
Abstract
Double-stranded break (DSB) repair of eukaryotic DNA is mainly accomplished by nonhomologous end joining and homologous recombination (HR). Providing exogenous templates during HR repair can result in the editing of target genes, which is the central mechanism of the well-established clustered regularly interspaced short palindromic repeats (CRISPR) gene editing system. Currently, exogenous templates are mainly DNA molecules, which can provoke a cellular immune response within the cell. In order to verify the feasibility of RNA molecules as repair templates for HR in mammalian cell genome editing, we fused RNA template molecules to the 3′-end of single guide RNA (sgRNA), so that the sgRNA and the homologous template RNA form a single RNA molecule. The results show this construct can be used as a repair template to achieve target gene editing in mammalian cells. In addition, the factors influencing HR mediated by RNA template molecules were investigated, and it was found that increasing the length of homologous arms and inducing an R-loop near the DSBcan effectively promote HR repair. Furthermore, intracellular homologous chromosomes may compete with exogenous RNA templates. The findings in this article provide a reference for the utilization of RNA template molecules to mediate target gene editing in eukaryotic cells, as well as a basis for the study of the mechanism by which RNA molecules mediate the repair of DSBs.
INTRODUCTION
CRISPR/Cas9 is well established as an important genome editing tool in eukaryotic cells. This system generates double-stranded breaks (DSBs) at a target site followed by homologous recombination (HR) if a homologous template exists, resulting in directed base substitutions in the target gene. 1 –5 Currently, templates used for HR mainly include linear double-stranded DNA, circular double-stranded DNA, and single-stranded DNA in three forms. 6 Comparison of the efficiency of HR mediated by the three forms of homologous templates showed that the integration efficiency of single-stranded DNA is significantly better than that of the other two. 6 Furthermore, the addition of sgRNA recognition sequences at both ends of the circular double-stranded DNA template, which is targeted and recognized by Cas9 in the cell and cleaved to release the linear DNA template, 7 can effectively improve the HR efficiency. Significant progress has been made in addressing the factors that affect the HR efficiency of DNA templates. For example, inhibiting the error-prone nonhomologous end joining, thus promoting accurate homology-directed repair (HDR), 8 –16 providing a favorable cellular environment for HDR repair (synchronizing the cell cycle in the S/G2 phase 17 –19 ) or co-localizing HR templates near DSBs 7,20 –24 can effectively increase the efficiency of HR.
The development of RNA vaccines is progressing rapidly. Karikó et al. found that the primary reason for the immune response triggered by exogenous RNA is the attachment method of uracil (U) to the ribose. They demonstrated that substituting uridine with pseudouridine (Ψ) in RNA significantly reduces the intracellular immune response. 25,26 Therefore, compared with DNA molecules, using base-modified RNA molecules (with uridine replaced by pseudouridine) as templates can effectively mitigate the intracellular immune response. DNA templates are time-consuming and labor-intensive to prepare. Furthermore, it requires simultaneous transfection of both the CRISPR system and DNA templates into the same target cell, which is inefficient when using DNA templates. Therefore, researchers have begun to explore the feasibility of RNA as a template to mediate HR. However, research on whether RNA molecules can be used as templates to mediate the realization of HR is still in its infancy. The few studies that have been conducted have found that RNA molecules can be used as templates for HR repair and can mediate the repair of target genes. For example, RNA molecules can be used as repair templates for HR in yeast. 27 In higher plants (rice cells), a HR efficiency of nearly 1% can be achieved using ssRNA as a template. 28
To investigate the use of RNA as a repair template to mediate HR in mammalian cells, we fused the homologous template RNA molecule to the 3′-end of sgRNA, so that the sgRNA and the homologous template RNA were transcribed into a single RNA molecule. The homologous template RNA molecule was transported to the vicinity of DSBs through the formation of a complex with Cas9 proteins to achieve the insertion of the exogenous short fragments (HR efficiency of about 1%) at the target genes. While this is lower than single-stranded DNA homology template-mediated recombination efficiencies of more than 10%, this does indicate that RNA molecules can also serve as a HR template in mammalian cells. In addition, we mapped and optimized the conditions for RNA template molecules to mediate HR. This included changing the length of homologous arms, analyzing intracellular homologous chromosome interference, and inducing the formation of an R-loop near the DSB. Increasing the length of homologous arms and inducing an R-loop near the DSB were both found to effectively promote HR repair. However, homologous chromosomes may compete with exogenous RNA templates. In conclusion, we found that RNA can be used as a template to mediate HR in mammalian cells, suggesting that genome editing can be achieved using either RNA templates or DNA templates.
