted PBMCs. The efficiency of each CD4 and CD8 depletion was evaluated by flow cytometry and shows less than 1% of remaining targeted cells after depletion (Fig. 3A shows one representative donor). These depletions showed that in all positive donors to AAV8 (n = 8), IFNγ secretion was mediated by CD8 T cells (i.e., by CD4-depleted PBMCs, Fig. 3B for one representative donor). No IFNγ secretion was observed in the CD8-depleted fraction for all donors tested (Supplementary Table S4). Regarding the AAV9 serotype, 7 positive donors were analyzed and also showed that IFNγ secretion was mediated by CD8 T cells (i.e., by CD4-depleted PBMCs, Fig. 3B). Nevertheless, in 3 out of 7 donors, in the CD8-depleted PBMCs (i.e., CD4 T cells), we also observed a slight IFNγ secretion signal above the negative threshold (dotted line) despite an efficient CD8 depletion (Fig. 3C for one representative donor and Supplementary Table S4). Therefore, IFNγ secretion seems to be mainly mediated by CD8 T cells in response to AAV9 with some donors also showing an IFNγ secretion mediated by CD4 T cells: the depletion of CD4+ T cells before ELISpot assay noticeably increased the intensity of responses. This is expected since CD4+ T cells are removed, CD8+ T cells are proportionally more frequent in the tested condition. These results suggest that CD4-depleted cells are the major contributor of IFNγ secretion.
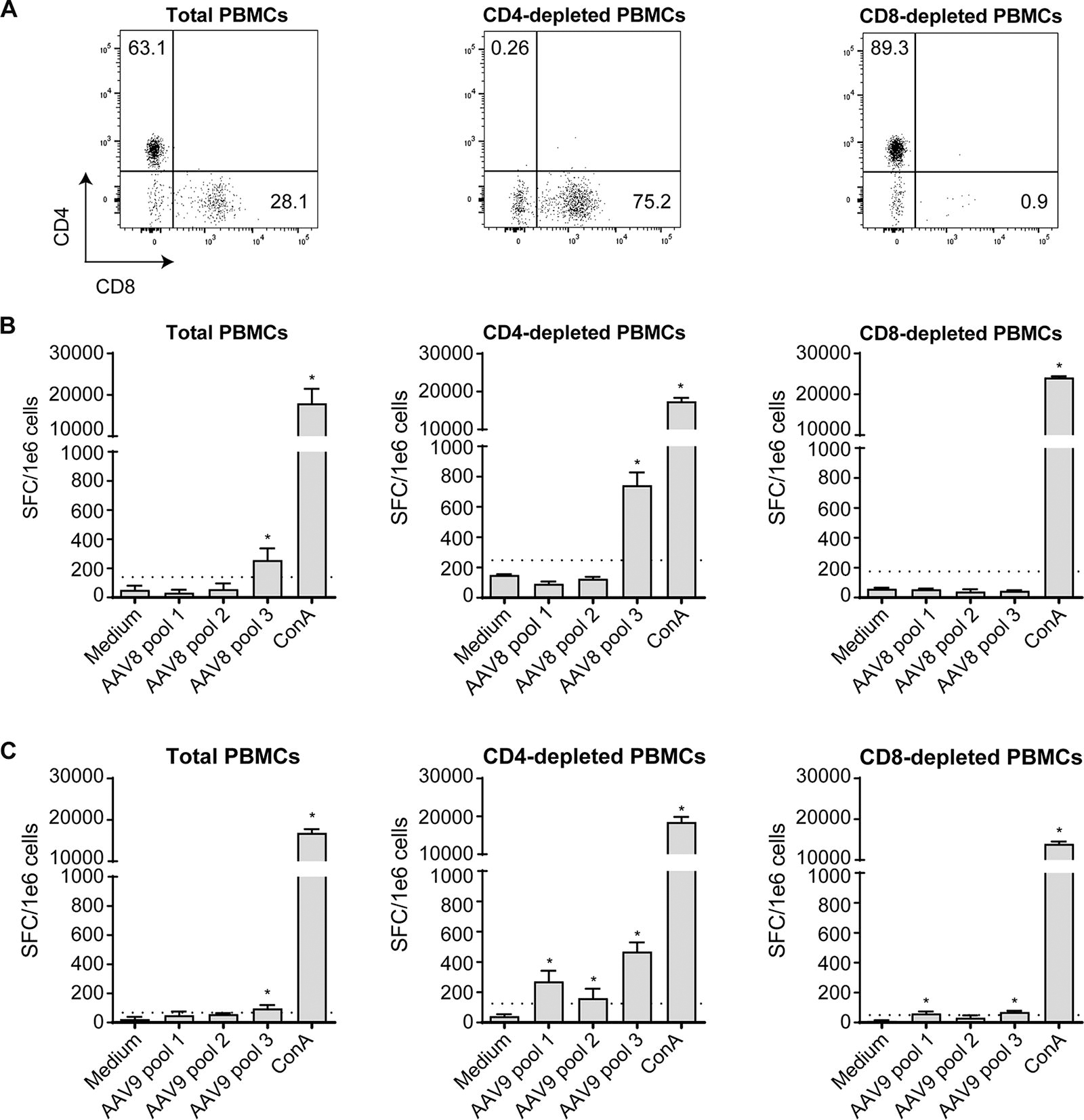
IFNγ secretion to AAV8 and AAV9 is mainly mediated by CD8 T cells.
