
Abstract
Mucopolysaccharidosis type IVA (MPS IVA) is an autosomal congenital metabolic lysosomal disease caused by a deficiency of the N-acetyl-galactosamine-6-sulfate sulfatase (GALNS) gene, leading to severe skeletal dysplasia. The available therapeutics for patients with MPS IVA, enzyme replacement therapy and hematopoietic stem cell transplantation, revealed limitations in the impact of skeletal lesions. Our previous study, a significant leap forward in MPS IVA research, showed that liver-targeted adeno-associated virus (AAV) gene transfer of human GALNS (hGALNS) restored GALNS enzymatic activity in blood and multiple tissues and partially improved the aberrant accumulation of storage materials. This promising approach was further validated in our current study, where we delivered AAV8 vectors expressing hGALNS, under the control of a liver-specific or ubiquitous promoter, into MPS IVA murine disease models. The results were highly encouraging, with both AAV8 vectors leading to supraphysiological enzymatic activity in plasma and improved cytoplasmic vacuolization of chondrocytes in bone lesions of MPS IVA mice. Notably, the ubiquitous promoter constructs, a potential game-changer, resulted in significantly greater enzyme activity levels in bone and improved pathological findings of cartilage lesions in these mice than in a liver-specific one during the 12-week monitoring period, reinforcing the positive outcomes of our research in MPS IVA treatment.
INTRODUCTION
Mucopolysaccharidosis type IVA (MPS IVA, Morquio syndrome type A) is a lysosomal disease caused by a deficiency of a lysosomal hydrolase, N-acetyl-galactosamine-6-sulfate sulfatase (GALNS). 1 –4 Such deficiency results in the accumulation of glycosaminoglycans (GAGs), including chondroitin-6-sulfate and keratan sulfate (KS), in systemic organs, especially bone and cartilage, resulting in severe skeletal dysplasia. 5 –8 The main skeletal symptoms of MPS IVA include pectus carinatum, kyphoscoliosis, genu valgum, joint laxity, and spinal cord compression. Nonskeletal abnormalities such as respiratory failure, audiology complications, and cardiovascular pathologies are also critical features recognized in patients with MPS IVA. Tracheal obstruction is life-threatening, causing a high mortality and morbidity rate. 9 –13
Enzyme replacement therapy is currently the only approved pharmacological treatment option for patients with MPS IVA. 14 –17 However, the previous studies have shown that even with the weekly infusion of recombinant human GALNS (hGALNS) enzyme initiated under 2 years of age, there was no significant reduction in plasma KS levels or accumulation of storage materials in cartilage lesions, 18 –21 and accumulation of storage materials, in cartilage lesions, 22,23 resulting in no improvement in bone growth. 24
This lack of improvement in bone growth could be attributed to the short half-life of the infused hGALNS enzyme, which leads to insufficient delivery of the enzyme into avascular cartilage tissue. 16,25 –27
Hematopoietic stem cell transplantation (HSCT), another therapeutic option, provides the donor’s level of GALNS activity at the full engraftment, showing more excellent activities of daily living scores in transplanted patients compared to similar-aged, untreated patients. 28 –30 However, HSCT has only shown limited improvement in bone growth in transplanted patients with MPS IVA, and there have been no case reports of patients under 2 years old who have received HSCT. 28,30 –32 These limitations underscore the need for alternative treatment approaches.
Gene therapy, a promising approach to treating rare genetic diseases, is currently being studied in numerous clinical trials. In particular, gene transfer using viral vectors, including adeno-associated virus (AAV), has shown effectiveness in animal models. 14,25,33
Clinical trials utilizing AAV-mediated gene therapy are ongoing in various types of MPS, including MPS I, II, IIIA, IIIB, and VI (clinicaltrials.gov). 34,35
Adeno-associated virus has several biological features that make it an attractive vector, including low immunogenicity, nonintegration or minimal integration into chromosomes, and long-term expression of transgene products. These features offer significant advantages over other viral vectors. 36 –41
Adeno-associated virus capsid modification and selection of a cell-specific promoter allow the effective expression of transgene products in specific tissues. However, targeting bone and cartilage lesions remains a significant challenge. Although a liver-specific promoter, thyroxin-binding globulin (TBG), packaged into AAV8, improved bone length in feline and rodent models of MPS VI, an effective promoter construct has yet to be defined for skeletal lesions in MPS IVA. 42,43 This underscores the need for further research in this area.
