
Abstract
The advent of clustered regularly interspaced short palindromic repeats (CRISPR)–CRISPR-associated nuclease 9 (Cas9) technology has revolutionized the field of genetic engineering, offering unprecedented potential for the targeted manipulation of DNA sequences. Advances in the mechanism of action of the CRISPR-Cas9 system allowed potential applicability for the treatment of genetic diseases. CRISPR-Cas9’s mechanism of action involves the use of an RNA guide molecule to target-specific DNA sequences and the Cas9 enzyme to induce precise DNA cleavage. In the context of the CRISPR-Cas9 system, this review covers nonviral delivery methods for gene editing based on peptide internalization. Here, we describe critical areas of discussion such as immunogenicity, emphasizing the importance of safety, efficiency, and cost-effectiveness, particularly in the context of treating single-mutation genetic diseases using advanced editing techniques genetics as prime editor and base editor. The text discusses the versatility of cell-penetrating peptides (CPPs) in forming complexes for delivering biomolecules, particularly ribonucleoprotein for genome editing with CRISPR-Cas9 in human cells. In addition, it emphasizes the promise of combining CPPs with DNA base editing and prime editing systems. These systems, known for their simplicity and precision, hold great potential for correcting point mutations in human genetic diseases. In summary, the text provides a clear overview of the advantages of using CPPs for genome editing with CRISPR-Cas9, particularly in conjunction with advanced editing systems, highlighting their potential impact on clinical applications in the treatment of single-mutation genetic diseases.
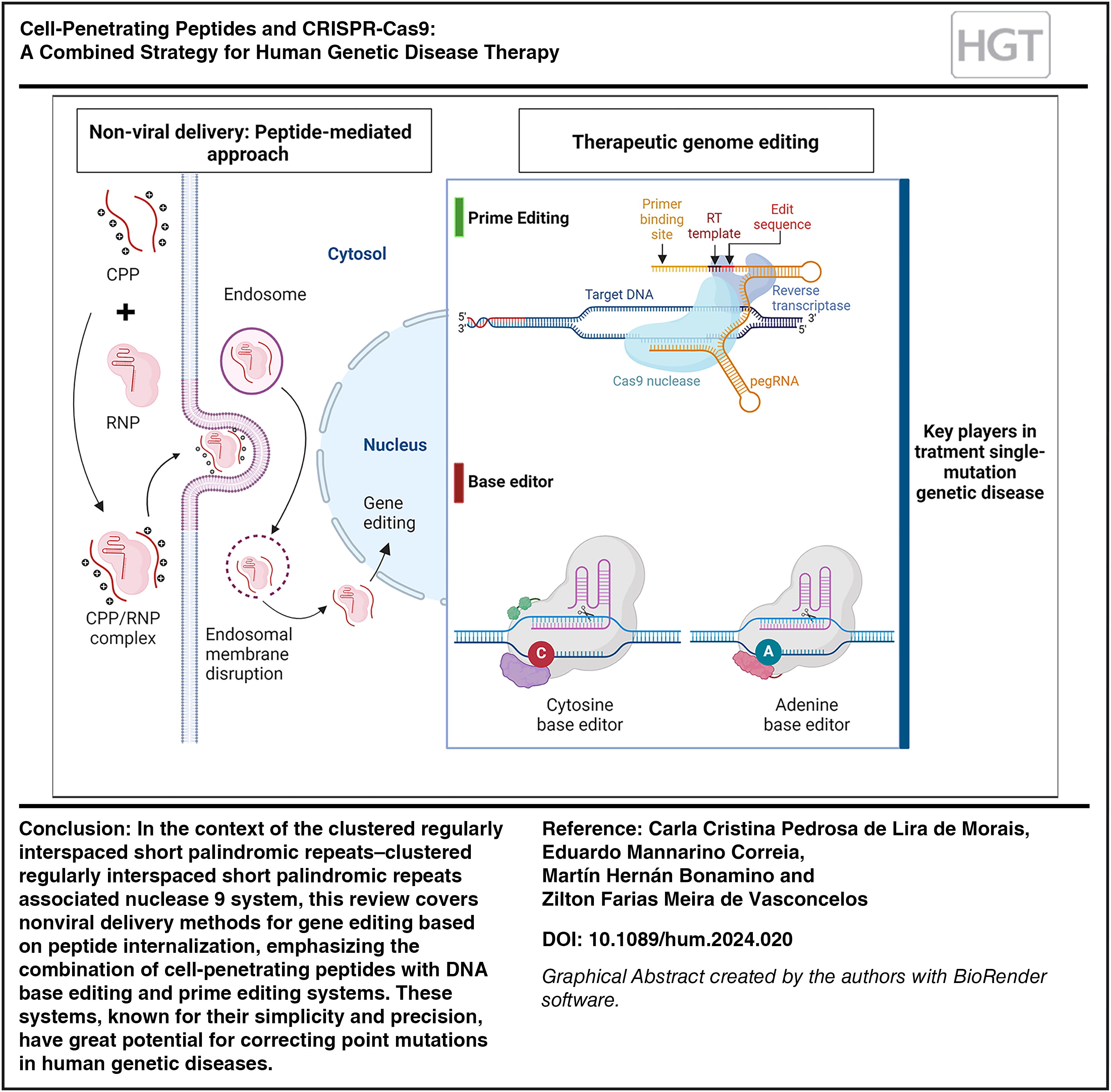
INTRODUCTION
In a concise view of the three historical periods in the development of molecular biology, focusing on the impact of DNA markers and genome sequencing, we highlight the era of pre-polymorphic DNA markers (1956–1979) characterized as the fundamental stage of the development of genetic theory within molecular biology. During this period, scientists were laying the groundwork for understanding genetics at the molecular level. The mention of “polymorphic DNA markers” signifies the critical shift that followed; the Pre-Genome era (1980–2001) coincided with the discovery of polymorphic DNA markers, a crucial moment in genetics research, as these markers allowed the creation of panels within the genome, which, in turn, facilitated the identification of responsible genes by Mendelian diseases, those caused by single genetic mutations; the Post-Genome era (2002–2010) followed the publication of human genome sequencing results in 2001, which marked a dramatic shift in genetic research due to the availability of comprehensive genetic information and the development of extensive marker panels, which made it possible to identify genotypic alterations without the need for prior knowledge of the individuals’ phenotypes. This was crucial for studying complex diseases influenced by multiple genetic and environmental factors over an individual’s lifetime. 1
The latest update to Online Mendelian Inheritance in Man® (OMIM®) shows that, of the ∼25,000 annotated genes in the human genome, mutations in over 4,800 genes have already been linked to disease phenotypes. 2
Advances in the field of genetics have enabled significant progress in the interpretation of disease-causing genetic variants and underscores ongoing efforts to decipher the complex genetic underpinnings of various diseases. This is a crucial aspect of personalized medicine as it allows for more accurate diagnoses and treatment strategies tailored to an individual’s genetic makeup. The ability to more accurately identify and classify these variants can revolutionize the approach to genetic diseases. Furthermore, such advances have allowed the intersection of genetics and medicine making mention of the ability to manipulate genes punctually using short nucleotide insertion or deletion is particularly noteworthy. This precision gene editing techniques, often associated with CRISPR-Cas9 technology, have opened new possibilities for therapeutic interventions (Fig. 1). By directly targeting the genetic root of disease, researchers and clinicians can unlock potential cures and treatments that were previously considered out of reach.