Cytokine secretion profile of preexisting anti-AAV T cells differs between AAV8 and AAV9
We further characterized the preexisting cellular immune response to AAV8 and AAV9 by performing a polyfunctional FluoroSpot assay. We evaluated the secretion of 3 cytokines indicative of a cytotoxic phenotype simultaneously: IFNγ, IL-2, and TNFα (Fig. 4A). The PBMCs were restimulated in vitro with overlapping peptide libraries spanning the AAV8 or AAV9 VP1 sequence. Six and 8 donors showing an IFNγ-positive response to AAV8 and AAV9, respectively, were tested (Fig. 4B). For each donor, we determined within the IFNγ-secreting cell population whether the cells had 1 (IFNγ+ only), 2 (IFNγ+ only and IFNγ+IL-2+ or IFNγ+ only and IFNγ+TNFα+), or 4 functions (IFNγ+ only, IFNγ+IL-2+, IFNγ+TNFα+, and IFNγ+IL-2+TNFα+). For the AAV8, we observed a diversity in the immune response profile among the 6 donors: 3/6 donors have cells showing 1, 2, and 4 functions, 2/6 donors have cells with 2 functions (IFNγ+ only and IFNγ+TNFα+), and 1/6 donor has cells with 1 function (IFNγ+ only). Surprisingly, for the AAV9, all the donors (n = 8) show the same pattern: 2 functions (IFNγ+ only and IFNγ+TNFα+); no IL-2 secretion was detected in any tested donors. Considering the limited number of healthy donors (n = 6–8), our results may suggest that preexisting cellular immune responses differ between AAV8 and AAV9 and need to be confirmed with additional healthy donor samples.
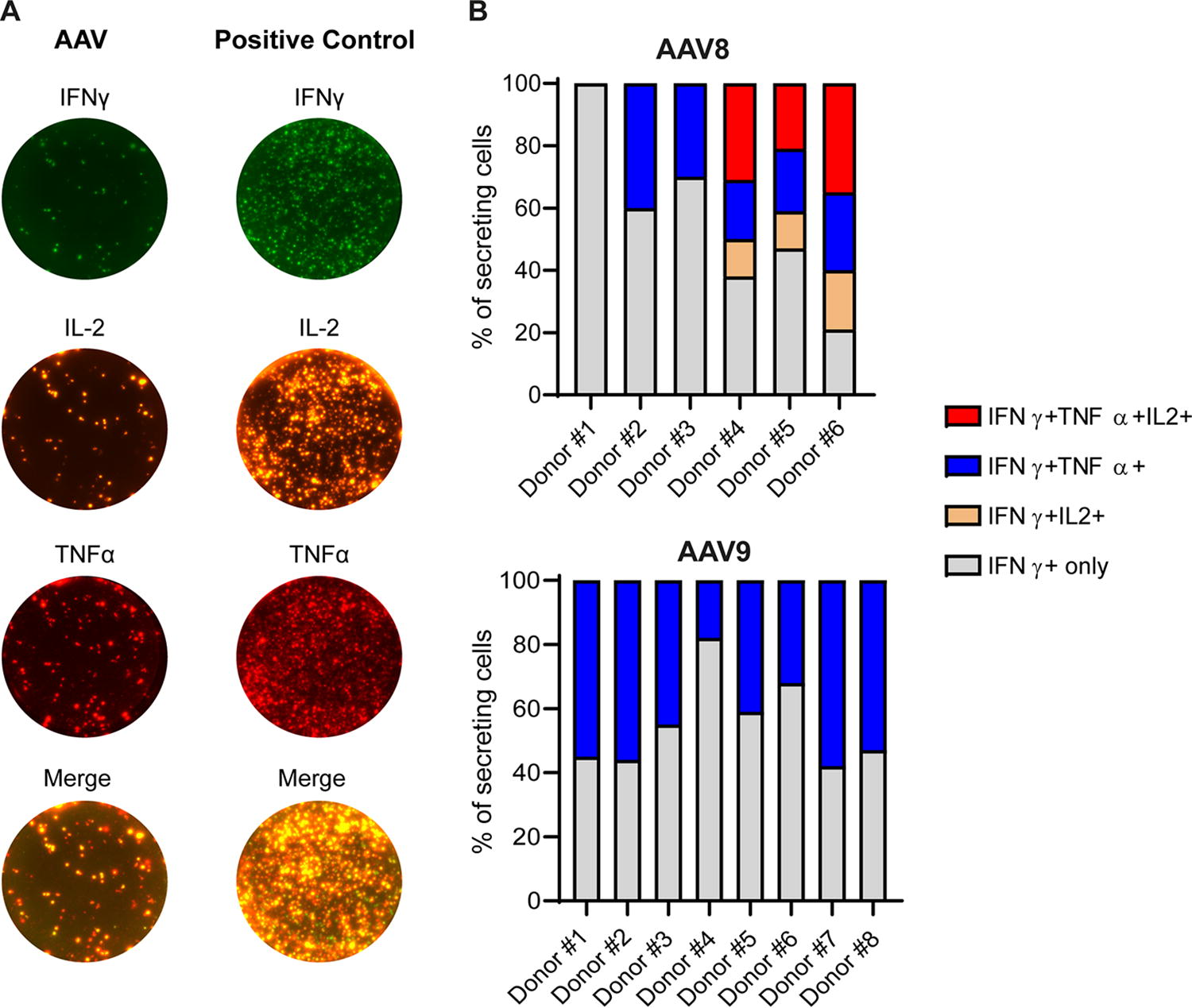
Polyfunctional assessment of anti-AAV8 and anti-AAV9 cellular immune responses. PBMCs were collected from healthy human donors (Donor#1 to #8) and analyzed by IFNγ/IL-2/TNFα FluoroSpot assays. Donors #1 to #6 were previously identified as positive for both anti-AAV8 and anti-AAV9 cellular immune responses. Donors #7 and to #8 were identified as positive for anti-AAV9 cellular immune response.
DISCUSSION
