
Abstract
Adeno-associated virus-mediated gene therapies for certain muscle disorders require regulatory cassettes that provide high-level, striated muscle-specific activity. However, cardiotoxicity has emerged as a serious concern in clinical trials for Duchenne muscular dystrophy and X-linked myotubular myopathy. While this may be caused by systemic inflammatory effects of the treatment, high transgene expression in the heart may also play a role. Thus, certain muscle disorders may require a modulated level of therapeutic expression in the heart, while others may not require any cardiac expression at all. Additionally, the size of some cargos requires regulatory cassettes to be small enough that large cDNAs and other therapeutic payloads can be accommodated. Thus, we have performed enhancer/promoter optimization to develop highly minimized regulatory cassettes that are active in skeletal muscles, with either low or no detectable activity in cardiac muscle. Our No-heart (NH) cassette is active in most skeletal muscles, but exhibits only very low activity in extensor digitorum longus (EDL), soleus, and diaphragm, and no activity in the heart. By contrast, our Have a Little Heart (HLH) cassette displays high activity in most skeletal muscles, comparable to the ∼800-bp CK8 cassette, with increased activity in EDL, soleus, and diaphragm, and low activity in the heart. Due to their small size, these cassettes can be used in therapeutic strategies with both flexible (e.g., antisense) and stringent (e.g., CRISPR/Cas or bicistronic) size limitations. Thus, our new cassettes may be useful for gene therapies of muscle disorders in which the need for low or almost no expression in cardiac muscle would outweigh the need for high levels of therapeutic product in certain skeletal muscles.
INTRODUCTION
Gene therapy approaches in preclinical models of muscle disease can result in cardiotoxicity caused by cardiac expression of the transgene, as in the calpain 3-deficient mouse model of limb–girdle muscular dystrophy type 2A (LGMD 2A/LGMD R1). 1,2 While this can be prevented with the use of muscle-specific regulatory cassettes in conjunction with miRNA detargeting sequences, 1,2 these increase the size of therapeutic cassettes containing cDNAs that may be close to the stringent packaging capacity of recombinant adeno-associated virus (AAV) vectors. Of even greater concern are the severe adverse events reported in human gene therapy trials for two muscle diseases. 3 –6 Deaths in the ASPIRO X-linked myotubular myopathy (XLMTM) clinical trial (ClinicalTrials.gov Identifier: NCT03199469) were due to liver failure, but several patients had laboratory findings indicative of myocarditis, and in a Pfizer trial for Duchenne Muscular Dystrophy (DMD) (ClinicalTrials.gov Identifier: NCT03362502), two instances of myocarditis were reported. Thus, there is a clinical need for minimized gene regulatory cassettes with appropriate activity in all skeletal muscles, but low or no activity in the heart.
In this study, we describe two Muscle Creatine Kinase (Ckm)-based regulatory cassettes that begin to address this need: (1) No Heart (NH), our previously reported cassette that is moderately active in most fast-twitch skeletal muscles, but essentially inactive in soleus (slow-twitch muscle), extensor digitorum longus (EDL), diaphragm, and heart, 7 and (2) Have a Little Heart (HLH), a cassette that displays high activity in most fast-twitch muscles, with increased activity in EDL, soleus, and diaphragm, and low activity in the heart (Fig. 1). Thus, the NH cassette may be useful for skeletal muscle indications where no cardiac expression of the transgene is needed (providing low-to-moderate expression in skeletal muscles is sufficient), whereas the HLH cassette should be useful for indications requiring moderate-to-high transgene expression in skeletal muscles and low expression in the heart, thus avoiding potential transgene-mediated cardiotoxicity. Additionally, these are by far the smallest cassettes that use three copies of a modified Ckm enhancer, and they are ∼14% smaller than even the minimal CK8e cassette, which contains one modified Ckm enhancer and a modified proximal promoter that increases both skeletal and cardiac muscle expression. 10 –17 As such, they may be useful for approaches that require large therapeutic cargos, such as CRISPR/Cas-based expression of effector proteins and polycistronic vectors. Altogether, these regulatory cassettes may be applicable to diverse gene therapy strategies targeting a range of myopathies.

Evolution of regulatory cassette platform. Schematic of several Muscle Creatine Kinase (Ckm)-based muscle regulatory cassettes and our two new cassettes based on the CK8 design. The seven known transcriptional regulatory elements within the Ckm enhancer are labeled. Changes from the endogenous sequence and/or previous Ckm-based cassettes are shown in blue. For CK8, these include: 1) deletion of 63 bp between the Right E-box and the MEF2 site; 2) 3 tandem copies of the modified enhancer; 3) mutation of sequence overlapping +1 to a consensus Initiator element (Inr); and 4) inclusion of +50 sequence downstream of the transcription start site. 8,9 The commonly used 438-bp CK8e cassette 10 –17 contains the following modifications from CK8: 1) only a single copy of a modified Ckm enhancer with small sequence deletions 5’ of the CArG site and 3’ of the MEF2 site and 2) −269 proximal promoter sequence used. The NH cassette 7 contains the following modifications from CK8: 1) CArG and AP2 sites removed; 2) sequence between all regulatory elements minimized; 3) Left E-box mutated to a Right E-box; 18,19 and 4) −80 basal promoter sequence used. 20,21 The HLH cassette contains the following modification from the NH cassette: additional Right E-box replaced with the original Left E-box. HLH, Have a Little Heart; NH, No Heart.