Different CRISPR editing tools discussed in the review. Created in BioRender.com. CRISPR, clustered regularly interspaced short palindromic repeats.
The exploration of peptide carriers for delivering therapeutic molecules to target cells has been a focus of research in recent decades. Currently, cell-penetrating peptides (CPPs) are being investigated for treating a range of diseases and serving as vehicles for targeted delivery. CPPs have the capability to transport therapeutic molecules not only to target cells but also to cellular organelles, demonstrating their versatility in drug delivery. 3 In the context of the CRISPR-Cas9 system, this review covers nonviral delivery methods for gene editing based on peptide internalization. Here we describe critical areas of discussion such as immunogenicity and post-editing cell recovery rate, emphasizing the importance of safety, efficiency, and cost-effectiveness, particularly in the context of treating single-mutation genetic diseases using advanced editing techniques genetics as prime editor and base editor.
CRISPR-CAS SYSTEM: A BRIEF HISTORY
The CRISPR-Cas system was first identified in 1987 when Japanese scientists were investigating the IAP gene in Escherichia coli (E. coli) bacteria. During their research, they discovered an unusual genetic structure adjacent to this gene, characterized by alternating repeated DNA sequences and non-repeating sequences. At that time, the biological significance of this structure was not understood. 4 Only 20 years after this observation, the biological function of this system was revealed. Then, experimental evidence established CRISPR as a crucial element of the bacterial defense system against bacteriophage infections. 5
In 2002, Jansen and collaborators designated the repeated sequences as “Clustered Regularly Interspaced Short Palindromic Repeats,” a universally accepted CRISPR acronym, located in the genome close to the “DNA repair system.” 6 Proximal genes, previously thought to be part of the DNA repair system, have been found to accompany the CRISPR gene and are therefore called CRISPR-associated genes (Cas). 7
In 2006, Koonin and his collaborators computationally analyzed the link between the functions of the CRISPR and CAS genes as a system and observed a similarity to a prokaryotic RNA-interfering immune system. 8
In 2007, Barrangou et al. 5 experimentally demonstrated the role of the CRISPR-Cas system in the dairy bacterial species Streptococcus thermophilus (S. thermophilus). After exposure to virulent (also known as lytic) streptococcus phages, the vast majority of phage-infected cells died, and in the small number of surviving cells, it was observed that a new spacer was integrated into the CRISPR locus along with the addition of a repeat.
In 2008, Marraffini and Sontheimer 9 demonstrated interference activity against plasmids. It has been shown in E. coli that the CRISPR locus is transcribed and further processed by Cas nucleases to serve as small guide RNAs (gRNA) that allow a complex of Cas proteins and CRISPR-RNA (crRNA) to interfere with the occurrence of an invasion by recognized foreign DNA.
In 2010, Garneau et al. 10 demonstrated that the S. thermophilus CRISPR-Cas system was capable of accurately cleaving target phage and plasmid DNA within the protospacer sequence in vivo, providing a clear explanation for phage resistance and plasmid interference. Unlike what happens in E. coli, where the need for several Cas nucleases in the interference step has been observed, in S. thermophilus and Streptococcus pyogenes, Cas9 is the only nuclease needed to cut the target DNA. 10,11
Bacterial immunity via CRISPR-Cas involves three phases: adaptation, expression, and interference. In the adaptation phase, also called insertion or acquisition, DNA fragments (protospacers) from invading viruses and plasmids are incorporated into the CRISPR matrix through the action of a distinct complex of Cas proteins that bind to the target DNA, often after recognizing a distinct, short motif known as the protospacer-adjacent motif (PAM), and cleaves a portion of the target DNA, the protospacer. After duplication of the repeat at the 5′ end of the CRISPR array, the adaptation complex inserts the protospacer DNA into the array to become a spacer. Some CRISPR-Cas systems employ an alternative adaptation mechanism, that is, RNA spacer acquisition, via reverse transcription by a reverse transcriptase encoded at the CRISPR-Cas locus. 5,12
In the expression phase, the CRISPR locus is transcribed into pre-CRISPR RNA (pre-crRNA), which is processed into mature CRISPR RNAs (crRNAs), each containing the spacer sequence and parts of the flanking repeats. 13
In the interference stage, the crRNA, which normally remains bound to the processing complex, serves as a guide to recognize the protospacer (or a very similar sequence) in the invading genome of a virus or plasmid, which is then cleaved and inactivated by one or more Cas nuclease(s) that are part of the effector complex or are recruited at the interference stage. 14
The diversity and number of CRISPR-Cas systems has increased substantially in recent years, and according to the most recent classification, published in 2020, it includes 2 classes, 6 types, and 33 subtypes. 12 Class 1 systems have effector modules composed of multiple Cas proteins, some of which form crRNA-binding complexes that contribute to pre-crRNA processing and interference. In contrast, class 2 systems encompass a single multidomain crRNA-binding protein (such as Cas9 in type II systems) that combines all the activities required for interference. 12