Our previous research indicated that delivering the hGALNS gene to the liver using AAV gene transfer at a dose of 5 × 1013 genome copies (GC)/kg led to higher than normal enzyme levels in the blood and provided partial improvements in bone and cardiovascular issues in both male and female MPS IVA mouse models. 44,45 We observed that the immune response (raising anti-GALNS antibody) differed between male and female mice, with female mice showing a more robust immune response to the treatment. This could lower the GALNS enzyme activity and reduce effectiveness in treating bone and heart problems compared to male mice. 44 However, we did not explore the impact of varying doses in these previous studies, and we only studied the MPS IVA knockout mouse model without considering other types of mouse models. We could not quantitatively analyze bone structure accurately, as the bone changes were subtle and not easily identified using previous micro-computed tomography (CT) technology. 46 Recent advancements in micro-CT technology have made it feasible to detect even minor changes in bone structure. 47 –52 In this study, we aimed to assess the effects and immune responses to AAV8 expressing hGALNS under the control of a liver-specific promoter at a higher dose (2 × 1014 GC/kg), using two MPS IVA mouse models (knockout and immune-tolerant to hGALNS). We compared the outcomes and immune responses between liver-specific and ubiquitous promoters using biomarkers and advanced micro-CT.
MATERIALS AND METHODS
Expression cassette design and AAV vector production
AAV8 vectors were used to express codon-optimized hGALNS with or without an aspartic acid octapeptide (D8) N-terminal tag. These vectors were placed under the control of the liver-specific promoter TBG (AAV8.TBG.hGALNS or AAV8.TBG.D8-hGALNS) and a ubiquitous cytomegalovirus early enhancer/chicken actin (CAG) promoter (AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS). As described previously, the Vector Core of Regenxbio Inc. (Rockville, MD) produced them. 45 Each vector construction is shown in Supplementary Fig. S2. The expression cassette plasmids were transfected into the human liver cell line, Huh-7 (Millipore Sigma; Cat. 01042712, St. Louis, MO), and GALNS activity levels were determined in both cell lysates and supernatants 48 h post-transfection (Supplementary Fig. S3).
Mouse experimentation
All mouse care and procedures followed NIH guidelines and were approved by the Nemours Children’s Health Institutional Animal Care and Use Committee at Nemours Children’s Health (IACUC; RSP19-12482-001). Our group previously created two MPS IVA mouse models, GALNS knockout (KO, Galns−/−) mice and MPS IVA mice tolerant to hGALNS protein (MTOL, Galnstm[hC79S.mC76S)slu]) in a C57BL/6 background. 53
Both mouse models had no detectable GALNS enzymatic activity in blood and tissues and displayed excessive accumulation of storage materials primarily within reticuloendothelial Kupffer cells, muscle, heart valves, and chondrocytes, including articular cartilage and growth plates. 44,45
The genotype of each mouse was confirmed by tail biopsies using PCR on day 14. Four-week-old homozygous MPS IVA mice, both male and female, were treated via lateral tail vein at a dose of 5 × 1013 GC/kg AAV8 with the CAG promoter or 2 × 1014 GC/kg for AAV8 with the TBG promoter. Another cohort of MPS IVA mice and unaffected C57BL/6 littermates (wild-type) was administered phosphate-buffered saline (PBS). The total dose volume administration was approximately 100 μL per mouse. Then, 100 μL of whole blood was taken in EDTA tubes (Becton Dickinson, Franklin Lakes, NJ) from all study groups every other week. Blood was centrifuged at 8,000 rpm for 10 min, and separated plasma was stored at −20°C before GALNS enzyme and GAG assays. At 16 weeks, mice were euthanized in a CO2 chamber and perfused with 20 mL of 0.9% saline. Livers, kidneys, lungs, spleens, hearts, and knee joints were collected and stored at −80°C prior to GALNS enzyme and GAG assays. In addition, for histopathology, knee joint samples were collected and stored in 10% neutral buffered formalin.
Adeno-associated virus genome copy quantification
The number of AAV GC in the liver was measured using the digital PCR method following the protocol previously described. 45,54
GALNS enzyme activity assay
GALNS activities in plasma and tissues were determined as described previously. 45,54 Frozen tissues were homogenized in a homogenization buffer of 25 mM Tris-HCl (pH 7.2) and 1 mM phenylmethylsulfonyl fluoride using a Polytron homogenizer. Plasma or tissue lysates and 22 mM 4-methylumbelliferyl-β-galactopyranoside-6-sulfate (a fluorogenic substrate for GALNS, Research Products International, Mount Prospect, IL) in 0.1 M NaCl/0.1 M sodium acetate (pH 4.3) were incubated in a water bath at 37°C for 16 h. Then, 10 mg/mL β-galactosidase from Aspergillus oryzae (Sigma-Aldrich, St. Louis, MO) in 0.1 M NaCl/0.1 M sodium acetate (pH 4.3) was added to the reaction sample, with additional incubation at 37°C for 2 h. The sample was transferred to the stop solution (1 M glycine, NaOH [pH 10.5]), and the plate was read at excitation 366 nm and emission 450 nm in a PerkinElmer Victor X4 plate reader (PerkinElmer, Waltham, MA). The GALNS enzymatic activity was expressed as nanomoles of 4-methylumbelliferone released per hour per milliliter of plasma or milligram of protein. Protein concentration was determined by a bicinchoninic acid protein assay kit (Thermo Fisher Scientific, Waltham, MA).