Recombinant AAVs are successful viral vectors for the treatment of inherited disorders as illustrated by recent EMA and FDA approvals of these vectors for the treatment of Leber congenital amaurosis (LCA), 18 SMA, 19 hemophilia A 20 and B, 21 aromatic L-amino acid decarboxylase deficiency (AADC), 22 and Duchenne muscular dystrophy (DMD) 23 and more than 230 clinical trials ongoing. However, when protocols were translated from preclinical models to patients, some limits related to the host immune response emerged and remain a major hurdle since protocols evolved from local to systemic injection of high doses of AAV vectors.
Preexisting immunity to the AAV capsid is one of the major limits for successful clinical translation of rAAV gene therapy products. To the best of our knowledge, only the prevalence of preexisting cellular immunity to AAV1, AAV2, AAV5, and AAV8 has been assessed in the general populations so far. 11 –13,24 Cellular immunity toward more recently emerging serotypes such as AAV9 or AAVrh10 has not been reported yet, whereas they are used in 33 and 10 active or recruiting clinical trials, respectively (clinicaltrials.gov database). In this study, we studied the prevalence of the cellular immune response to 6 AAV serotypes: AAV2, 4, 5, 8, 9, and rh10 in a large cohort of healthy donors. Our results show that AAV9 is the most prevalent (46%) followed by AAV8 (24%), AAV4 (10%), AAVrh10 (9%), AAV2 (7%), and AAV5 (1%). The AAV2 prevalence is comparable with results obtained by Chirmule et al. 24 in a cohort of 45 healthy donors from the United States but lower than reported by Li et al. (47%, n = 17) 13 or by Kruzik et al. (19%, n = 90). 11 In this study, they also described a prevalence of 24% for the AAV5, whereas we showed only 1 positive donor out of 84. These differences across studies might be explained by the different assays used and the absence of standardization of these assays (e.g., discrepancy for the positive thresholds determined in different ELISpot assays), and also by an analysis performed on donors coming from different countries or continents (France versus Germany, Austria, and the United States), which has an impact on natural AAV infections as previously shown for humoral prevalence. 4 These differences highlight the need of standardized assays to compare studies and to measure T cell-mediated immune response to AAV before and after gene therapy in clinic.