The CRISPR system, which uses the Cas9 nuclease guided by an RNA molecule, composed of 100 unique nucleotides, is the most studied currently. The structure of S. pyogenes Cas9 (SpCas9) reveals a bilobed architecture composed of a recognition lobe (REC) and a nuclease lobe (NUC). 15 The REC lobe has two domains: (1) REC, responsible for binding single-stranded guide RNA and (2) bridge helix (BH), which plays a role in initiating cleavage activity shortly after DNA binding. The NUC lobe is composed of three domains: (1) integration with PAM (PI), responsible for binding PAM, contributing to the separation of the double strand of target DNA and the formation of the gRNA-DNA hybrid; (2) HNH, responsible for cutting the complementary strand to the gRNA target DNA; and (3) RuvC, responsible for cutting the noncomplementary strand to the gRNA target DNA. Thus, the HNH and RuvC domains work together when activated with the recognition of the PAM sequence, resulting in the DNA double-strand break. 16,17
SpCas9 recognizes a 5′-NGG-3′ PAM located downstream of the cleavage site on the complementary DNA strand and generates “blunt ends” after 3 base pairs above the PAM site. 18 The purine, guanine, and adenine bases (G and A, respectively) at position 20 of the gRNA proximal to the PAM appear to have a positive impact on SpCas9 performance, increasing the effectiveness of genome editing. 19 However, concern about genome editing resulting from double-strand break repair through Cas9 continues to be considered due to the existence of a potential for off-target DNA cleavage, which can cause undesirable mutations, as Cas9 is capable of recognizing and cleave regions of DNA that may not perfectly correspond to gRNA or tracrRNA-crRNA. 20
DNA double-strand breaks are among the most genotoxic DNA lesions that, if not repaired, can cause chromosomal rearrangements, genomic instability, and cell death. Eukaryotic cells have evolved several pathways to repair DNA double-strand breaks. 21 The two main pathways are homologous recombination (HDR), error-free and dependent on a DNA template, and classical non-homologous end joining (NHEJ), independent of a template and error prone. 21 –23 Additional pathways include microhomology-mediated end joining (MMEJ), also known as alternative end joining (alt-EJ), and single-strand annealing (SSA), both error-prone. 24,25
The target locus, upon being cleaved by Cas9, undergoes one of the two main DNA repair pathways. In the absence of a template DNA, the DNA strands are relegated through the NHEJ pathway, with the initial recognition of the Ku70/80 heterodimer that protects the ends of the DNA from processing by nucleases and acts to recruit the other proteins necessary for repair; however, this type of repair can cause insertions, deletions, or precise correction of the target DNA sequence, and this mechanism can be used to mediate the inactivation of specific genes. 16,26 In the presence of template DNA, the HDR pathway, which occurs at a lower frequency than NHEJ, can be leveraged to modify the target locus. 16 When NHEJ is absent due to lack of Ku or the DNA ligase complex, blunt ends can be joined using the MMEJ repair mechanism that involves the alignment of microhomology sequences (within 5–25 bp) flanking a double-strand break. This process results in either deletions between homology arms or, in some cases, complex insertions. 17
The molecular mechanisms of preferential post-cleavage cellular repair using CRISPR-Cas9 are not yet fully understood. 27 However, the choice of which type of repair pathway will be activated depends on many factors, such as cell cycle phase, chromatin structure, and CRISPR-Cas construction. 28 –30
Although the detailed mechanisms are not yet elucidated, the local structure of chromatin can affect both the efficiency of Cas9 cleavage and the choice of DNA double-strand break repair pathways. 21,31 The efficiency of Cas cleavage is dependent on the accessibility of the DNA target site, and in eukaryotic cells, nucleosomes, formed by histones, may represent a barrier to Cas9. 32
The CRISPR-Cas9 system, due to its simplicity and cost, compared with other gene editing platforms, has already been widely used as a research tool, and its application is currently directed to biotechnology and medicine, for the treatment of genetic disorders, which concerns human health. 33 Therefore, the application of CRISPR-Cas9 can be divided into five groups: (1) innate Cas9-mediated genomic modification; (2) mutated Cas9-mediated genomic modification; (3) Cas9-mediated activation or inactivation of transcriptional control; (4) high-throughput genome modification and silencing; and (5) separation of certain genomic areas using Cas9 with inactive catalytic sites attached to a marker (Fig. 2). 34 –38
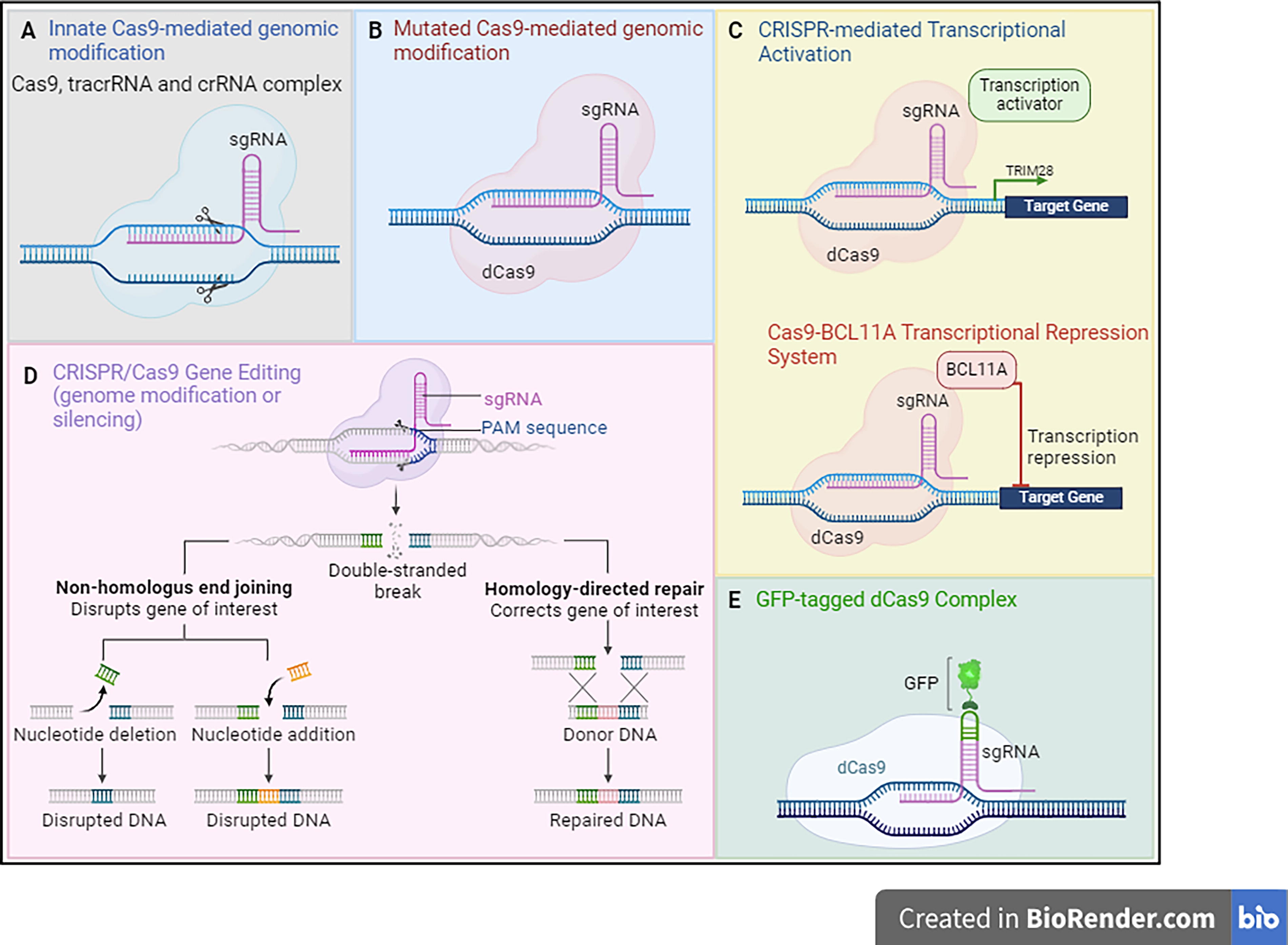
Applications of CRISPR-Cas9 system in genetic disease. (
CRISPR-CAS TECHNOLOGY IN CLINICAL GENE THERAPY APPLICATIONS
Studies in cells or animals have shown that the CRISPR-Cas system represents a significant advance in gene therapy because they have demonstrated the ability to be administered in vivo by methods that are potentially translatable to human use. 39 –41 However, considering clinical use as a final application, researchers have focused on overcoming still existing obstacles, such as off-target effects, efficacy, and development of in vivo corrected gene delivery methods, HDR efficacy, immunogenic potential of the delivery vehicle, and capacity of edited cells. 42