MATERIALS AND METHODS
Plasmids
The NH cassette 7 consists of three copies of a modified Ckm (mouse) enhancer in tandem, upstream of an 80-bp basal Ckm promoter. Main features are as follows: (1) Left E-box mutated to Right E-box; 18,19 (2) enhancer CArG and AP2 sites removed; (3) 63 bp between Right E-box and MEF2 site removed; 8 (4) sequence between transcription factor-binding motifs minimized; (5) −80, 20,21 to +50, 8 promoter sequence used; and (6) consensus Initiator element (Inr) added. 8 The HLH cassette is based on the NH cassette, with the following modification: the additional Right E-box was replaced with the original Left E-box from the Ckm enhancer. CK8 is the ∼800-bp version originally described. 8,9 For all cassettes, the mCherry reporter was inserted downstream of the transcription start site. Reporter cassettes were synthesized and fully sequenced by GenScript. For evaluation of cassette activities in vivo, each regulatory cassette driving the mCherry reporter was cloned between the AAV2 inverted terminal repeats (ITRs) (using MluI and RsrII) of the pAAV-CA plasmid, 22 a gift from Naoshige Uchida (Addgene plasmid # 69616; http://n2t.net/addgene:69616; RRID:Addgene_69616). Recombinant AAV9(Y731F) particles were produced, purified, and titered using quantitative real-time PCR to determine vector genomes (vg)/mL by Vector Biolabs. Vector sequences are available upon request.
Animals
All animal experiments were approved by the Institutional Animal Care and Use Committee of the University of Nevada, Reno (IACUC protocol #22-08-1175). Wild-type C57BL/6J mice (3 per vector) were weighed and given systemic retro-orbital injections (ROI) with AAV9(Y731F) (2.6 × 1014 vg/kg) containing the NH, HLH, or CK8 cassette regulating expression of the mCherry reporter. Due to practical constraints, male mice were used for NH and HLH, and female mice were used for CK8. Potential effects of sex differences on Ckm transcriptional regulation were addressed using qRT-PCR analysis of endogenous gene expression, as described below. Mice were anesthetized with 3.5% isoflurane before and during injection. Mice were injected at 3 weeks of age and sampled 12 weeks postinjection.
Whole-body optical imaging
The fur of the ventral abdominal area was removed with depilatory cream before imaging. Fluorescent images of mCherry in live mice under anesthesia (2%-isoflurane) were captured weekly for 10 weeks after ROI using the IVIS Lumina III and Living Image software ver. 4.3.1 (PerkinElmer) with λex = 580 nm/λem = 620 nm filters, exposure time of 1 s, 12.5 × 12.5-cm FOV, f/stop 4. All images were processed in Living Image equally with an epifluorescence scale of min = 7 × 108 and max = 3.2 × 1010, smoothing = 5 × 5, and binning = 2. The total flux (photons/second) and maximum radiance (photons/second/cm2/steradian) were measured using Living Image software.
Ex vivo optical imaging
At 12 weeks postinjection, all mice were euthanized, tissues were dissected, and mCherry signals in whole or partial tissues were captured with the Leica THUNDER/DFC-7000T fluorescent imaging system and Leica LAS X 3.4.1 software, using the same exposure time, unless indicated in the figure. Images were assembled with Adobe Photoshop 24.1.0, and exposures were adjusted equally. The fluorescent signals in selected tissues were further measured using IVIS Lumina III and Living Image software with filters λex = 580 nm/λem = 620 nm, exposure time 0.5 s, FOV 4 × 4-cm, and f/stop 8. The total flux (photons/second) and maximum radiance (photons/second/cm2/steradian) were measured using Living Image software.
Immunofluorescence of tissues
Skeletal muscles and heart were coated with O.C.T. compound (Tissue-Tek) and snap frozen in liquid nitrogen-chilled isopentane immediately after dissection. Half of the diaphragm was embedded in 18% sucrose/40% O.C.T. and frozen in liquid nitrogen-chilled isopentane. Tissues were sectioned using Leica cryostat CM1950 at 10 µm thickness, and fixed immediately with formaldehyde vapor for 20 min. Following 1X PBS wash for 5 min, sections were mounted in ProLong Diamond Antifade Mountant with DAPI (Thermo Fisher). Fluorescent images were captured with Leica DMi8, DFC365 FX camera, and LAS X software at same exposures for each panel, as indicated in figure legends.