The humoral immune response to AAV2, 8, 9, and rh10 has also been analyzed. As already described, the highest IgG prevalence is observed for AAV2 4,5 and AAV8 followed by AAV9 and rh10. This is in accordance with previous studies performed on anti-AAV neutralizing antibodies (NAbs) and/or total IgG antibodies (TAbs). 4,5,11,25 It is noteworthy that some variability may apply between studies since there is no standardized assay with a defined cutoff to measure either NAbs or TAbs in healthy donors or patients enrolled in clinical trials (1:10 in the present study versus 1:20 11 or 1:30, 5 e.g.). Moreover, the seroprevalence may vary depending on the age of the donors and their origin. 3,4,11 As previously described, our results confirm that there is a high cross-reactivity between serotypes; more than 88% of donors responding to AAV2 are responding to AAV8 and 93% of the donors responding to AAV8 are responding to AAV9. The more cross-reactive donors are the ones positive for anti-AAVrh10 (n = 9/39). This can be explained by phylogenic relatedness. 26,27 Our study also confirms that there is no correlation between humoral and the IFNγ-mediated cellular immune responses to AAV vectors. 11,12
As expected, we also report a cellular immune response cross-reactivity between serotypes. This analysis was performed on 73 donors for serotypes 2, 4, 5, 8, and 9 and on 40 donors for serotypes 2, 8, 9, and rh10. Our results show that 26% of the positive donors are also reacting to 2 or more serotypes. All the positive donors to AAV2 were also found responsive to at least another serotype in contrast to AAV4, AAV8, AAV9, and AAVrh10. For AAV2 and AAVrh10, pool 2 elicited a majority of positive response, whereas for AAV4, AAV8, and AAV9 it is the pool 3 (Supplementary Table S1). Surprisingly, 73% of AAV9-positive donors are reactive to this serotype only, whereas the AAV9, AAV8, and AAVrh10 VP1 amino acid sequences share more than 80% of homology (Supplementary Table S5). For AAV2, 4 out of 6 of the positive donors were responding to pool 2 or 3 with 2 donors responding to both peptide pools. These 2 peptide pools contain immunodominant peptides that were previously described in HLA-A*0201 and HLA-B*0702 restricted donors. 28 Some of these immunodominant peptides are also present in AAV9 and AAVrh10 pool 3 although might not be the immunodominant peptides for these particular capsids. Indeed, for these 2 serotypes, donors sharing positive IFNγ-ELISpot do not necessarily respond to the same peptide pools: a positive donor for both AAV2 and AAV9 might have a positive response to AAV2 pool 2 and AAV9 pool 3 (not shown). This observation has been made for all the studied serotypes except AAV5 where the only positive donor is also positive for AAV8, AAV9, and AAVrh10 pool 2. An in-depth analysis using a matrix approach would be useful to identify immunodominant peptides depending on donor’s HLA genotype and to develop engineered capsids to lower or prevent the adaptive immune response without altering AAV tropism such as described for a neutralizing antibody response. 26,29
Then, we performed an in-depth analysis of the positive response to AAV8 and AAV9 (which are at present preferred for systemic administration and used in patients (NCT02122952; NCT03199469; NCT03368742). They also appear to be the most prevalent serotypes for a cellular immune response in our healthy donor cohort. Our results show that the preexisting cellular immune response to AAV8 and AAV9 is mainly mediated by CD8 T cells. Regarding AAV9, some of the donors are also showing an IFNγ secretion mediated by CD4 T cells. This observation has also been described for AAV2 13 and AAV1 12 and reactive cells were identified as CD4+ effector memory T cells. Since CD8 and CD4 depletions were performed on total PBMCs to evidence a CD8- or CD4-mediated response, we cannot exclude that IFNγ secretion might also be mediated by other immune cells especially in seronegative donors. Indeed, Kuranda et al. described that CD16brightCD56dim natural killer (NK) cells are the major contributor of IFNγ secretion in AAV-seronegative healthy donors toward the AAV2 capsid. 30