MATERIALS AND METHODS
Construction of the CRISPR-Cas9 editing system with RNA templates
Design of donor_RNA with different length homology arms (55, 100, 200, and 400 nt). The designed donor template (Supplementary Table) was fused from two predesigned short fragments and transcribed in vitro using the sgRNA Enzyme Mix reaction kit. The donor has a region of 20 nt base complementary pairing with target-specific sgRNA at one end. The primers for plasmid construction are shown in (Supplementary Table).
In vitro RNA transcription
Using the EnGen® sgRNA Synthesis Kit, PCR assembly reactions were set up in 20 μL volumes by adding the reaction components in the given sequence: 2 μL nuclease-free water, 10 μL EnGen 2× sgRNA Reaction Mix, Streptococcus pyogenes, 5 μL target-specific DNA oligo (1 μM), 1 μL DTT (0.1M) and 2 μL EnGen sgRNA enzyme mix. Reactions were mixed thoroughly and pulse spun in a microcentrifuge prior to incubation at 37°C for 30 min followed by transfer of the reaction to ice and treatment addition of 30 μL of nuclease-free water to bring the volume to 50 μL. Then 2 μL of deoxyribonuclease I was added, and the reaction was mixed and incubated at 37°C for 15 min. RNA purification and gel electrophoresis analysis were then performed.
Inverted fluorescence microscopy and flow cytometry
At the 60 h time point, the fluorescence intensity of enhanced green fluorescent protein (EGFP) was detected by flow cytometry (Beckman), and the average value of three experimental results was calculated. The statistical differences in transfection efficiency of each group were calculated using SPSS 20.0 (IBM) and GraphPad Prism (GraphPad Software) software.
Prediction of DNA regions prone to R-loop formation
Two R-loop formation prediction applications, rlooper and rlooperplusstats, were used in GitHub to simultaneously predict and analyze the R-loop-prone regions of the left and right fragments of the target gene with an average total length of 1,200 bp, including the target site.
The computer-predicted R-loop-prone region closer to the protospacer adjacent motif (PAM) site was in the 100–300 bp region at the 5′ end of PAM. This region is rich in GC bases and is in a transcriptionally active region. Based on literature reports, we fused a segment at the 5′ end of sgRNA that readily induces the generation of R-loop structures with the left homology arm extending into the R-loop-inducing region. A segment from the mCherry gene was inserted between the R-loop induced structure and the sgRNA_RNA, and sequence comparison was performed to ensure that there was no contiguous base complementary pairing sequence with the human genome to serve as the linker. The synthetic constructs R-loop-sgRNA-HR (R-loop induced) and sgRNA-HR (no R-loop induced) were commissioned from IGE BIO LTD.
Genome extraction
Genomic DNA of HEK 293T cells and Jurkat cells was extracted as previously described using the TaKaRa MiniBEST Universal Genomic DNA Extraction Kit Ver.5.0.
Preparation of libraries and deep sequencing
Next-generation sequencing primers for the target region were designed in the 200–300 bp size range (Supplementary Table). Primers were not permitted to reside within a 50-nt region on either side of the expected sgRNA cut site to ensure coverage of varied deletion sizes. A two-step PCR fusion method was performed to attach bar-coded sequencing adaptors for multiplex deep sequencing. For experiments with long homology arms, we used a three-step PCR method to amplify the 1,000 bp end of the slice before performing a two-step PCR fusion method to connect the sequencing adapters. Sequencing of the libraries was completed with an Illumina NovaSeq. The percentage of indels and substitutions was analyzed using CRISPResso 2. We homogenized HR efficiency to reduce the effect of transfection efficiency, that is, the HR efficiency of the target region = (HR) %/(indels + substitutions) %.