Quantitative reverse transcription PCR analysis and vector genome quantification
Tissues were dissected, snap frozen in liquid nitrogen, and stored at −80°C until further processing. Total RNAs were extracted from tissues using TRIzol (Invitrogen) and purified from aqueous phase using the RNeasy Mini Kit (Qiagen) after on-column DNase I digestion. One microgram of total RNA was used for cDNA synthesis using iScript RT Supermix for qRT-PCR (Bio-Rad). The mCherry expression was quantified by SYBR Green qPCR assay using 10 ng of cDNA and primers 5′-GAACGGCCACGAGTTCGAGA and 5′-CTTGGAGCCGTACATGAACTGAGG; Rpl37 expression for normalization was quantified using 5 ng of cDNA and primers 5′-CATCCTTTGGTAAGCGTCGCA and 5′-TGGCACTCCAGTTATACTTCCT; Ckm expression was quantified using 5 ng of cDNA and primers 5′-CCCAGGTCACCCCTTCATC and 5′-GCCAGAATACGAACAGACACC. For vector genome quantification in each tissue, DNA was isolated from remaining interphase and organic phase of TRIzol/chloroform (following aqueous phase removal for RNA isolation above). Vector genomes were quantified by SYBR Green qPCR assay (100 ng genomic DNA) using primers to the bovine growth hormone polyadenylation signal (bGH PAS) present in the reporter construct and normalized to the endogenous single-copy Rosa26 locus. Three biological replicates were used for each tissue. Oligonucleotide primer sequences are as reported. 7,23
RESULTS
Our initial goal was to design a highly minimized skeletal muscle-specific regulatory cassette for use in a clinically relevant CRISPR inhibition (CRISPRi) approach to facioscapulohumeral muscular dystrophy (FSHD). 7 To efficiently accommodate all therapeutic CRISPRi components (regulatory cassette, dCas protein fused to effector domain, nuclear localization signals, and sgRNA expression cassette) within the optimal size constraints (∼4.4 kb between the ITRs) of a single AAV vector required a regulatory cassette smaller than those currently in use. Building on key work from the Hauschka laboratory 8,9 and taking into account that cardiac muscle is not involved in FSHD, 24 –27 we designed a minimized skeletal muscle regulatory cassette to drive the expression of larger therapeutic components.
Over the years, many regulatory cassettes optimized for expression in striated muscles have been engineered 8,9 ; these have been widely disseminated and are being actively used in both academic and clinical settings. However, inconsistent nomenclature among users is a serious issue, with researchers often referring to different cassettes by the same name. For example, the “tMCK” cassette in Addgene plasmid #105556 uses three copies of the wild-type Ckm enhancer, while “tMCK” in Addgene plasmid #149410 uses three copies of a modified Ckm enhancer. Additionally, the commonly used 438-bp “CK8e” cassette 10 –17 contains only one modified Ckm enhancer, but this is sometimes inaccurately referred to in the literature as “CK8.” With these complications in mind, we started with the ∼800-bp CK8 cassette, as originally described: three tandem copies of a modified Ckm enhancer upstream of a modified Ckm promoter 8,9 (Fig. 1). We removed additional space between elements and deleted the CArG and AP2 sites (Fig. 1). The latter appears to bind a repressive factor, as mutations in the AP2 site increased activity in cultured skeletal and cardiac myocytes. 20 The CArG site is indispensable to Ckm expression in cardiac muscle 20,21 ; however, its importance in skeletal muscle is not so clear. While mutations in the CArG site reduced activity in cultured skeletal myocytes, 20 they had no effect in transgenic mouse skeletal muscle. 21 Thus, it seemed worthwhile to test removal of this sequence for the purpose of minimizing the cassette and selectively lowering expression in the heart. We also mutated the Left E-box sequence to create an additional Right E-box, a change that was reported to increase activity in skeletal myocytes, 18,19 and reduced the skeletal and cardiac muscle-specific proximal promoter to the 80-bp basal non-muscle-specific sequence. 20,21 In our previous study, this cassette was active in all skeletal muscles tested, except for soleus, EDL, and diaphragm; as expected, it was virtually inactive in the heart and nonmuscle tissues. 7 Thus, we named this regulatory cassette No Heart (NH).
Although NH is active in most skeletal muscles, the critical target tissue for FSHD and a host of other skeletal myopathies, it displays extremely low activity in soleus and diaphragm, two muscles containing a high proportion of slow-twitch fibers, and in EDL, a fast-twitch muscle. 7 This suggested that one or more of the NH modifications may have lowered overall activity in skeletal muscles as compared with the original CK8. In an effort to increase activity in all skeletal muscles while maintaining minimal cardiac expression, we redesigned the NH cassette by substituting the original Left E-box from the Ckm enhancer in place of the additional Right E-box modification 9 (Fig. 1). Following the analysis described below, this new cassette was named Have a Little Heart (HLH) due to the low cardiac activity that accompanied the restoration of moderate-to-high activity in most skeletal muscles.
In vivo activities of NH and HLH were compared directly with CK8. Due to practical constraints, male mice were used for NH and HLH, and female mice were used for CK8. While equal numbers of male and female mice would have been ideal for comparing all regulatory cassettes, expression of the endogenous Ckm gene has not been reported to be affected by sex. Genes in human muscle that are differentially expressed between the sexes fall mainly into pathways that regulate muscle mass and do not include Ckm. 28 Indeed, a recent transcriptomic study showed that Ckm was expressed equally between male and female mice. 29 Nonetheless, we examined endogenous Ckm levels in the same tissues that were harvested for reporter expression, and found no significant differences in our male vs. female mice (Supplementary Fig. S1). While we cannot exclude the possibility that Ckm-based cassettes, which are modified from the endogenous sequence, may express differently in male and female mice, these data strongly suggest that the relative activities of these cassettes are not due to differences in sex.