GAG extraction from tissues
GAG extraction from mouse livers was performed using a previous method 55 with minor modifications. The dissected mouse liver was frozen in liquid nitrogen and homogenized in cold acetone. The obtained powder was dried under a centrifuge vacuum. The defatted tissue powder was suspended in 0.5 M NaOH and incubated at 50°C for 2 h to remove GAG chains attached to its core protein. After neutralization with 1 M HCl, NaCl was added to a final concentration of 3 M. The suspension was then centrifuged to remove insoluble materials, and the pH of the supernatant was adjusted below 1.0 with 1 M HCl to precipitate nucleotides. Then, samples were centrifugated; the supernatant was neutralized with 1 M NaOH, and crude GAGs precipitated by adding two times volumes of ethanol containing 1.3% potassium acetate. After centrifugation, the residue was resuspended in distilled water.
GAG assay
Monosulfated KS levels in blood and liver were measured by liquid chromatography-tandem mass spectrometry after enzymatic digestion of KS polymers, as described previously. 56,57 Briefly, 50 mM Tris-HCl (pH 7.0) and samples were added to 96-well Omega 10K molecular weight cutoff filter plates (Pall Corporation, Port Washington, NY) on a 96-well receiver plate. Samples were centrifuged for 15 min at 2,500 ×g. The filter plate was then transferred to a fresh receiver plate, and a cocktail mixture of 50 mM Tris-HCl (pH 7.0) with 1 mU keratanase II (Seikagku Co., Tokyo, Japan) and 5 μg/mL chondrosine as an internal standard (IS) was added to the filter plate. Samples were incubated at 37°C in a water bath overnight. Then, the samples were centrifuged for 15 min at 2,500 ×g. The equipment consisted of a 1290 Infinity liquid chromatography coupled with a 6460 triple quad mass spectrometer (Agilent Technologies, Palo Alto, CA). Injection of 10 μL volume was injected into a Hypercarb column (2.0 mm i.d., 50 mm, 5-μm; Thermo Fisher) at 60°C to separate disaccharides. The mobile phase consisted of a binary mixture of 5 mM ammonium acetate (pH 11.0) (solvent A) and 100% acetonitrile (solvent B). The flow rate was 0.7 mL/min, with a running time of 5 min per sample, and the gradient was as follows: 0 min, 100% solvent A; 1 min, 70% solvent A; 2 min, 70% solvent A; 2.20 min, 0% solvent A; 2.60 min, 0% solvent A; 2.61 min, 100% solvent A; and 5 min, 100% solvent A. The mass spectrometer was operated with electrospray ionization in the negative ion mode (Agilent Jet Stream Technology). Specific precursor and product ions and m/z ratios were used to quantify each disaccharide, respectively (IS, 354.3→193.1; monosulfated KS, 462→97).
Bone pathological assessments
Toluidine blue staining was performed as described previously. 44,45,54,58 –62 Knee joints were collected from 16-week-old MPS IVA and wild-type (WT) mice to evaluate levels of storage granules by light microscopy. Tissues were fixed in 2% paraformaldehyde and 4% glutaraldehyde in PBS, post-fixed in osmium tetroxide, and embedded in Spurr’s resin. Then, toluidine blue-stained 0.5-µm-thick sections similar to the size of electron microscopy were examined. To evaluate chondrocyte cell size (vacuolization) in the growth plates of femur or tibia, approximately 300 chondrocytes in the proliferative area were measured in each mouse by Image J software, and results were expressed as folds-change from the wild-type group. Pathological slides from knee joints of treated and untreated MPS IVA and wild-type mice were evaluated for reduced vacuolization and improved column orientation in growth plates, as described and validated before. 63 –65 The amount of storage materials and the degree of disoriented columns were scored “No storage or very slight” was 0 (−), “slight but obvious” was 1 (+), “moderate” was 2 (++), and “marked” was 3 (+++). Each pathological slide was assessed in a double-blind manner three times. We then averaged the scores in a group of mice per section of bone (growth plate, articular disc, meniscus, and ligament). 44,45,63,64
Micro-CT analysis
Micro-CT analysis was performed as described previously 58,64,68 on the femurs of male MPS IVA mice. This analysis was conducted at a crucial time point, 12 weeks post-injection of AAV vectors, with or without a bone-targeting signal. The samples were then examined using a SkyScan 1275 system (Bruker, Billerica, MA), with a source voltage and current of 80 kV and 125 μA, respectively. The resulting three-dimensional microstructural images were reconstructed using SkyScan NRecon software. Subsequently, Bruker CTAN software was utilized to calculate trabecular bone volume (BV/TV, %), trabecular number (Tb.N, 1/mm), and bone mineral density (BMD, gHA/cm3).