Statistical analysis
The data reported here are means ± standard error of three independent culture experiments. Statistical comparisons were performed using the one-way analysis of variance. A p value of < 0.05 was considered statistically significant.
RESULTS
RNA homology templates mediate exogenous fragment insertion of target genes in HEK 293T cells
In order to verify the possibility that sgRNA carries RNA templates to the vicinity of target genes and realizes target gene modification, we first transcribed and synthesized donor RNA (both left and right homology arms 100 bp in length) and sgRNA in vitro, and then electrotransfected HEK-293T cells with this complex. This construct has a 20-base complementarity between the 3′ end of donor RNA and sgRNA, and an ribonucleoprotein (RNP) complex can be formed by incubation of Cas9 protein, donor RNA, and sgRNA in vitro. The Cas9 protein generates DSBs by sgRNA-mediated cleavage of the target site, and the RNA template is translocated to the vicinity of the DSBs by interbinding with the sgRNA (Fig. 1a). The insertion sequence carrying eight bases in the donor RNA is depicted. After 72 h of transfection, the cell genome was extracted and the target sites were amplified for library construction, and the editing of target sites was analyzed by next-generation sequencing. As shown in Fig. 1b, the HR efficiency is 1.73%. Thus, preliminary indications suggest that RNA templates can mediate HR repair of DSBs in mammalian cells. We next explored the fusion of donor RNA to the 3′ end of sgRNA (Fig. 1c), thereby forming an RNA molecule that can mediate the modification of target genes. Due to the tediousness of in vitro transcription of donor RNA and sgRNA, as well as in vitro incubation to form RNP operators, we continued our exploration using plasmid transfection. We found that the efficiency of HR mediated by fusing donor RNA with left and right homology arm lengths of 100 bp to the 3′ end of sgRNA was 1.06%, and the efficiency of homologous template-mediated HR by cotransfection of single-stranded DNA was 10.88% (Fig. 1d). Meanwhile, we found that RNA homology templates could achieve HR in both HEK 293T cells and Jurakt cells without significant differences in efficiency (Supplementary Fig. S1). The integration of epitope peptide sequences at target sites has important applications in tumor immunotherapy. To validate the versatility of RNA template-mediated HR repair, we examined the insertion of a 27 bp fragment (derived from the human hepatitis B virus antigenic epitope) into the EMX1 locus of the target gene. This resulted in an integration efficiency of 4.46% for the 27 bp fragment, as shown in Fig. 1e. These results suggest that sgRNA fusion RNA templates can achieve HR and mediate DSB repair; however, the recombination efficiency is low and needs to be further improved.

Homologous template RNA fused to the 3′ end of sgRNA mediates HR repair of DSBs.
Increasing homology arm length effectively enhances the efficiency of RNA molecule-mediated HR
To further improve the efficiency of RNA template-mediated HR, we next explored the factors influencing this process. Previous studies on DNA template-mediated HR revealed that homology arm length has a significant effect on HR efficiency. 7,29 –32 Therefore, we first explored the effect of varying the length of the homology arm on the efficiency of RNA template-mediated HR.
Using a 100 bp length homology arm as a control, we shortened the homology arm length to 55 bp, or increased the homology arm to 200 or 400 bp to study the effect of different lengths of homology arms on the efficiency of HR mediated by RNA templates (Fig. 2a). Increasing homology arm length increases HR efficiency from 1.02% at 100 bp to 2.63% at 200 bp (a 2.59-fold increase) and 5.30% at 400 bp (a 5.21-fold increase) (Fig. 2b). We also analyzed imperfect HR (where the integration of the target fragment occurs at the target site, but with a 1–2 base mutation). As shown in Supplementary Fig. S2A, imperfect HR could occur, and an increase in homology arm length did not affect the efficiency of imperfect HR (Supplementary Fig. S2B). In addition, we found that an increase in homology arm length did not significantly affect the Indel ratio at the target site. However, a decrease in the Indel ratio was observed when the homology arm length was 400 bp (Supplementary Fig. S2C).