Different protocols have been developed for the detection of off-target effects of the Cas9 nuclease on the genome of cells. 43 –48 Although the standardization of optimization methods is still necessary, these protocols, for identifying potential off-target sites, associated with computational tools used to design guide RNA, are capable of predicting the effectiveness of the CRISPR-Cas system. 48 To increase the effectiveness of editing via HDR, Yu and collaborators, in 2015, showed that small molecules, such as Brefeldin A and L755507, can modulate the activity of the CRISPR-Cas system, effectively increasing repair activity via HDR in the presence of a donor sequence. 49
The delivery method is an important factor that can also impact off-target undesirable effects. 47,50 There are three main delivery methods for Cas9, each with advantages and disadvantages: (1) the mature Cas9/gRNA ribonucleoprotein (RNP) complex can be delivered by different routes, with the advantage of tight control over timing and dosage of the Cas9/RNAg complex, which can minimize off-target effects with editing faster due to the elimination of the need for intracellular transcription and translation, in addition to offering a robust platform for cells with low transcription and translation activity, also allowing its applicability in various cell types and, as a disadvantage, presents the challenge of delivering a large complex, in addition to the potential to trigger immunological responses 50 –53 ; (2) the genes encoding Cas9 and the gRNA can be delivered with a DNA vector, such as a plasmid or viral vector, the advantage in this case is the long-term expression of Cas9 and gRNA; however, this persistent expression of the complex can increase off-target effects 54 –58 ; and (3) the gRNA can be delivered together with the messenger RNA encoding the Cas9 protein, the advantage of this approach is the transient expression of Cas9 within a limited time window, minimizing off-target effects; however, this method has limited efficiency for genome editing due to rapid degradation of the messenger RNA encoding Cas9. 21,59
The use of CRISPR-Cas9 in the manipulation of laboratory animal genomes has contributed to the understanding of many gene functions; in addition, it has led to multiple advances in the diagnosis of human diseases and the implementation of targeted therapeutics. Although there are still some challenges that need to be overcome for the safe and effective use of CRISPR-Cas technology in clinical gene therapy applications, advances in the area have allowed clinical trials using CRISPR-Cas9 approach for different disorders (source: ClinicalTrial.gov accessed on 02/06/2024), as demonstrated in Table 1, one of them recently approved by the Medicine and Healthcare Products Regulatory Agency in the United Kingdom of the first CRISPR-Cas9-based gene therapy in the world, called Casgevy, for the treatment of sickle cell anemia and beta-thalassemia (Fig. 3). However, the benefits of RNP delivery are emerging in the broader context of genome editing, as is its potential for clinical use.

Timeline focused on advancing CRISPR technology aimed at treating genetic diseases and the development of delivery strategies. Cas9, clustered regularly interspaced short palindromic repeats-associated nuclease 9.
Overview of clinical trials using CRISPR-Cas9 and/or CPP approach for different disorders
CPP, cell-penetrating peptides; HIV, human immunodeficiency virus; HPV, human papillomavirus; NA, not applicable.
NONVIRAL DELIVERY: PEPTIDE-MEDIATED APPROACH
One of the main delivery strategies used is electroporation, which brings several problems. Some of them are related to the recovery of the cells after the procedure and the alteration of the transcriptome, which can be involved in lower cell yield, possibly impairing some applications of the RNP use in the manufacturing of modified cells for gene therapy.
Kim et al. 58 delivered RNPs instead of plasmids, via the electroporation route, to modify genes in human primary and embryonic stem cells (ESCs) and revealed that high gene editing efficiency could be achieved in these cell types that were difficult to transfect. However, physical methods are widely used in vitro but have limits regarding in vivo applications, mainly because of several physiological obstacles. Therefore, as an alternative, chemical delivery methods, which have the merits of favorable biocompatibility, low immunogenicity, and flexible cargo size, have received more attention. 60
As demonstrated by Foss and colleagues, comparisons of electroporation and peptide-mediated delivery in terms of cell yield, efficiency, and transcriptome disturbance show better outcomes with the use of peptides, although some minimal loss of editing was noticed. 61
The last method listed above tries to optimize the use of RNP in cost and aggressiveness to the cells. Those peptides are called CPPs and are classified based on their origin, chemical charges, biding type, sequence, and physical–chemical properties, each one of them with subclassification, detailed review in Fig. 4. 3,62 In the realm of nonviral delivery of CRISPR-Cas components, CPPs have emerged as a promising strategy for effective delivery. CPPs are short peptides typically composed of 5–30 amino acids, often carrying a positive charge. They can be categorized based on their hydrophobic and amphipathic properties. While certain groups of CPPs share high sequence identity and structural features, as a whole, CPPs lack sequence homology, resulting in diverse absorption patterns influenced by cell lineage and environmental conditions. 63,64