Each cassette driving the mCherry reporter was delivered systemically to wild-type mice using AAV9(Y731F), an AAV9 variant that is robustly delivered to body-wide striated muscles. 30,31 Viral particles were delivered by ROI at 2.6×1014 vector genomes kg body weight. This dose was chosen as the lowest that produces uniform transduction within muscles (lower doses produced mosaic reporter expression). Expression of the mCherry reporter was quantified as epifluorescence in abdominal and pectoral muscles in live mice, monitored weekly over 11 weeks. Epifluorescence was similarly analyzed in noninjected wild-type mice to control for background and autofluorescence, which was negligible (Supplementary Fig. S2). As anticipated, NH showed lower skeletal muscle activity than CK8 at all time points (Fig. 2). In contrast, the profile of HLH activity was very similar to CK8 (Fig. 2). As with CK8, HLH rapidly achieved high activity. Compared with the peak total flux of NH (observed at 8 weeks), total flux of HLH approached these levels after 2 weeks and was 5× higher by 3 weeks (Fig. 2A). The total flux is the epifluorescence signal (radiance) summed over the area of detection, and may vary between animals; however, even by the maximum radiance measurement (the highest radiance for a pixel in the field), HLH was comparable to CK8 and outperformed NH (Fig. 2B). Compared with the peak maximum radiance of NH, maximum radiance of HLH approached these levels after 1 week and was 6× higher by 3 weeks (Fig. 2B).
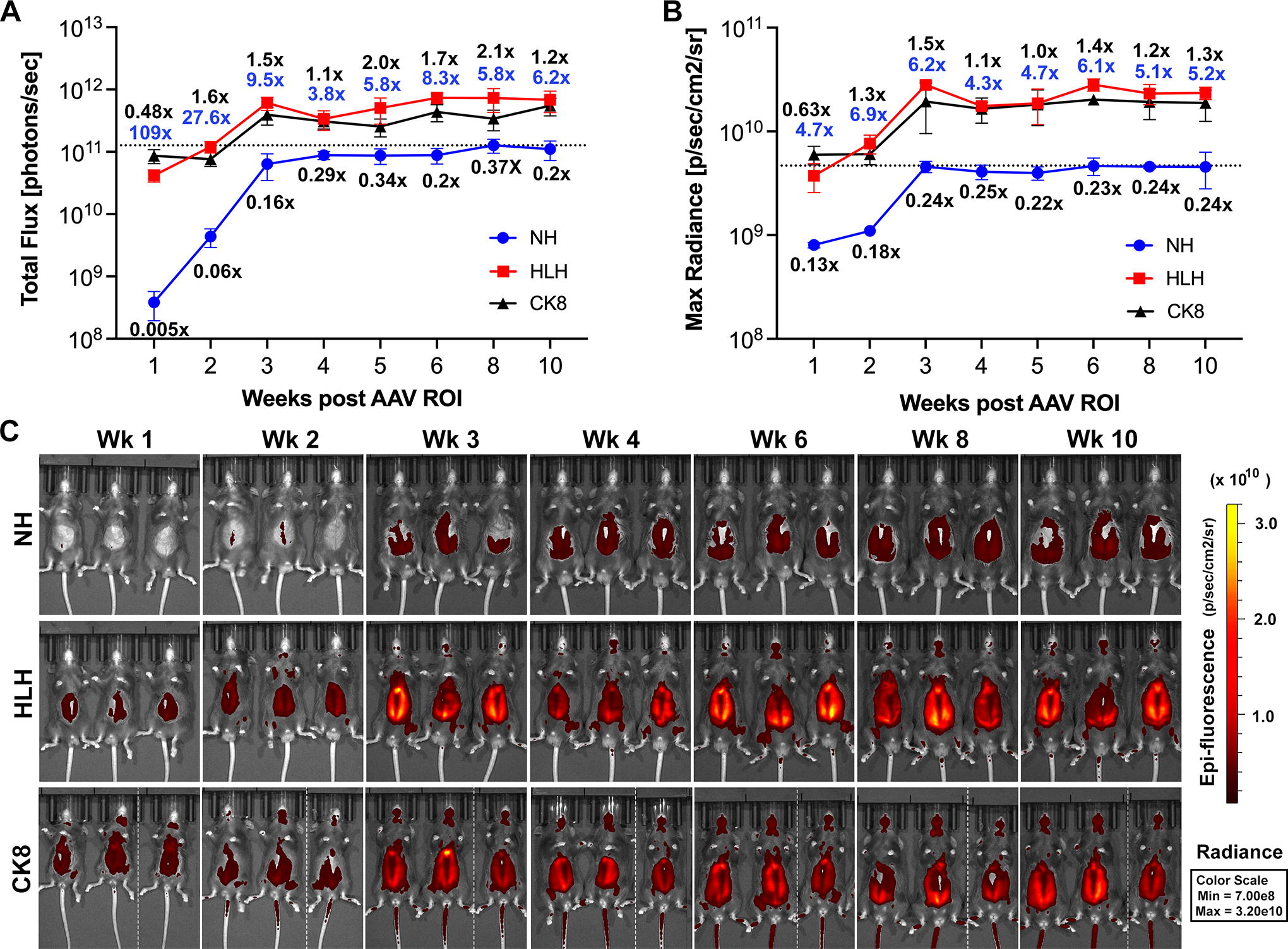