Detection of plasma anti-GALNS IgG antibodies
To evaluate the immune reaction of the gene product, we checked the level of anti-hGALNS antibodies in plasma as reported previously. 44,45,58 We used the indirect ELISA method. 45,57,58,66 –68 First, 96-well microtiter plates were coated with 2 μg/mL purified rhGALNS (R&D Systems, Minneapolis, MN) and then blocked with 3% bovine serum albumin in PBS (pH 7.2). Next, we added diluted plasma (1:100) to the wells and incubated them at 37°C for 2.5 h. Afterward, we added a peroxidase-conjugated goat antimouse IgG (Thermo Fisher) at a 1:1,000 dilution, which was then incubated for 1 h at room temperature. We then added peroxidase substrate (ABTS solution, Invitrogen, Carlsbad, CA) and incubated the plates for 30 min. The reaction was stopped with the addition of 1% SDS, and the absorbance was read at an optical density of 410 nm on a Perkin Elmer Victor X4 plate reader (PerkinElmer).
Statistical analysis
All data were expressed as means and standard deviations. Multiple comparison tests were performed using one-way ANOVA, with Bonferroni’s post hoc test, in GraphPad Prism 5.0 (GraphPad, San Diego, CA). Statistically significant differences were considered at p < 0.05.
RESULTS
Human GALNS activity in blood and tissues in MPS IVA mice
Our previous findings revealed a supraphysiological level of GALNS enzyme activity, 4- to 19-fold higher than wild-type mice, using AAV8 vectors expressing hGALNS. 45 This crucial observation, under the control of the liver-specific TBG promoter, forms the basis of our current study. We have also tested immune-tolerant mice, which express an inactive form of hGALNS as a model of missense mutation since over 90% of patients with MPS IVA have the missense mutation in one of the mutant alleles. In this research, we delivered AAV8 vectors (AAV8.TBG.hGALNS, AAV8.TBG.D8-hGALNS, AAV8.CAG.hGALNS,, or AAV8.CAG.D8-hGALNS) into two types of MPS IVA mouse models [knockout (Galns−/−); KO and immune tolerant; MTOL], as previously established (Supplementary Fig. S1 and Supplementary Table S1). 53,69 The inclusion of the peptide sequence (aspartic acid octapeptide) D8 as a bone-targeting signal further enhanced the GALNS enzyme affinity for hydroxyapatite, a key component of bone. 65 KO mice were intravenously administered 2.0 × 1014 GC/kg of AAV8. TBG.hGALNS or AAV8.TBG.D8-hGALNS, or 5.0 × 1013 GCs/kg of AAV8.CAG.hGALNS or AAV8. CAG.D8-hGALNS. MTOL Mice were intravenously administered 5.0 × 1013 GC/kg of AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS at 4 weeks old. Treated and untreated MPS IVA and wild-type mice were monitored for 12 weeks after injection. Also, this study included mice of both sexes (females and males).