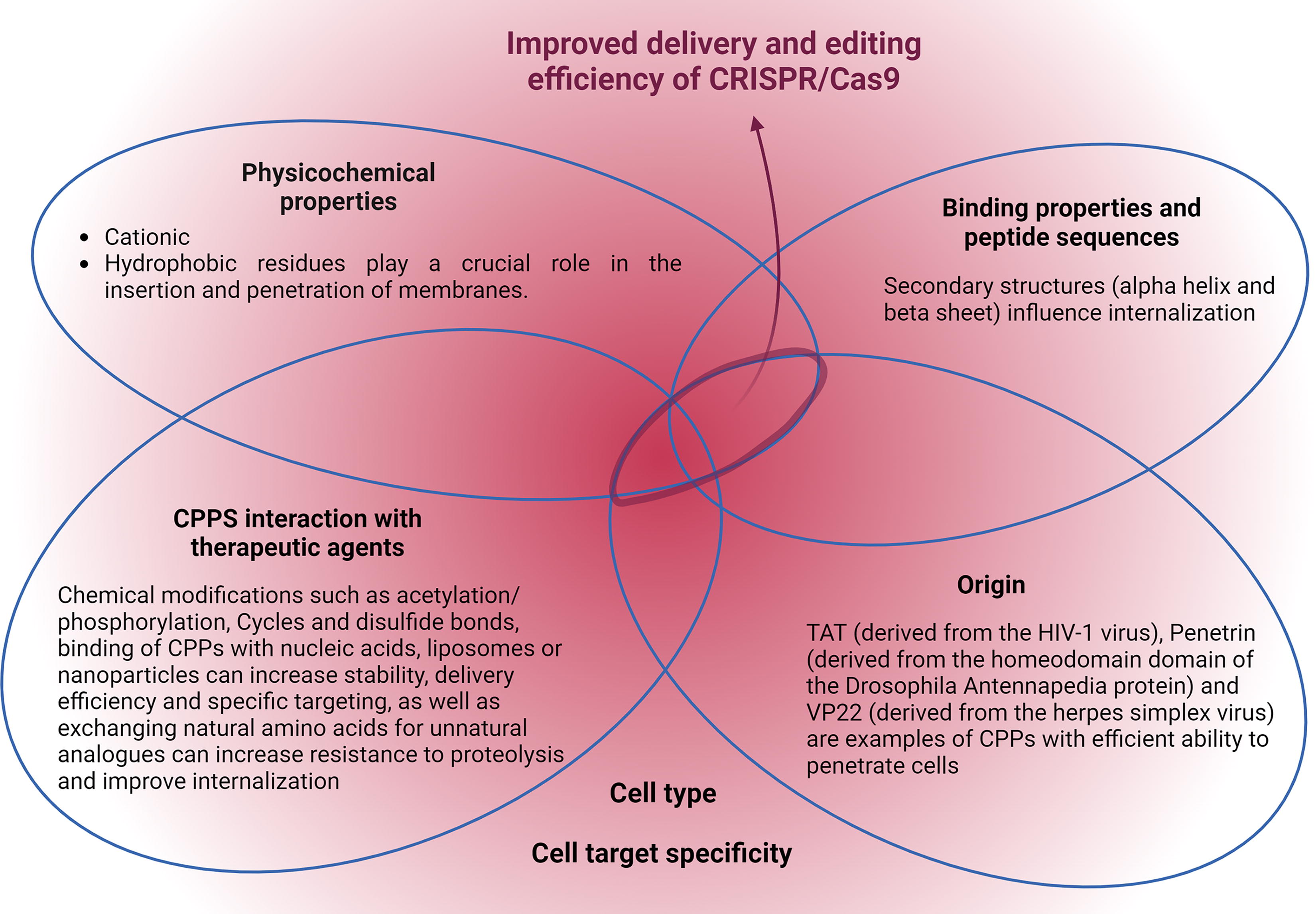
Features of CPPs that improve delivery and editing efficiency. The strategies, when combined, can lead to significant improvements in gene delivery and editing, increasing editing efficiency and cell target while minimizing adverse effects. However, cell type and target specificity must be considered. Created in BioRender.com. CPP, cell-penetrating peptides.
Moreover, the type of cargo attached to CPPs, whether covalently or noncovalently linked, also influences absorption dynamics. The realm of CPPs encompasses over 100 types, with the list continually expanding. Recent classifications have categorized CPPs based on physicochemical attributes, structural characteristics, origin, application, and entry mechanisms. 64 –67 A functional taxonomy classification proposed by Gori and collaborators offers a recent framework for understanding CPP diversity 3 (Table 2).
CPPs classification and general characteristics. Adapted, in table format, from Gori et al, 2023 (3)
Although these peptides were first introduced in 1988 with the uptake of trans-activating transcriptional peptide (TAT), derived from human immunodeficiency virus type 1 (HIV-1), 68 its application for delivery just came out in 2011 with the carriage of small interfering RN, using in the formulation the TAT peptide. 69 The CPPs have two major uptake strategies that will depend on the type of peptide, direct translocation through the membrane (with no cost of energy) and endocytosis (energy-dependent), the TAT peptide, commonly used, employs the last one. With this, it faces the problem of endosomal escape to deliver the cargo, with the advantage of not having limits for the type of cargo.
Endosomolytic peptides have a lower penetrance into the cell, requiring further modifications to alone penetrated, but it has a great endosomal escape due to the capability of the acidification of the endosome to change its conformation in a pH-dependent matter allowing the cargo release. One of the most used is the glycopolypeptide HA2 derived from the influenza virus, with the disadvantage of its lower solubility and high hydrophobicity. 70 –72 With this in mind, one strategy to improve the peptide-mediated delivery is using chimeric ones, in this case, derivatives from a fusion between TAT and HA2, increasing the cell-penetrating and endosomal escape capability. This allows only a co-incubation of those peptides containing the cargo with the target cell, not requiring difficult and expansive machinery to deliver the RNP complex, as demonstrated by Foss and colleagues. 61
The therapeutic potential of RNP delivered via CPPs has garnered attention, particularly for in vitro gene editing to address genetic diseases. In 2021, Gustafsson and collaborators 73 investigated the repurposing of the RNA-delivering CPP, PepFect14 (PF14), for noncovalent RNP delivery. Their study demonstrated high rates of gene editing (up to 80% in HEK293T cells) with negligible toxicity. Importantly, CPP-RNP complexes exhibited robust stability under various storage conditions, including freeze–thaw cycles and freeze-drying, without compromising editing efficiency. This underscores the complementarity of CPP-based delivery alongside existing technologies, presenting novel avenues for RNP delivery with potential applicability to diverse gene editing modalities.
In 2022, Shalay and collaborators 74 combined a small fragment of the rabies virus glycoprotein (C2) with CPP PF14 (C2-PF14) to form complexes that deliver Cas9 RNP to neuronal cells. They observed that the RNP/C2-PF14 complexes form nanosized, monodisperse, and nontoxic nanoparticles capable of specifically targeting neuronal cells. This proof of concept illustrates a straightforward and safe approach to targeted genome editing for treating Parkinson’s disease and other neurological disorders.
Modifications to peptide groups have led to increased success rates in gene editing, making them compatible with the delivery of DNA templates for homology repair. In 2023, Öktem and collaborators 75 demonstrated the use of an amphipathic CPP called LAH5 for gene editing of the CCR5 receptor, crucial in the HIV viral adsorption process. They combined the peptide with RNP and a single strand of DNA for genomic DNA repair, achieving up to 70% targeted indel editing across various cell lineages. Moreover, the DNA homology repair rate reached 20% in HEK293T cells. In both cases, whether knockout or knock-in, this delivery strategy showed minimal cytotoxicity in tested cells. Although editing efficiencies varied from 20% to 70%, sufficient levels of editing were consistently achieved. However, future research should explore the stability of these nanocomplexes under simulated in vivo conditions, such as exposure to whole serum, to gain further insights into their stability.
Regarding in vivo application of CPPs, one of the major concerns when talking about delivery of therapeutic agents for brain therapy is overcoming the blood–brain barrier. One way to circumvent this problem was performed by dos Santos and collaborators. They used a lipid nanoparticle decorated with CPP, to enhance internalization, and transferrin ligand, to direct the liposomal vesicle to the transferrin receptor, loaded with a plasmid DNA. With this, they achieved around 6% of crossing the blood–brain barrier, without tissue damage. 76 Their result shows a possible strategy to be explored for drug or gene delivery that needs to cross the blood–brain barrier.