The HLH cassette displays similar activity to CK8 and higher activity than No Heart NH in skeletal muscle over time. The mCherry reporter under control of the NH, HLH, or CK8 regulatory cassette was delivered in AAV9(Y731F) at 2.6 × 1014 vg/kg by retro-orbital injection to wild-type mice (n = 3 per virus). The epifluorescent signal in abdominal and pectoral muscles was measured as
To compare cassette activities in other muscles and tissues, mice were sacrificed at 12 weeks postinjection for in vitro epifluorescence quantification and fluorescence imaging of whole tissues and sections. Quantitative PCR (qPCR) for vector genomes present across tissues showed that AAV9(Y731F) strongly transduced liver, with moderate transduction of heart, kidney, and skeletal muscles (Supplementary Fig. S3). 31
Mice injected with NH-mCherry in AAV9(Y731F) showed expression in tibialis anterior (TA), gastrocnemius, and quadriceps, as well as pectoral, abdominal, back, and facial muscles (Figs. 3 –5; Supplementary Fig. S4). Expression was virtually undetectable in soleus, EDL, diaphragm, and heart (Figs. 3 –5), even at a higher exposure (Supplementary Fig. S4), and in nonmuscle tissues (Supplementary Fig. S5). This recapitulates the expression pattern we observed with this cassette delivered in AAV9 virions, 7 confirming that very low activity in certain skeletal muscles and virtually undetectable activity in the heart is intrinsic to NH and not due to differences in vector transducibility of the muscles.
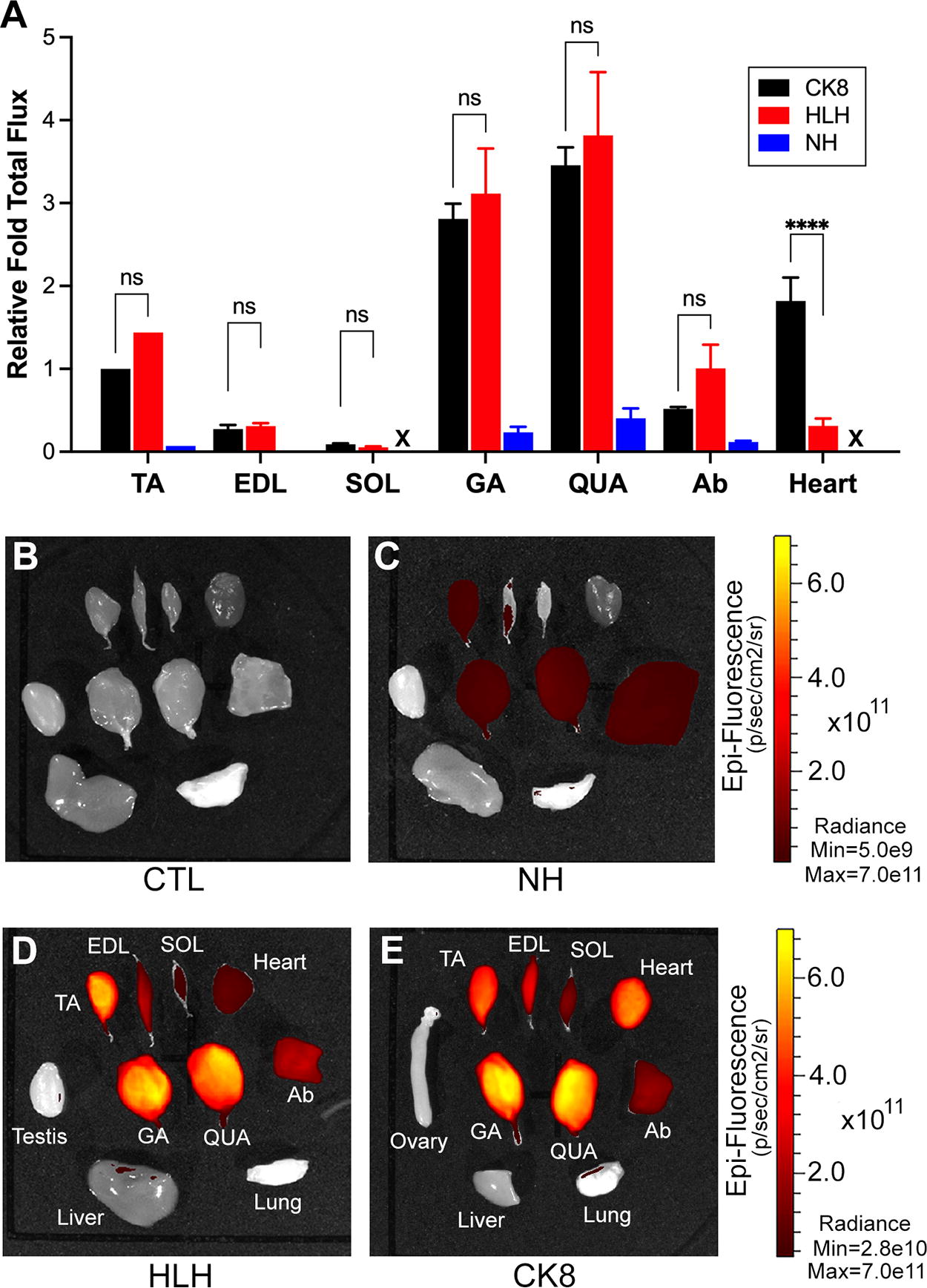
The HLH cassette displays similar activity to CK8 in skeletal muscles and much lower activity in the heart, whereas NH displays lower activity in all striated muscles. All mice from Figure 2 were sacrificed at 12 weeks postinjection and individual tissues were dissected.