Plasma enzyme activities in KO and MTOL mice are shown in Fig. 1A and B. No GALNS activity was found in the plasma of untreated MPS IVA mice (data not shown). Two weeks after treatment, the plasma hGALNS activity in KO mice that received AAV8.TBG.hGALNS was ∼8 nmol/h/mL, and the mice with AAV8.TBG.D8-hGALNS had 86.4 nmol/h/mL, markedly higher than the activity in wild-type mice (0.53 nmol/h/mL). The plasma enzyme activities from the AAV8.TBG.D8-hGALNS group were about 10 times higher than those from the AAV8.TBG.hGALNS group. However, 12 weeks after treatment, there was no difference in plasma hGALNS activity between these two AAV vector groups. Similar patterns were also observed in KO mice treated with AAV8.CAG.hGALNS (8 nmol/h/mL) or AAV8.CAG.D8-hGALNS (142 nmol/h/mL) two weeks post-injection.
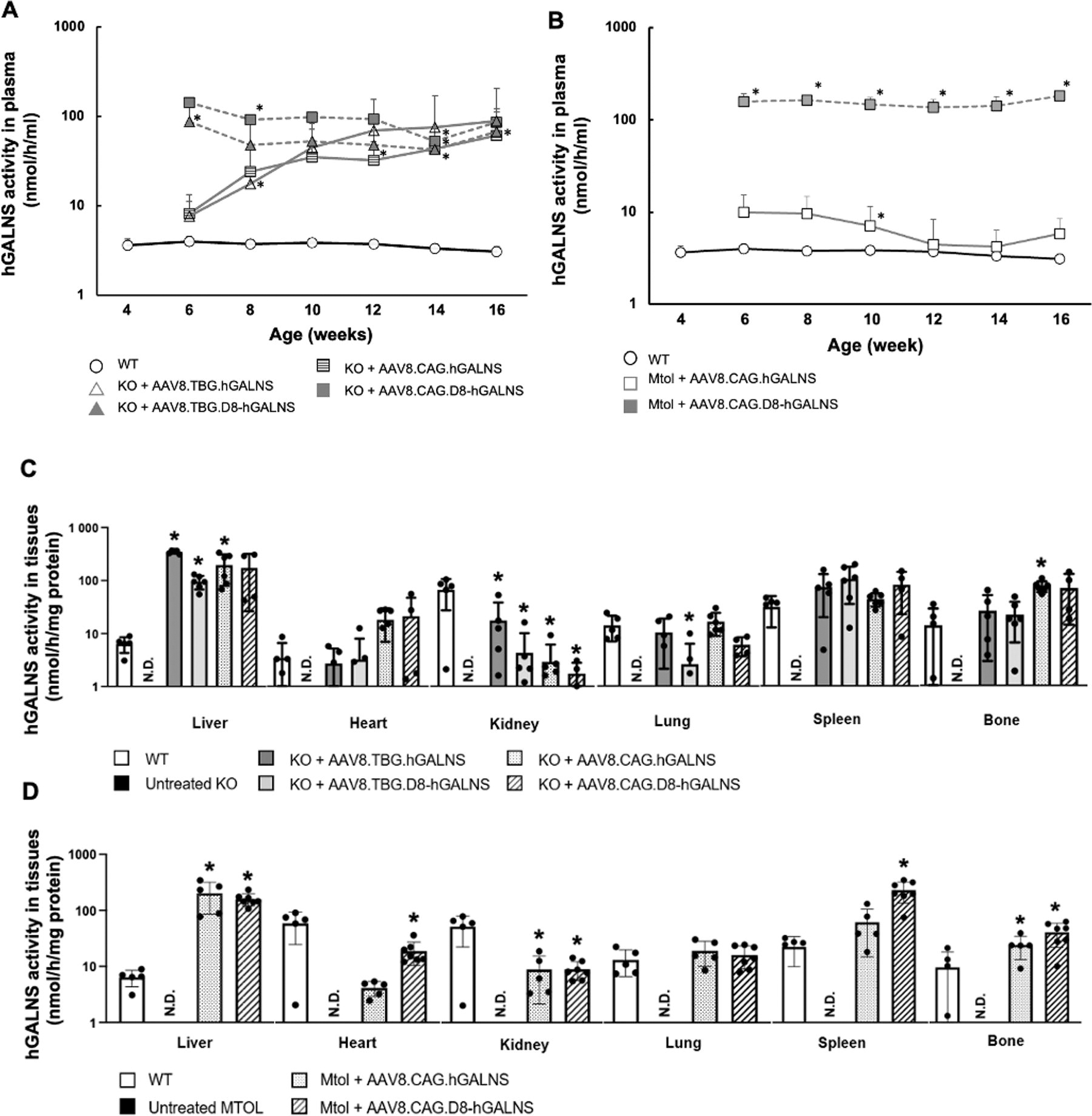
hGALNS enzyme activity in blood and tissues of MPS IVA mice treated with AAV8 vectors.
At 16 weeks (12 weeks post-administration), AAV8.TBG.hGALNS, AAV8.TBG.D8-hGALNS, AAV8.CAG.hGALNS, and AAV8.CAG.D8-hGALNS vectors induced similar levels of plasma GALNS activity, indicating that the 4-fold lower dose of the AAV8.CAG vectors achieved the same level as those with a higher dose of TBG. In MTOL mice treated with AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS, plasma hGALNS activity levels exceeded wild-type mouse levels two weeks post-injection. Enzymatic activity levels in MTOL mice treated with AAV8.CAG.D8-hGALNS (156 nmol/h/mL) were approximately 20-fold higher than those from MTOL mice treated with AAV8.CAG.hGALNS (10 nmol/h/mL) during the study period. Additionally, the enzyme activity detected between females and males showed a slight variation (data not shown), indicating a sex-dependent effect on the hGALNS activity. These results align with our previous findings. 44
Supraphysiological enzyme levels of circulating hGALNS were achieved after a single administration of the four AAV vectors in KO and MTOL mice and maintained throughout the entire study. Despite the markedly elevated levels of the hGALNS enzyme, treated mice did not show any toxic effect, change in lifespan, or tissue damage (data not shown).
The levels of hGALNS activity in the liver and other tissues 12 weeks after injecting AAV vectors are illustrated in Fig. 1C and D. We found significantly higher hGALNS activity in the livers of KO and MTOL mice treated with each AAV vector than in wild-type mice. A dose-independent increase in hGALNS activity was observed in the livers of KO mice, with lower levels seen for the D8 transgenes (Supplementary Fig. S4). This dose-dependent increase in hGALNS activity in the livers of KO mice treated with AAV8.TBG.hGALNS or AAV8.TBG.D8-hGALNS suggests that the efficacy of the treatment is influenced by the dosage of the AAV vectors. Furthermore, the hGALNS GC numbers at 16 weeks of age were higher in the livers of KO mice treated with AAV8.TBG.hGALNS or AAV8.TBG.D8-hGALNS at the dose of 2.0 × 1014 GC/kg compared with the numbers in mouse livers treated with AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS at 5.0 × 1013 GCs/kg (Table 1). A dose-dependent rise in GC number was noted in the livers of KO mice treated with AAV8.TBG.hGALNS or AAV8.TBG.D8-hGALNS (Supplementary Fig. S4).