More recently, in 2023, Takenaka and collaborators 77 explored the use of CPPs to deliver proteins into insects, specifically aphids such as Acyrthosiphon pisum. Traditional delivery strategies for protein complexes, such as CRISPR RNP, in insects rely on microinjections, which are unsuitable for this type of animal. Takenaka’s team used Penetratin, a CPP conserved in insects, and fused it with a fluorescent protein marker called mVenus to track the delivery. This method was successfully taken up in vivo. Additionally, a further review of CPP use in vivo was done by Bolhassani collaborators in 2017, and more recently, in 2023, by Gori and collaborators. 3,62 Taken together, these data show the applicability of CPPs in vivo.
STRATEGIES IN THE DELIVERY OF CAS9 RIBONUCLEOPROTEIN
For effective gene editing, delivery of CRISPR-Cas9 elements to the required location with the appropriate concentration is an important prerequisite. In this context, nanomaterials have proven to be an important class of carriers for delivering elements of the CRISPR-Cas9 systems. However, a series of physiological barriers, from extracellular to intracellular compartments, restrict the delivery of gene editing elements. 60
CRISPR-Cas9 delivery routes, in vivo or ex vivo, may also influence therapeutic genome editing. In the in vivo route, genome editing agents are injected directly into the body through certain administration routes for direct transfection of the editing components. 60 Ex vivo delivery requires the isolation of cells from the animal or human body for in vitro transfection and subsequent administration of the edited cells back into the body. To be considered therapeutic, the dosage of CRISPR-Cas9 must reach the therapeutic window after in vivo delivery. 60,78
There are different methods for RNP delivery, such as physically induced intracellular delivery dependent on disruption of cell membranes to introduce macromolecules. Among common physical approaches, microinjection involves injecting macromolecules into cells and offers high delivery efficiency but can cause damage to cells and requires skilled handling and specialized equipment; electroporation is a technique that uses electrical pulses to introduce macromolecules into cells, and it is generally efficient but can also cause cell damage and requires specialized equipment; microfluidics is a method that uses microscale fluidic channels to deliver macromolecules, can achieve high delivery efficiency, but may require specialized skill and equipment; nanotube spearing is a method that employs nanotubes to pierce cell membranes and introduce macromolecules; biolistics is a technique used mainly in plant cells, which involves the use of microscopic particles to deliver macromolecules; and transformation is mediated by polyethylene glycol. Such physical approaches have limitations and disadvantages associated with mechanisms of action that hinder their in vivo clinical applications. 79 The osmocytosis and propanebetaine-induced transduction (iTOP) method is presented as a new delivery method suitable for native Cas9 proteins and RNPs. However, it is mentioned that iTOP also faces challenges when applied for therapeutic purposes. 79
The iTOP delivery system, developed in 2015, leverages sodium chloride to promote hyperosmolality and incorporates propanebetaine, a compound that enhances cell membrane permeability. The combined effect of hyperosmolality and propanebetaine triggers macropinocytosis, facilitating the intracellular uptake and release of macromolecules. 79,80 This strategy was tested for the delivery of CRISPR-Cas9 RNP components.
In a preliminary test, D’Astolfo and collaborators 80 first prepared KBM7 cells using AAVS1 to deliver a sequence containing the dTomato fluorescent protein out of frame, targeting the AAVS1 sequence. If iTOP works for CRISPR-Cas9 RNP uptake, it should promote a double-strand break at the target site, bringing the dTomato sequence in-frame and resulting in a fluorescent signal. In the first round of CRISPR RNP delivery via iTOP, 30% of KBM7 cells expressed the fluorescent reporter, increasing to 56% in the second round. Similar experiments in H1 human ESCs achieved 10% efficiency in the first round and 26% in the second round.
The authors also validated their system by targeting the endogenous gene DPH7, whose biallelic deletion confers resistance to diphtheria toxin. KBM7 cells were transduced twice with CRISPR RNP via iTOP, and 7 days later, the cell culture was treated with diphtheria toxin for 48 h. Biallelically edited cultures had higher cell survival, while cultures not transduced with RNP or transduced with an RNP complexed with a scramble gRNA had no viable cells. Similar results were obtained in H1 human ESCs.
These experiments demonstrate the potential of iTOP as a delivery system for CRISPR-Cas9 RNP. However, while promising, iTOP requires two rounds of delivery for optimal results and is not suitable for in vivo modifications.
Regarding the development of carriers for RNP delivery, the challenges are related to the large size and positive charge of the Cas9 protein. In this way, the RNP complex can be packaged into virus-like particles (VLP) or extracellular vesicles (EV) using protein engineering. 81,82 VLP has been shown to have efficient genome-editing effects in a variety of primary cells. However, VLP poses a potential risk for immune responses and difficulty in further functionalization. Among nonviral vectors, lipid-based ones are the most investigated for RNP delivery to date and to overcome challenges related to the characteristics of the RNP complex, Cas9 modifications with negatively charged tags can promote stable interactions in nanoparticle formation. That, although it is a crucial step in RNP delivery, this formation is insufficient for successful cellular entry; therefore, multifunctional nanoparticles have been created containing coatings of cationic polymers or lipids to increase stability and promote cellular uptake. 79,83 –86
VLP was developed as a delivery strategy considering the natural capability of viruses to infect cells. With this in mind, the idea was to pack only the cargo to be delivered in the capsid and/or envelope, without the viral genome, and maintain the capability to release the cargo inside the target cell. 87 Many types of viruses can be used, and each of them presents advantages and disadvantages regarding the efficiency of delivery, cargo size, and whether the cargo is going to be integrated or not into the genome (depending on the purpose). More recently in the genome editing context, where it is used tools like CRISPR in different formats of cargo (such as mRNA, DNA plasmid, and RNP), the most widely used capsid type is from lentivirus, which guarantees a more safe and efficient genome editing, because promote a transient delivery of these tools. 88
EVs are defined as lipid vesicles secreted by cells into the extracellular space and are formed from endosomal compartments that fusion to the cell membrane. 89 Currently, the EVs are classified into three groups, microvesicle (0.2–1 um), exosome (30–150 nm), and apoptotic bodies (1–5 um). They are differentiated based on their formation, size, content, function, and release pathway. The cargo loaded into these vesicles could be nucleic acids, such as DNA or RNA, protein, and bioactive lipids. 90
The EVs can also be further optimized for a better outcome of delivery, for example, the membrane of the vesicles can be decorated with other structures providing more stabilization to the particle, or a protein structure to have a specific target. Those modifications can happen before or after the release from the parental cell that produces the EVs. When it is a pre-modification, the parental cells are modified at genomic or metabolic levels. At the same time, post-modifications occur after the collection of EVs and can provide more options of components and ligands, compared to pre-modified. However, pre-modification gives more stable and homogenous vesicles. 91