Finally, another major concern in AAV-mediated gene transfer is the fact that IFNγ secretion alone, as measured today, is not sufficient to predict the immune outcome of gene transfer. Actually, the IFNγ secretion is not always associated with a loss of transgene expression, 31,32 suggesting that, aside the vector serotype, the dose, or the transgene, the anti-AAV immune response after gene transfer might vary depending on the immune patient profile. While we performed the prevalence study using the standard IFNγ ELISpot assay, we also performed an in-depth characterization using a multiparametric IFNγ/IL-2/TNFα FluoroSpot assay on a limited cohort of healthy donors (n = 6–8). Our results show a diversity in immune response profile among the 6 AAV8-positive donors, whereas the 8 donors tested for AAV9 showed the same pattern with cells that are not secreting IL-2. The absence of IL-2 secretion suggests that these cells may not be cytotoxic effector T cells but rather be exhausted or terminally differentiated effector memory (TEMRA). 33,34 Even though these T cell subtypes are reported in chronic viral infections, 35,36 they have also been described in the context of AAV. CD8-exhausted T cells expressing the PD1 marker have been described after AAV-mediated gene transfer in mice, nonhuman primates, and humans, 37 –39 whereas TEMRA cell reactivity to AAV has been described in healthy donors. 40 Further investigation, including phenotyping assays, will be necessary (i) to confirm that T cells are actually exhausted or terminally differentiated in the present study and (ii) to better characterize and understand preexisting anti-AAV T cells.
CONCLUSION
Altogether, our study shows a high cellular prevalence to AAV9 and AAV8 in healthy donors based on IFNγ secretion. Since this cytokine cannot be considered a specific and exclusive marker for deleterious immune responses as shown in clinical trials, it does not appear relevant to prescreen eligible patients to AAV-derived gene therapies yet in opposition to anti-AAV NAbs or TAbs that can prevent cell transduction 41 or lead to innate immune pathway activation. 42 Although the present study highlights the need of better characterizing the preexisting T cell response to AAV using multiparametric assays and developing relevant in vitro and in vivo models to test them and foresee the outcome on gene transfer efficacy.
Footnotes
ACKNOWLEDGMENTS
The authors acknowledge the Cytocell-Flow Cytometry and FACS core facility (SFR Bonamy, BioCore, Inserm UMS 016, CNRS UAR 3556, Nantes, France) for its technical expertise and help, member of the Scientific Interest Group (GIS) Biogenouest and the Labex IGO program supported by the French National Research Agency (n°ANR-11-LABX-0016-01). They also acknowledge the UMR1089 Vector Core (Vivem) for providing the AAV vectors for ELISA screening, the EFS for providing them with buffy coats, and Manon Loirat and Marion Grard for their help on performing ELISA and ELISpot assays.
AUTHOR CONTRIBUTIONS
Conceptualization: R.X., C.V., O.A., and G.G. Investigation: R.X., A.A., C.V., N.J., M.D., C.C., J.L.D., A.C., M.G., and G.G. Methodology: R.X., A.A., C.V., O.A., and G.G. Visualization: R.X., A.A., O.A., and G.G. Writing—original draft: R.X. and G.G. Writing—review and editing: R.X., A.A., M.G., O.A., and G.G. Supervision: O.A. and G.G. Funding acquisition: O.A. and G.G.
AUTHOR DISCLOSURE
A.A., M.D., N.J., C.C., J.L.D., A.C., M.G., O.A., and G.G. declare no competing financial interests with regard to this work performed at INSERM TaRGeT UM1089 (Nantes, France). R.X. is at present an employee of Roche, Basel, Switzerland. C.V. is at present an employee of Charles River Laboratories, Evreux, France.
FUNDING INFORMATION
This work was supported by the INSERM, the French Ministry of Research, the FRM (Fondation pour la Recherche Médicale), the University Hospital of Nantes, the Fondation pour la Thérapie Génique en Pays de Loire, Région Pays de Loire (IMBIO-DC consortium), EU Horizon 2020 (UPGRADE-Unlocking Precision Gene Therapy)-grant agreement N_825825, the French National Research Agency (ANR) (ANR-22-CE18-0043-01), and the IHU-CESTI, which is supported by the ANR via the “Investment Into The Future” program ANR-10-IBHU-005.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.