Liver Vector Genome Copies in MPS IVA Mice Treated with AAV Vectors
Note: MPS IVA mice models were treated with 2 × 1014 GC/kg of AAV8.TBG.hGALNS, AAV8.TBG.D8-hGALNS, or 5 × 1013 of AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS, at 4 weeks of age. Liver vector genome copies were quantified using digital PCR 12 weeks’ postinjection of AAV8 vectors. Numbers represent AAV genome copies per molecule of the diploid genome (GC/diploid genome) and are expressed as mean values ± SD. n = 4–7.
AAV, adeno-associated virus; CAG, cytomegalovirus early enhancer/ chicken β-actin; D8, aspartic acid octapeptide; GC, genome copies; hGALNS, human N-acetyl-galactosamine-6-sulfate sulfatase; KO, knockout; MPS IVA, mucopolysaccharidosis type IVA; MTOL, immune tolerant; SD, standard deviation; TBG, thyroxin-binding globulin; WT, wild type.
Next, we measured hGALNS activity in tissues of MPS IVA mice to evaluate the potential cross-correction of the deficient enzyme. The activity was observed in all examined tissues, including the heart, kidney, lung, spleen, and bone (femur), in KO mice after either AAV8.TBG.hGALNS or AAV8.TBG.D8-hGALNS treatment (Fig. 1C). Whereas the AAV8.TBG.hGALNS showed higher levels of activity than AAV8.TBG.D8-hGALNS in liver, kidney, and lung, both AAV vectors successfully restored hGALNS activity in bones and hearts of treated MPS IVA mice to levels observed in wild-type mice. In addition, in AAV vectors with the CAG promoter, the hGALNS activity was observed in all examined tissues in KO and MTOL mice after either AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS treatment (Fig. 1D). Levels of hGALNS activity in bone were significantly higher in KO mice treated with 5.0 × 1013 GC/kg of AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS than treatment with 2.0 × 1014 GC/kg of AAV8.TBG.hGALNS or AAV8. TBG.D8-hGALNS (Fig. 1C and Supplementary Fig. S4A). Similar findings showed higher levels of the GALNS activity in MTOL mice treated with the ubiquitous CAG promoter construct relative to the liver-specific TBG construct. Further, the AAV8.CAG.D8-hGALNS showed an increase ∼2-fold in activity levels compared with AAV8.CAG.hGALNS (Supplementary Fig. S4B). These results suggest that AAV vectors, under the control of the CAG promoter, are preferable to those of the TBG promoter in delivering the hGALNS enzyme into the bone of MPS IVA mice.
Levels of monosulfated KS in blood and liver
Monosulfated KS is a biomarker as a storage product that accumulates in the different tissues, such as plasma and livers of KO and MTOL mice, similar to patients with MPS IVA. Plasma monosulfated KS levels in KO and MTOL mice are shown in Fig. 2A and B. Before administering AAV vectors, we observed that plasma monosulfated KS levels in untreated KO mice were significantly higher than in wild-type mice (mean 34.9 vs. 17.5 ng/mL). This difference in baseline levels is important as it indicates the disease state. Throughout this study, we found that monosulfated KS levels in the plasma of all treated groups were significantly lower than in untreated MPS IVA mice. In MTOL mice, plasma monosulfated KS levels were normalized entirely for both AAV vectors at 6 weeks of age, and these levels were maintained for at least another 10 weeks (at autopsy).