Regarding the load into the vesicles, there are two ways possible. The pre-isolation load is done with modification in the parental cell with the cargo to be loaded in it, this approach preserves the vesicle form and quality, independent of the cargo being held. However, it has low loading efficiency, and the type of cargo is limited. On the contrary, post-isolation load presents more possibilities. In terms of CRISPR RNP delivery by EVs, it has shown to be a promising method with successful rates of delivery and also works with in vivo delivery, not being limited to in vitro. However, the use of EVs for in vivo delivery faces challenges, such as the potential to trigger the immune system and issues related to the characteristics of the RNP complex. As a result, this method remains limited. 92
THERAPEUTIC GENOME EDITING
SpCas9 is the most frequently used CRISPR-Cas nuclease in basic and clinical research settings. It has been shown that to maximize the efficiency of SpCas9, cytosine is desirable at the DNA cleavage site (position −3 PAM site). 92 Interestingly, Cas9 nuclease isolated from different species, despite structural similarities, can identify different PAM sequences, such as Cas9 from Staphylococcus aureus and Neisseria meningitidis Cas9, require PAM sequences of 5′-NNGRRT-3′ and 5′-NNNRRT-3,′ respectively. 93,94
DNA double-strand breakage is a highly cytotoxic event, provoking a rapid response to DNA damage in a biochemical, temporal, and cell cycle-dependent manner. 93,94 However, many genetic diseases occur due to changes in base pairs in genomic DNA; therefore, the evolution of Cas-based technologies has allowed precise modifications in cellular DNA without the need for DNA double-strand breaks. 95
Base editing
Base editors (BEs) comprise alternative tools to overcome the limitations imposed by DNA double-strand breaks for clinical application in the treatment of genetic diseases. There are two types of DNA base editors: cytosine base editors (CBE), which catalyze the conversion of C•G base pairs to T•A base pairs, and adenine base editors (ABE), which catalyze A•T-to-G•C conversions. 96,97 Direct conversion from one base to another at a programmable target locus without the need for Double Strand Break (DSB) could increase the efficiency of gene correction over HDR without introducing an excess of random indels. 96 Cas9 with mutations that inactivate its catalytic activity maintains its ability to bind to DNA in a manner programmed by the guide RNA, without cleaving the DNA backbone. 98
The base editing strategy comes from understanding R-loop complex formation, where studies show that at least nine nucleotides of the displaced DNA strand are unpaired when the Cas9:gRNA:DNA “R-loop” complex forms. 99 Cas9 undergoes conformational changes following R-loop formation, leading to the activation of its nuclease domains, which is a crucial step in the CRISPR-Cas9 process of gene editing. 17,100,101 Furthermore, the structure of this complex suggests that the first 11 nucleotides of the PAM sequence in the displaced DNA chain are disordered, indicating some degree of flexibility, which makes the complex a potential substrate for editing base. Such speculation indicates that induced mutations of either nuclease domain generates a Cas9 nickase, with cleavage of only one of the DNA strands, while inactivation of both nuclease domains generates a catalytically dead Cas9 (dCas9) that can, however, still bind to specific DNA sequences. 54,102,103
The CBEs use cytidine deaminases to transform cytosines within the R-loop into uracils, which are recognized by polymerases as thymines. 96 ABEs use laboratory-developed deaminases, called TadA*, to transform adenosines within the R-loop into inosines, which are read by polymerases as guanines. 97 The success of editing specific base nucleotides within the R-loop depends on productive interactions between the deaminase and substrate nucleotides. Editing efficiency can be influenced by several factors, including the state of the DNA, such as its chromatin structure, which can differ depending on the specific genomic location or cell type. 96,97
Off-target base editing can occur through guide RNA-dependent or -independent mechanisms. 97,103 –107 Guide RNA-dependent off-target editing arises when the Cas9 domain binds to DNA sites that resemble, but are not identical to, the target locus specified by the guide RNA Strategies to mitigate RNA-dependent off-target base editing guide include introducing mutations into the Cas9 component of BEs to increase DNA specificity by modifying the single guide RNA by adding 5′-guanosine nucleotides or delivering the base editor as a RNP complex. On the contrary, guide RNA-independent off-target editing can occur when the deaminase domain of a base editor binds to bases C or A in a Cas9-independent manner. 108
Great efforts have been made to improve the performance of ABEs by minimizing off-target effects, such as developing versions of TadA by introducing point mutations into TadA or using only a modified TadA, such as ABE max-F148A 7 (109) (an F148A mutation introduced into TadA), ABE max-AW (with TadA E59A and wild-type TadA V106W mutations), and SECURE-ABEs 109 (with eTadA K2aA/R21A or V82G mutation). To improve editing efficiency and targeting scope, two groups of ABE variants were developed through molecular evolution of the TadA monomer, ABE8e, 110 and ABE8s. 111,112
Building on prior research showing that the F148A mutation in TadA effectively nullifies its activity in wild-type Escherichia coli, where TadA functions structurally in base editing, Zhou and colleagues demonstrated that this mutation preserved high efficiency in DNA targeting while minimizing off-target effects. 113
Rees and colleagues 108 introduced E59A inactivators into the TadA or TadA* monomer of ABEmax and showed that this base editor (TadA* E59A) was able to edit the RSL1D1 mRNA at position 152, specifically targeting an adenosine highly analogous to the substrate native tRNAArg of TadA. These findings suggest that both wild-type TadA and TadA* in ABEmax have the ability to deaminate RNA independently of Cas9. To mitigate off-target RNA editing, the wild-type TadA monomer can be rendered inactive; however, residual RNA editing activity persists in the TadA* component, which is necessary to maintain DNA editing functionality. Furthermore, the authors explored substitutions at TadA*’s Val106 in ABEmax(TadA E59A), generating variants with significantly reduced RNA editing while maintaining DNA editing comparable with ABEmax(TadA E59A) and ABEmax. Notably, ABEmax(TadA E59A, TadA* V106W), referred to as ABEmaxAW, emerged as the most efficient DNA base editor and the lowest RNA off-target editing among the Val106 mutants studied. 108
To address the problem of induced RNA base editing independently of guide RNA, Grünewald and colleagues developed SECURE-BE3 (SElective Curbing of Unwanted RNA Editing-BE3) variants, which significantly reduce unintended RNA editing activity. Based on this approach, SECURE-ABE variants were designed using a protein truncation strategy to limit RNA recognition by the optimized ABEmax fusion. Thus, notable variants, K20A/R21A and V82G, exhibited markedly reduced self-editing activities or no detectable self-edits. 109
Considering the evolution of adenine base editors, ABE8e and ABE8s showed very high editing efficiencies in the livers of mice and nonhuman primates 114 or hematopoietic stem cells from patients with sickle cell anemia, 115 demonstrating their potential for gene therapeutics. However, with increasing deamination activity, ABE8e exhibits significant Cas9-independent off-target editing of DNA and RNA. 110,112,115
The effectiveness of base editing occurs within a specific “window of activity,” which primarily involves silent mutations. For some target sites, multiple editable cytosines or adenines exist within or near this activity window. This can lead to the conversion of bases other than the intended target base, known as bystander nucleotides, which can be challenging. Bystander editing is often inconsequential, especially when the goal is to disrupt promoters, splice sites, or other regulatory sequences, or to knock out gene function by introducing premature stop codons. When editing protein-coding genes within the typical 5-base editing window, most cases result in only the desired single amino acid change. This is largely because the genetic code ensures that almost all transitions at the third position of a codon are silent and do not change the amino acid. 103