Effect of homology arm length on the efficiency of HR mediated by homologous template RNA fusion at the 3′ end of sgRNA. Schematic diagram of the construction of homologous recombinant vectors with homologous arms of different lengths
To investigate the different modifications mediated by this approach to the target genes, we also used RNA HR repair to induce base mutations at the EMX1 locus (Fig. 2c). We found that the HR repair efficiency increased significantly with the increasing homology arm length, from 4.72% at 100 bp to 10.85% at 200 bp and 20.75% at 400 bp (Fig. 2d). These results suggest that, similar to DNA template-mediated HR, increasing the homology arm length can also improve the efficiency of RNA-mediated HR.
Induction of an R-loop in the vicinity of DSB effectively promotes RNA molecule-mediated HR repair
R-loops are three-stranded DNA/RNA hybrid structures 33 that exist during gene transcription. Previous findings have shown that after DSB occurs in DNA, there is an R-loop near the DSB, which recruits HR repair-associated proteins to the vicinity of the R-loop. 34 Thus, we wanted to determine if RNA molecule-mediated HR repair would be facilitated by R-loop formation in the vicinity of Cas9-mediated generation of DSBs. First, we used R-loop formation prediction software (rlooper and rlooperplusstats) to predict and analyze the R-loop-prone region at the DSB of the target gene (PD1 gene and EMX1 gene) in the range of 600 bp in length at both ends of the gene. The results showed that the R-loop was prone to occur in the range of 100–300 bp from the 5′ end of the PAM site (Supplementary Fig. S3), and the GC content of the bases in this region was also consistent with the GC base-rich feature.
R-loops are three-stranded DNA/RNA hybrid structures 33 that are present during gene transcription. Previous results have shown that after a double-strand break (DSB) occurs in DNA, an R-loop forms near the DSB and recruits HR repair-associated proteins to the vicinity of the R-loop. 34 Therefore, we aimed to determine whether Cas9-mediated R-loop formation near the DSB promotes RNA molecule-mediated HR repair. First, we predicted and analyzed the R-loop-prone regions at the DSB within 600 bp of the ends of the target genes (PD1 and EMX1) using R-loop formation prediction software (rlooper and rlooperplusstats). The results showed that R-loops are prone to appear in the range of 100–300 bp from the 5′ end of the PAM site (Supplementary Fig. S3), and the GC content of the bases in this region is consistent with the GC-rich feature.
We fused the R-loop-inducing sequence to the 5′ end of the sgRNA to construct the HR system (R-loop-sgRNA-HR) (Fig. 3a), with a piece of an unrelated sequence originating from mCherry between the R-loop-inducing sequence and the 5′ end of the sgRNA. We also constructed editing systems that do not contain inducible sequences as controls. For the PD1 site (Fig. 3b), the recombination efficiency with R-loop-inducing sequences was markedly increased to 34.18%, which was significantly higher than that of the co-transformed single-stranded DNA group (15.66%), while the control group without R-loop-inducing sequences was only 3.14%. For the EMX1 site (Fig. 3c), the recombination efficiency with R-loop-inducing sequences was markedly increased to 25.14%, while the control group without R-loop-inducing sequences was only 5.24%. These results suggest that induction of the R-loop in the vicinity of DSBs can effectively promote HR repair mediated by RNA molecules.