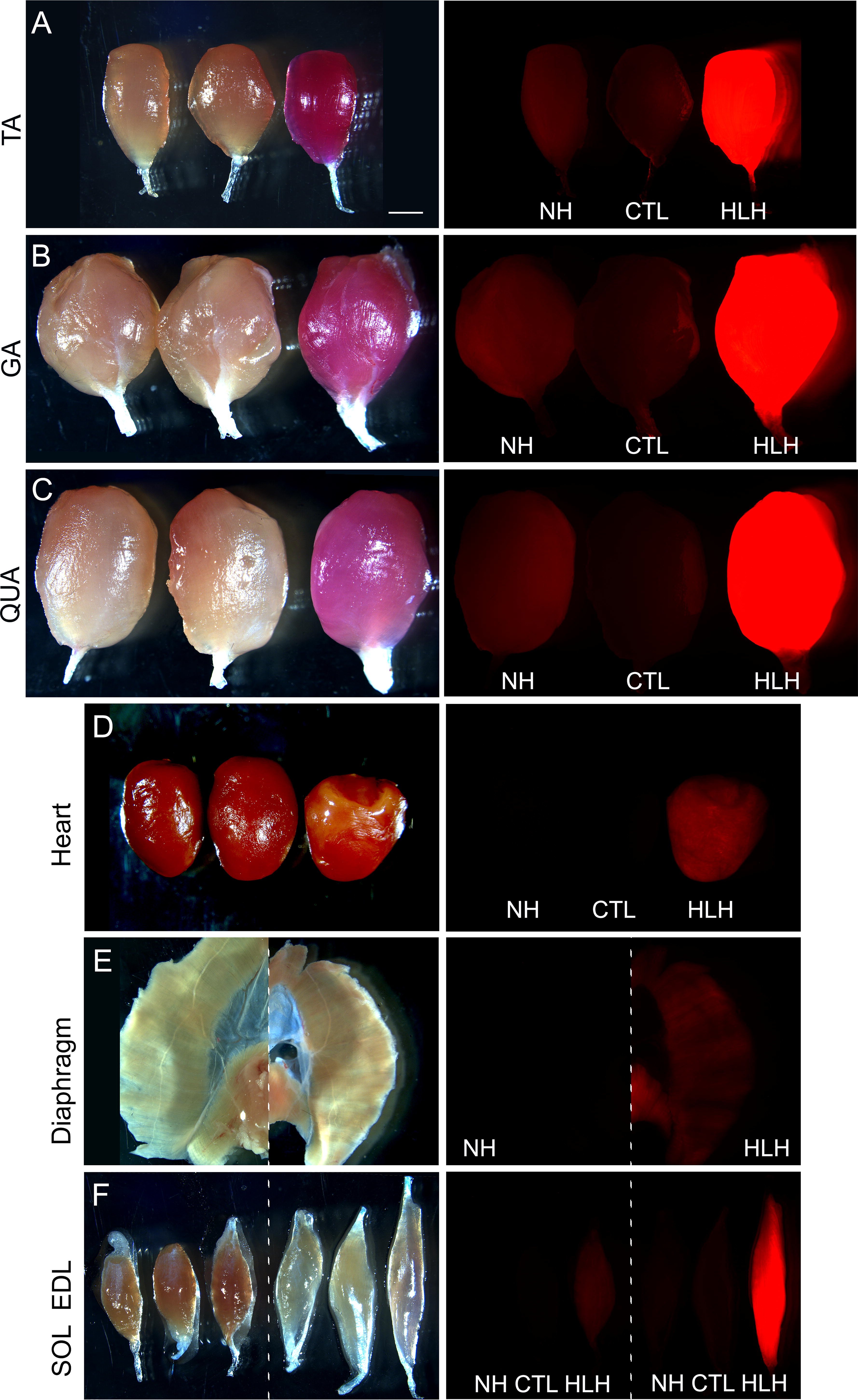
The HLH cassette displays higher activity than NH in all striated muscles. Fluorescent signals in the isolated muscles from Figure 3 were visualized with a 0.2-s exposure time.