Eliminating the bystander effects of adenine and cytosine, as well as Cas9-independent editing of ABEs, is crucial, especially for clinical applications; therefore, Chen and collaborators demonstrated precise correction of pathogenic single nucleotide variants, particularly at adenosine homopolymeric sites. Importantly, the observed rates of Cas9-independent RNA and DNA off-target effects are infinitesimally small. 112
Prime editor
The prime editor is a recently developed important CRISPR-Cas-derived genome editing tool. BEs can generate transition point mutations without the need for double-strand breaks; however, they are not capable of generating transversion point mutations (C•G-to-A•T, C•G-to-G•C, T•A-to-A•T, and T•A-G•C), precise insertions, or deletions. Furthermore, BEs have the disadvantage of being able to generate unwanted mutations when there are multiple target nucleotides in the base editing window, and the availability of PAM sometimes prevents targeting specific C or A bases. 103
Aiming to correct genetic variations that contribute to disease in a highly efficient and precise manner while minimizing the production of unwanted by-products, Prime Editing has recently emerged with a sophisticated genome editing approach that allows for the direct insertion of new information genetics at a specific location in the DNA. This method involves using a nickase Cas9 (nCas9), which stands for a Cas9 with one of its active catalytical domains. 38
The nCas9 is fused with a reverse transcriptase enzyme through a linker, and the size of the linker can influence the efficiency. Furthermore, the gRNA to the prime editor is different from the traditional Cas9, is longer, and can be divided into four parts that form the prime editor guide RNA (pegRNA): (1) the gRNA target sequence, (2) primer-binding site, (3) the template of RNA sequence to the reverse transcriptase, and (4) structural RNA sequence for the gRNA. Understanding the pegRNA is vital to knowing how Prime Editor (PE) works. Generally, the gRNA portion first binds to its target, allowing the active domain of Cas9 to clive one of the DNA strands. This action exposes a 3′ or 5′ flap, where the primer-binding site, from pegRNA, has homology. With this, the reverse transcriptase produces a small sequence of DNA, continuous of the original DNA sequence where the primer-binding site is, to be inserted in the cleaved region. Now, the flap contains the edit sequence that will be inserted into the genome or not, through the cell’s natural DNA damage repair. To increase the probability of the edited DNA sequence flap being inserted into the genome, another nicking can be done, in the opposite strand of the modification, within a few base pairs away (around 40 base pairs or more) from where the editing is happening, for the edited strand serves as the template model for the repair. 38
Four main types of editors were developed: PE1, PE2, PE3, and PE3b, with different levels of efficiency in editing human cells, with better efficiencies being observed in PE2, PE3, and PE3b depending on the experimental conditions and target sequences of interest. 38 Considering that both PE3 and PE3b are composed of PE2 and an additional gRNA, the efficiency of PE2 on a given target sequence may also affect the efficiencies of PE3 and PE3b on the target. 38,116 Thus, information about factors affecting PE2 efficiency such as those reported by Kim et al., 116 through high-throughput analyses, can facilitate master editing. Factors include (1) using a 13-nt Reverse Transcriptase (RT) primer binding site and a 12-nt RT template; (2) high GC count in the region of the RT initiation site, if possible; (3) use a G in the last templated nucleotide when the RT template length is <12 nt; and (4) PAM editing, which, together with the use of computational models (DeepPE, PE_type, and PE_position), allows the editing of small genetic mutations in a very efficient way, without the use of donor DNAs. 117
The effective delivery of base editing and prime editing components, both in vitro and in vivo, poses a significant challenge. In terms of relevance, delivery methods for basic and mainstream editing share common goals of achieving efficient and precise genetic modifications in target cells. However, delivering prime editing presents unique challenges due to its distinct mechanism and additional components required for editing. As prime editing continues to advance, optimizing delivery methods will be crucial for its broad application in research and clinical settings. 118
Delivery methods for basic editing tools such as CRISPR-Cas9 have been extensively studied and optimized. These methods typically involve the delivery of the Cas9 protein and guiding RNA into target cells using viral and nonviral vectors. For prime editing, realizing its potential in mammalian research and therapeutic applications necessitates safe and efficient delivery methods capable of transporting prime editors to tissues in vivo. Although some progress has been made, with several groups reporting in vivo delivery via viral methods such as adenovirus and adeno-associated virus (AAV), these methods come with limitations. 119 –125 Viral delivery requires encoding the transgene directly into the viral gene expression cassette, which restricts the size of the transgene. AAV, in particular, has a cargo gene size limit of about 4.7 kb, leading to the fragmentation of larger cargos like PEs into multiple AAVs. 126 This fragmentation can impair editing efficiency, especially at moderate or low vector doses. 125,127 Furthermore, viral delivery methods present safety concerns, including increased off-target editing due to sustained transgene expression and the potential integration of unwanted DNA cargo into host cell genomes. 128
Nonviral delivery approaches, such as CPPs, show promise and may offer alternatives to overcome some of these limitations. Continued research and development in delivery methods are essential for unlocking the full potential of prime editing in various applications.
CONCLUSION
For effective genomic editing, considering the delivery of the CRISPR-Cas9 component within the cell, direct delivery of the RNP complex offers several advantages, as it is fast; it does not depend on transcription, translation, and subsequent assembly of the active complex within the cell; and due to its transient nature, which reduces the chance of off-target cutting events and low risk of insertional mutagenesis, which is often associated with viral vectors. CPPs are versatile tools for forming complexes and have been shown to be effective in delivering various biomolecules, including RNPs for genome editing in human cells, especially the chimeric peptides which have better penetrance to the target cell and a greater endosomal escape. This effectiveness arises from its ability to form noncovalent complexes with varying charges and covalent conjugations with RNA and proteins. This delivery method combined with base editing and primer editing systems, tools known for their simplicity and precision, has the great potential for correcting point mutations in human genetic diseases. Ongoing research efforts aim to improve BEs and prime editors by increasing efficiency, expanding the scope of targeting, and ensuring the purity of edited products. Despite the existing challenges, these gene editing approaches show a huge potential for future clinical applications in the treatment of single-mutation genetic diseases.
Footnotes
ACKNOWLEDGMENT
The authors wish to thank the Brazilian Ministry of Health and Public Policies and Health Care Models Program (PMA-FIOCRUZ) for support for the development of research.
AUTHORS’ CONTRIBUTIONS
C.C.P.D.L.D.M.: Conception and design, article writing, and final approval of the article; E.M.C.: Design, article writing, and final approval of the article; M.H.B.: Design, article writing, and final approval of the article; Z.F.M.D.V.: Design and final approval of the article and financial support.
AUTHOR DISCLOSURE
The authors declare no conflicts of interest.
FUNDING INFORMATION
Public Policies and Health Care Models Program (PMA-FIOCRUZ) by funding source (VPAAPS-002-FIO15-2-1).