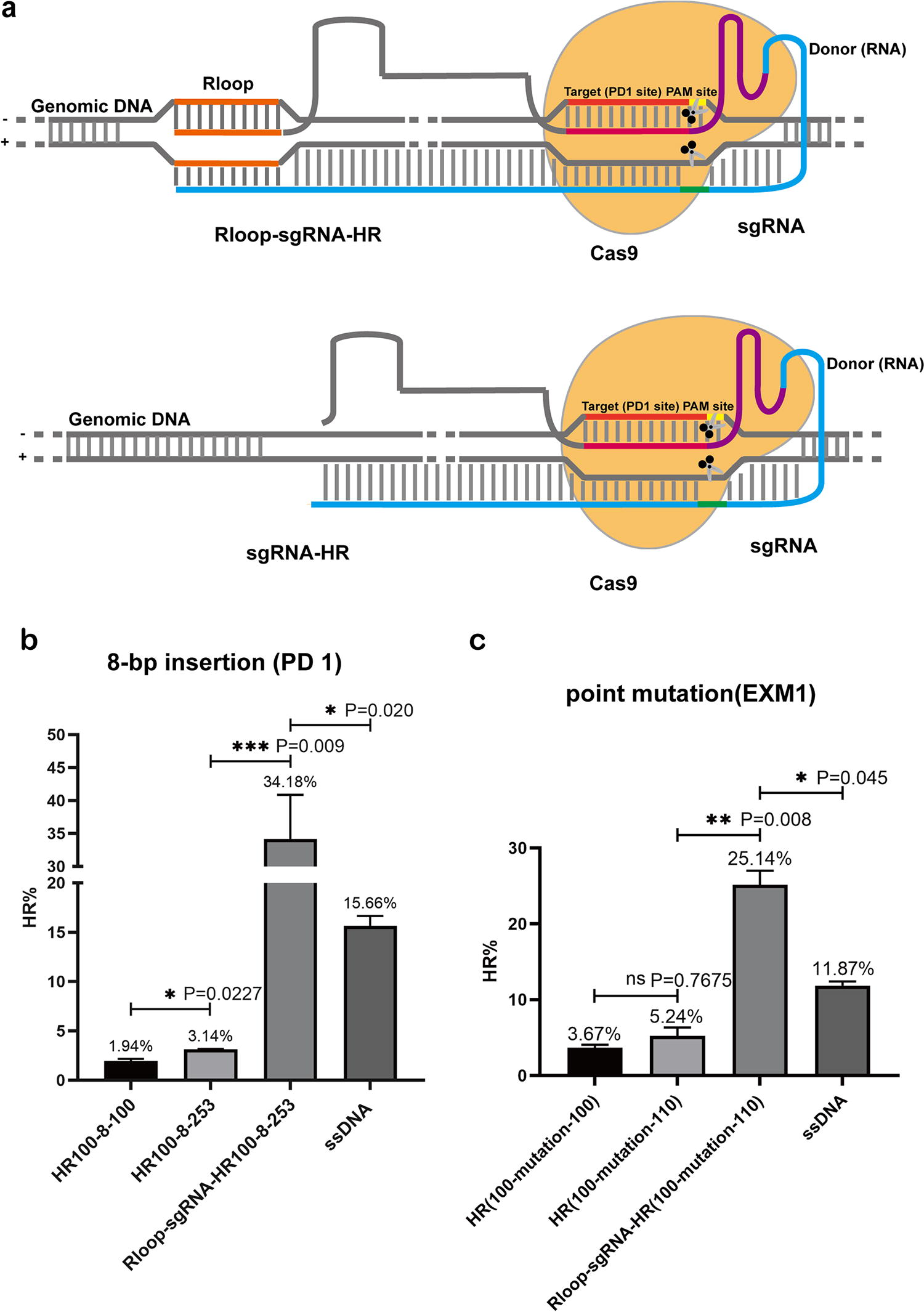
Effect of inducing R-loop near DSB on the efficiency of RNA template-mediated HR.
Homologous chromosomes can serve as repair templates and influence RNA molecule-mediated homologous recombination
Previous studies have shown that when DSBs are produced in target genes, homologous chromosomes can act as homologous templates to participate in the repair of DSBs if no exogenous homologous template exists. 35 –37 We next wanted to determine if homologous chromosomes can influence RNA template-mediated HR. To this end, we selected the X chromosome G6PD gene and the autosomal PD1 gene as target sites to analyze the efficiency of RNA molecule-mediated HR in Jurkat cells (male-derived cells) and HEK 293T cells (female-derived cells), respectively (Fig. 4a). As shown in Fig. 4b, the efficiency of HR targeting the G6PD locus was significantly higher in Jurkat cells than in HEK 293T cells. When the homology arm length was 100 bp (G6PD gRNA-HR[100-8-100]), the recombination efficiency was 6.02% in Jurkat cells and 1.29% in HEK 293T cells. As for the PD1 locus, when the homology arm length was 100 bp (PD1 gRNA-HR[100-8-100]), the recombination efficiency was not significantly different between HEK 293T and Jurkat cells. These results suggest that homologous chromosomes can compete with exogenous RNA templates. Thus, the effect of homologous chromosomes as homologous templates should be taken into account when using RNA molecules to mediate HR.