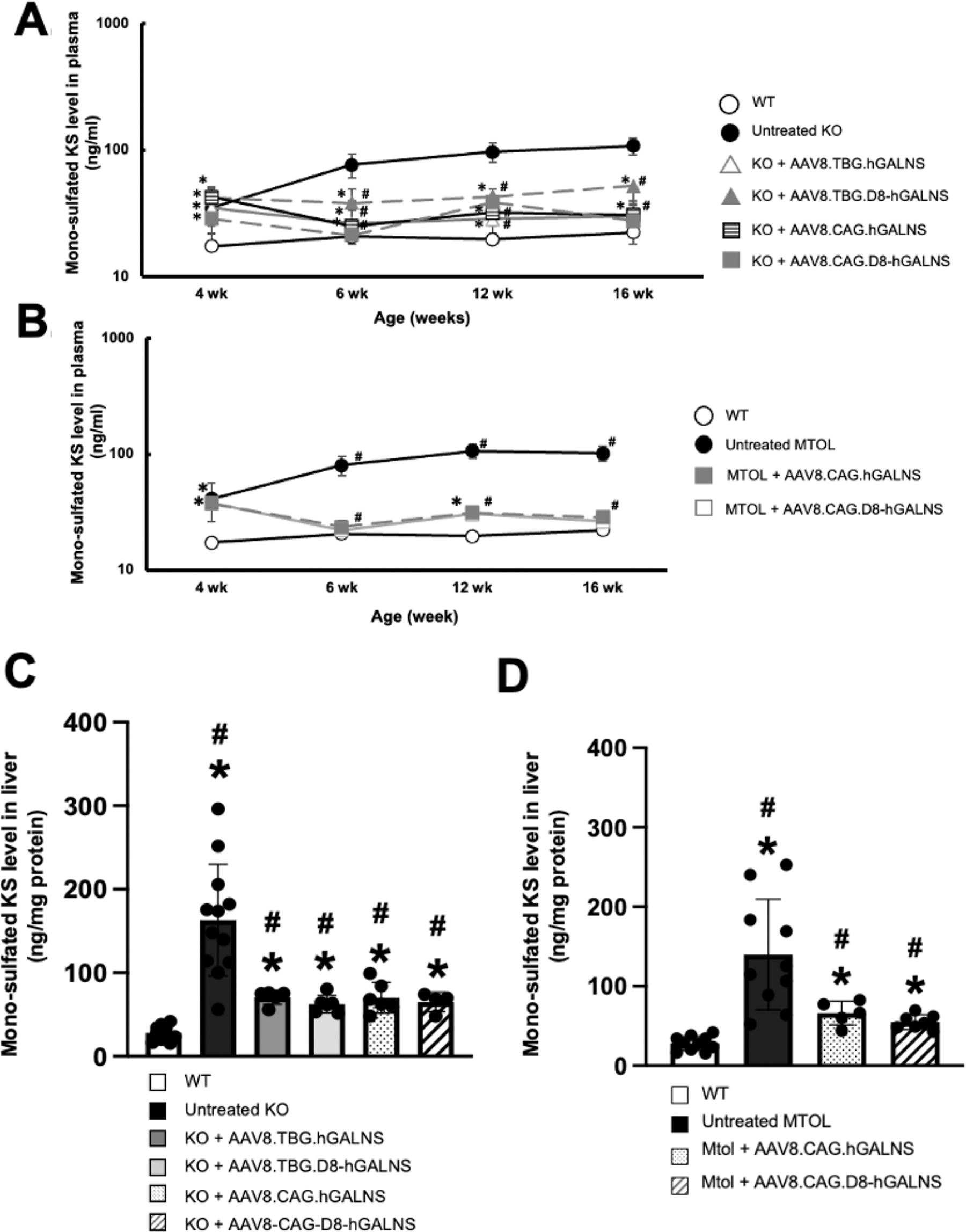
Monosulfated KS levels in plasma and liver of MPS IVA mice treated with AAV8 vectors. Monosulfated keratan sulfate (KS) levels in plasma and liver of MPS IVA mice treated with AAV8 vectors.
Our study also examined monosulfated KS levels in the livers of MPS IVA mice. Necropsy revealed excessive storage of GAGs in the livers of both KO and MTOL mice. There was a significant decrease in monosulfated KS in the livers of both KO and MTOL mice treated with each AAV vector (Fig. 2C and D). This significant decrease, although not complete normalization, marks a significant step forward in our understanding and potential treatment of MPS IVA, offering hope for the future.
Correction of bone and cartilage pathology in MPS IVA mice
Our research on the effects of each AAV vector on the femurs and tibias of MPS IVA mice has revealed promising results. Firstly, the histological analysis in untreated KO and MTOL mice at 16 weeks old exhibited excessive GAG storage as vacuolated formation in the growth plates of tibias (Fig. 3A), articular discs (Fig. 3B), and ligaments surrounding the knee joint (Supplementary Fig. S5). Disorganized column structures with swollen and vacuolated chondrocytes in growth plates were also observed in untreated MPS IVA mice (Fig. 3A and B). In KO and MTOL mice treated with each AAV8 vector, growth plates, articular cartilage, ligaments, and meniscuses of the knee joint had a partial reduction of storage material, and the column structure of chondrocytes was improved. This suggests the potential for therapeutic intervention in the future. To objectively evaluate the improvement of vacuolization in cartilage cells of the growth plate, chondrocyte cell size in the proliferative zone was quantified in KO and MTOL mice growth plate areas (tibia) (Fig. 3C and D) as described previously. 44,45,63 –65 We observed significantly reduced chondrocyte size in growth plate lesions, including femurs (Fig. 3C and D) and tibias (data not shown) of KO and MTOL mice treated with AAV8.TBG.hGALNS, AAV8. TBG.D8-hGALNS, or AAV8. CAG.hGALNS, compared to untreated KO mice (Fig. 3C) or MTOL mice (Fig. 3D). Moreover, we assessed the amelioration of vacuoles and disorganized column structures of MPS IVA mice as pathological scores, which moderately improved in KO mice treated with each AAV vector compared to untreated KO mice (Table 2). Pathological scoring of KO mice treated with AAV8 under CAG promoter showed a better trend in improving the vacuolation and column structure in articular discs and ligaments than in those receiving AAV8.TBG.hGALNS. These results suggest that the ubiquitous CAG promoter construct has more significant potential impact on the bone lesions of MPS IVA mice evaluated compared to the liver-specific TBG promoter construct, even though the dose of the vector under the control of the CAG promoter is 4-fold lower than the TBG vector.