The HLH cassette displays similar activity to CK8 in skeletal muscles and much lower activity in the heart, while NH displays lower activity in all striated muscles. Fluorescent signals in cryosections of striated muscles from Figure 3 were visualized with
Restoration of the Left E-box in HLH (Fig. 1) dramatically increased reporter expression in all fast-twitch muscles compared with NH, and most importantly increased expression from near background levels in soleus, EDL, and diaphragm (Figs. 3 –5; Supplementary Fig. S6), while maintaining undetectable expression in nonmuscle tissues (Supplementary Fig. S7). The total flux measurements showed ∼20×, 37×, 12×, and 9× higher mCherry expression with HLH versus NH in TA, EDL, gastrocnemius, and quadriceps muscles (Fig. 3a, blue numbers); Table 1 this level of expression was high enough to turn muscles noticeably pink even under visible light. Although reporter expression in the heart was also increased with HLH versus NH, it was still considerably lower than with CK8 (Figs. 3 –5). Importantly, mice transduced with CK8 and HLH had very similar levels of vector genomes in the heart (Supplementary Table S1; Fig. 6); thus, the strikingly low cardiac expression seen with HLH is not due to lower vector transduction. Except for the diaphragm, where CK8 is more active than HLH, the pattern of relative activity across skeletal muscles is very similar between the two cassettes. To eliminate the influence of differences in total areas on signal measurements between different tissues, we used max radiance data from the in vitro optical imaging analysis to compare all cassette activities in skeletal muscles and heart, which confirmed the results obtained from the total flux data (Supplementary Fig. S8).
Relative Activities of Regulatory Cassettes
Data from Figure 3A are presented as relative activities of the indicated cassettes in each tissue (e.g., in the TA, HLH was 1.4X as active as CK8 and 20X as active as NH, whereas NH was only 7% as active as CK8). For additional details, see Figure 3A.
HLH, Have-a-Little-Heart; NH, No Heart; TA, tibialis anterior; EDL, extensor digitorum longus; SOL, soleus; GA, gastrocnemius; QUA, quadriceps; Ab, abdominal muscles; nd, signal not detected.

Reporter transcript levels verify that the HLH cassette displays similar activity to CK8 in skeletal muscles and much lower activity in the heart, while NH displays lower activity in all striated muscles.
For a more quantitative measurement of relative cassette activities, mCherry expression was assessed by qRT-PCR (Fig. 6). Interestingly, reporter levels in individual mice did not correlate well with levels of vector genomes (i.e., for every given tissue type, the highest-transduced sample did not have the highest reporter expression) (Supplementary Table S1). This suggests that past a certain threshold, increased vector transduction does not necessarily lead to increased cargo expression. Nonetheless, taking vector copy number into account, the activity of HLH was higher than CK8 in quadriceps and abdominal muscles, and lower in TA, EDL, soleus, gastrocnemius, and diaphragm (Fig. 6).
We conclude that HLH displays moderate-to-high activity in most skeletal muscles, comparable to that of CK8, while significantly reducing activity in the heart and maintaining undetectable expression in nonmuscle tissues. Thus, HLH may prove particularly useful for gene therapy of FSHD and similar indications.
DISCUSSION
The size constraint for efficient transgene packaging into AAV vectors has spurred the engineering of minimized gene regulatory cassettes that can drive the expression of large therapeutic cargos in striated muscle. In this study, we describe two novel variations of the Ckm-based regulatory cassettes whose iterations have been extensively used in gene therapy approaches for muscular dystrophies. The smallest of these to date is the CK8e cassette now widely used in clinical settings, which contains only a single copy of a modified Ckm enhancer, plus a modified Ckm proximal promoter that also provides striated muscle-specific expression. In contrast, our NH and HLH cassettes contain three copies of a modified Ckm enhancer, as with the tMCK and CK8 cassettes, while still being slightly smaller than CK8e. As with CK8e, due to their small size, both NH and HLH can be used for diverse strategies, including gene replacement; exon skipping; polycistronic transgene expression; and CRISPR-based gene editing, activation, or inhibition approaches. Allowing for increased transgene capacity downstream of tissue-specific gene regulation is not trivial, since clinically relevant gene therapies using AAV require therapeutic cassettes contained within single vectors. 32 Eliminating the need for two or more vectors is critical for increasing the efficiency of delivery, lowering the high cost of therapy, and reducing the immunotoxicity associated with high vector doses. These concerns have been greatly mitigated by newly engineered myotropic AAV vectors, which provide increased delivery to body-wide muscles and detargeting of the liver. 33 –35 However, the use of highly minimized regulatory cassettes is still necessary for many CRISPR-based approaches, which often use multiple vectors to accommodate all therapeutic components, and for gene replacement approaches to DMD, in which the ability to increase cargo size by even a few hundred bp can be critical to efficacy. While some editing systems require so much space that dual-vector delivery is unavoidable, highly minimized regulatory cassettes such as CK8e and the two reported in this study enable a single-vector platform for many therapeutic approaches. At 378 bp, HLH and NH are by far the smallest reported regulatory cassettes containing three copies of a modified Ckm enhancer; this multimerization greatly increases cassette activity 8,9 and likely enhances the potency of the modifications. As an example, they can be accommodated into all-in-one therapeutic cassettes containing dSaCas9 fused to a minimized effector domain and all sgRNA components within the packaging limit for AAV vectors (4.4 kb of available space between ITRs). When combined with the engineered mini-Cas9 proteins or recently described smaller Cas9 orthologs, 36 –39 which could allow for expression of multiple sgRNAs or larger effector domain fusions, the potential therapeutic applications are even greater.