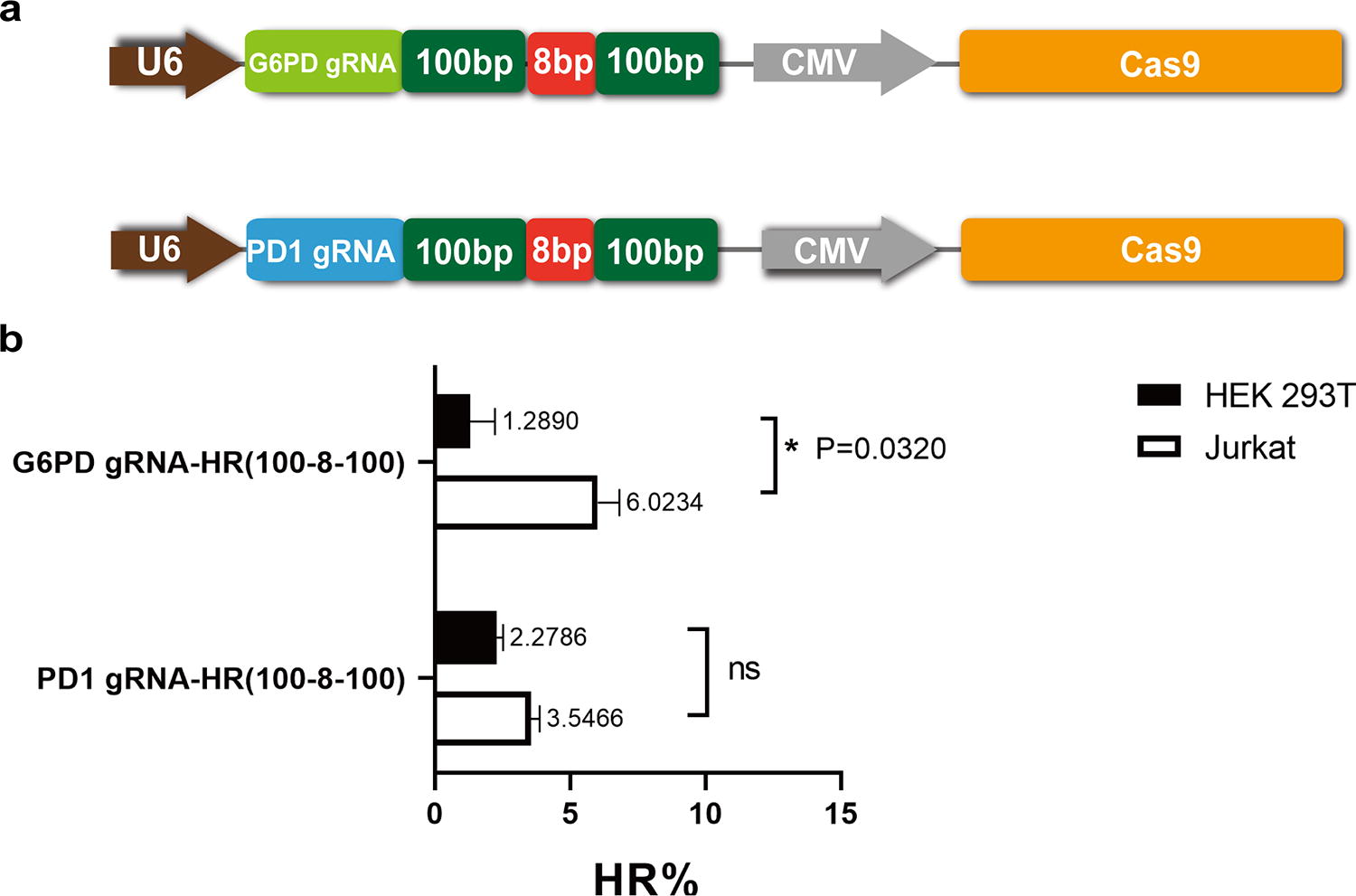
Effect of intracellular homologous chromosomes on the efficiency of RNA template-mediated HR.
DISCUSSION
DNA templates are time-consuming and labor-intensive to prepare, as well as it is inefficient to simultaneously transfect both the CRISPR system and DNA templates into the same target cell. Therefore, researchers began to explore the feasibility of RNA as a template to mediate HR. In this article, we verified that RNA can serve as a repair template to mediate HR repair in mammalian cells, as well as confirming that RNA can be used as a HR repair template in yeast 27 and rice. 28 Therefore, RNA molecules show promise for use as repair templates to mediate HR repair in eukaryotic cells. When using HR for genome editing, in addition to traditional DNA templates, RNA molecules can also be considered.
Previous studies have shown that when DNA is used as a template for HR repair, the longer the homology arm is (within a certain range), the more efficient the HR. 7 In this article, similar results were found when RNA was used as homology template repair, but because the RNA template was fused to the 3′ end of sgRNA, the length of the homology arm was limited due to potential effects on the cutting efficiency of Cas9. In a previous study on prime editor, Anzalone et al. found that even if nickase Cas9 was fused to inactive reverse transcriptase, base modification of the target site could still be achieved in eukaryotic cells, albeit with very low efficiency. 38 In conjunction with this article, it was found that RNA templates containing only one homology arm could also mediate HR repair (Supplementary Fig. S4), albeit with a recombination efficiency of 0.08%. These results imply that reverse transcriptase present in eukaryotic cells could be involved in this HR process.
Previous studies have found that homologous chromosomes can participate in DSB repair as homology templates. In this article, we find that homologous chromosomes can compete with exogenous RNA homology templates and reduce the efficiency of RNA molecule-mediated HR (Fig. 4). Steven et al. found that synchronizing the cell cycle to M phase effectively increases the efficiency of DNA template-mediated HR, although the exact mechanism remains unclear. 18 In the M phase of the cell cycle, the two sister chromatids are separated and pulled by the spindle fiber to the poles of the cell. As a result, the homologous chromosomes are farther away from each other and presumably interfere less with the exogenous templates. Thus, for RNA molecule-mediated HR repair, synchronization of the cell cycle to the M phase followed by the introduction of exogenous RNA templates and Cas9 proteins might also increase recombination efficiency.
Previous studies on RNA molecules mediating DSB repair in yeast cells have revealed the presence of R-loops in the vicinity of DSBs, which can recruit DSB repair proteins and thus promote DSB repair, while inhibiting the formation of these R-loops decreases the repair efficiency. 39,40 This phenomenon suggests that artificially inducing R-loop formation in the vicinity of the DSB might increase the efficiency of HR. In fact, we found that induction of an R-loop in the vicinity of DSB could effectively improve the efficiency of RNA-mediated HR in mammalian cells (Fig. 3b and 3c). Future studies will be needed to identify the specific protein factors involved in this repair pathway in mammalian cells. This could be accomplished through proximity labeling or RNA coprecipitation techniques. Moreover, directed localization of these proteins to the vicinity of the DSBs could be used to further improve the efficiency of HR. In addition, in this study, we primarily used plasmid-encoded RNA molecules. For future applications, we can use in vitro transcribed RNAs as HR templates. Considering the length limitation of chemically synthesized RNAs, we can transcribe sgRNA molecules fused with homologous templates in vitro, substitute uridine with pseudouridine, and then transfect the cells.
CONCLUSIONS
HR repair after Cas9-mediated DSB production was achieved by fusing the RNA homology template at the 3′ end of sgRNA. Furthermore, increasing the length of the homology arm and inducing the R-loop in the vicinity of the DSB could effectively improve the efficiency of HR. Thus, these findings may contribute not only to enriching the toolbox of CRISPR-based genome editing but may also provide a useful reference for the study of HR mechanisms based on RNA molecules.
Footnotes
Acknowledgments
This work was supported by the Guangdong Basic and Applied Basic Research Foundation (2018A030313860, 2020A1515010889, 2018A030313114), the Guangzhou Science and Technology Project (202002030477), the Guangdong Project of Graduate Education Innovation (2024SFKC_058), and the Innovation Team project in regular colleges and universities of Guangdong Province (Natural Science) (2023KCXTD021).
AUTHORS’ CONTRIBUTIONS
Conceiving, designing, and supervising the project: W.Z. and H.S. Writing: Y.W. Conducting the experiments: Y.W., H.W., M.L., X.L., N.D., and H.L.
AUTHOR DISCLOSURE
No competing financial interests exist.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.