Our NH cassette is inactive in cardiac muscle and active in skeletal muscles with the exception of diaphragm, EDL, and soleus, tissues which can be severely affected in many muscle disorders. Thus, its use would likely be limited to cases where any expression in the heart might be detrimental, while moderate expression in some skeletal muscles would provide some benefit. If so, NH may be useful in mild cases of skeletal myopathies where cardiac muscle is unaffected (e.g., FSHD, XLMTM, central core myopathy, inclusion body myositis, most cases of nemaline myopathy, distal myopathy, centronuclear myopathy, oculopharyngeal muscular dystrophy, and certain subtypes of LGMDs where patients show no cardiac involvement). Examples of the latter include LGMD2A/R1 (calpainopathy), one of the most common LGMD subtypes, and most dysferlinopathies. Of these skeletal myopathies, gene therapy treatments for FSHD, XLMTM, and several LGMDs are under active development. 7,40 –43
In contrast to NH, HLH provides moderate-to-high expression in most skeletal muscles, comparable to that of the originally described ∼800-bp CK8 cassette, in conjunction with low expression in the heart. This makes it potentially well suited to the treatment of disorders that affect all striated muscles, but where high expression in cardiac muscle could be detrimental. These include DMD and several subtypes of LGMD and Emery-Dreifuss muscular dystrophy (EDMD). For these diseases, effective treatment requires some therapeutic expression in cardiac muscle, but the results from clinical trials suggest that lowering expression levels in the heart may prevent myocarditis from occurring. 6 Since AAV capsids that are highly tropic for skeletal muscle are also highly tropic for cardiac muscle, the best way to selectively reduce transgene expression in the heart is through transcriptional regulation (or cardiac-specific miRNA detargeting). Thus, our HLH cassette may provide beneficial regulation for therapeutic transgenes in these indications. In addition, since HLH is moderately to highly active in most skeletal muscles, it may be applicable to therapies in which high transgene expression in certain skeletal muscles is desirable and low cardiac expression is well tolerated. In FSHD, for example, the heart is unaffected, but the soleus is often severely affected and a desirable target muscle for therapeutic delivery. Thus, the HLH cassette may be a better choice than NH for this disease. It is worth emphasizing that the utility of either cassette depends on the goals and limitations of the treatment. If, for example, high cardiac expression is well tolerated and higher expression than HLH can provide is required in certain skeletal muscles (such as diaphragm or soleus), the use of CK8 or CK8e may be a better choice. To the best of our knowledge, no regulatory cassette displays very high activity in slow-twitch muscles, a predominant fiber type in humans as opposed to mice, or in the diaphragm; the latter remains a particular limitation for gene therapies of DMD, since patients often die of respiratory failure. However, depending on the indication and therapy, therapeutic gene expression may be achieved with only modest levels; thus, even a low-activity cassette may be useful in some contexts.
CONCLUSIONS
Our minimized muscle regulatory cassettes provide increased space for transgene cargo combined with either no (NH) or low (HLH) activity in the heart. The latter aspect is of particular therapeutic relevance, as too much cardiac expression is proving clinically problematic. We propose that these new cassettes may be useful for increasing the safety of gene therapy approaches for FSHD and a number of similar myopathies, providing low expression in certain skeletal muscles is acceptable.
Footnotes
ACKNOWLEDGMENT
The authors thank Monique Ramirez and Maryam Farooqi, PharmD for technical assistance.
AUTHORS’ CONTRIBUTIONS
C.L.H., T.I.J., and P.L.J. designed the study. C.L.H. designed the regulatory cassettes and wrote the article. T.I.J. performed all experiments and analyzed data. P.L.J. secured funding and advised on the execution and writing. All authors edited and approved the final version of the article.
AUTHOR DISCLOSURE
C.L.H., T.I.J., and P.L.J. are cofounders of Renogenyx, Inc., a company focused on bringing FSHD therapeutics to the clinic, and inventors on four U.S. patent applications pertaining to the use of CRISPR inhibition for FSHD (No. 63/527,698, PCT/US2021/025940, PCT/US2017/052919) or muscle-specific regulatory cassettes for NMDs (PCT/US2023/070498).
FUNDING INFORMATION
This study was funded by the Mick Hitchcock, PhD, Endowed Chair in Medical Biochemistry at UNR, an award from
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.