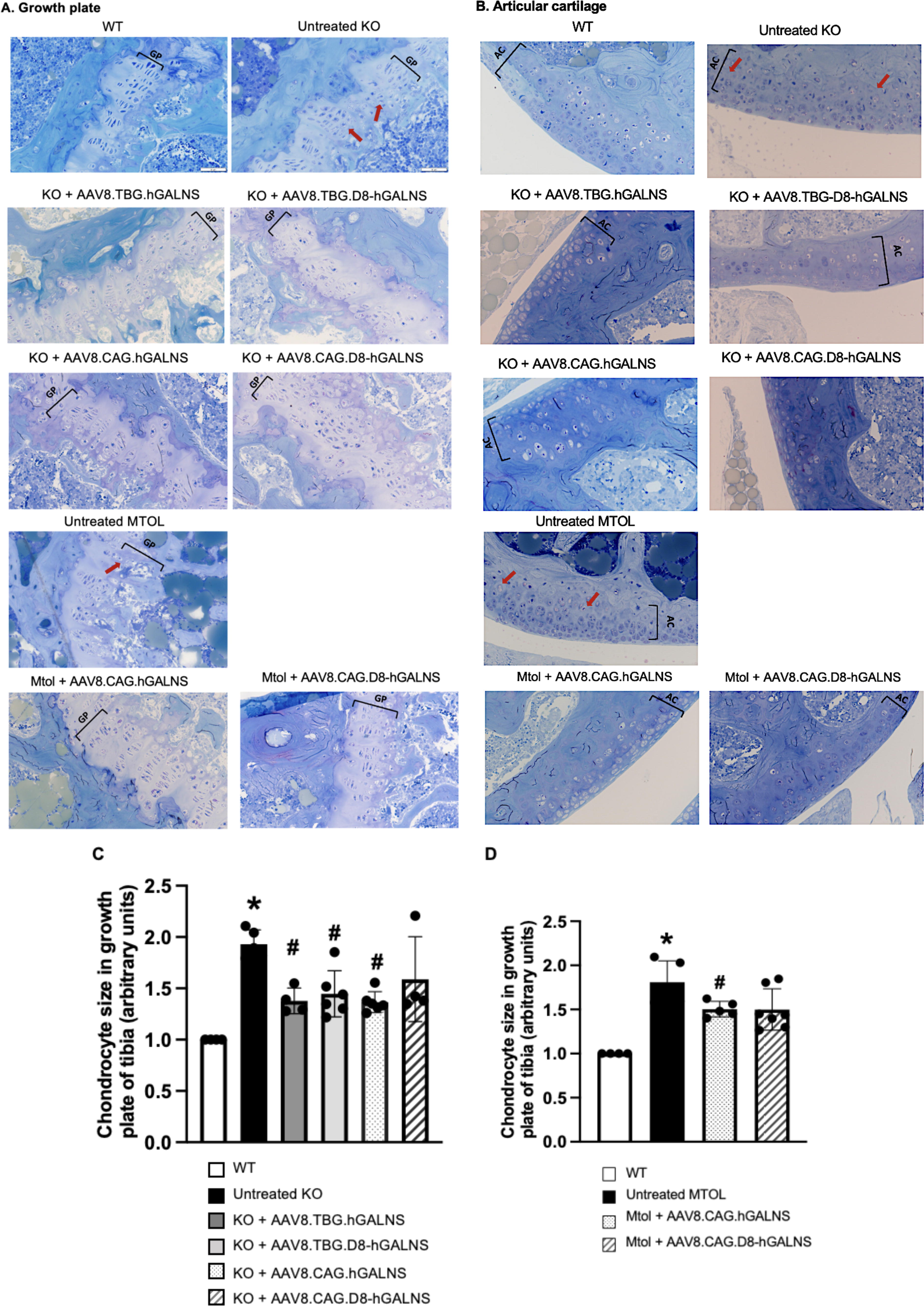
Analysis of the effect on bone pathology in growth plates and articular cartilage of MPS IVA mice treated with AAV8 vectors.
Pathological Scores in Bones of Mucopolysaccharidosis Type IVA Mice Treated with AAV8 Vectors
Note: Scores for vacuolization and column structure were assigned between 0 and 3 as follows: no storage (0), slight storage (1), moderate storage (2), and abundant storage (3). Slides from wild type (n = 4), KO (untreated) (n = 6), AAV8.TBG.hGLANS (n = 5), AAV8.TBG.D8.hGLANS (n = 6), AAV8.CAG.hGLANS (n = 6), and AAV8.CAG.D8.hGLANS (n = 4).The scores were given in a double-blinded manner three times.
p ≤ 0.05. Two-way ANOVA test. WT, wild type.
Bone architecture in MPS IVA mice
We examined internal bone structures, including volume and BMD, using micro-CT, allowing quantitative assessment of the treatment efficacy of AAV vectors on bone and cartilage lesions of MPS IVA mice. Micro-CT scans indicated that the density of femoral trabecular bone was higher in untreated KO male mice than in male WT mice. AAV8-TBG and -CAG vector treatments normalized bone densities in KO mice 12 weeks postinjection (Fig. 4A). Moreover, in trabecular bone, bone volume fraction (%) (BV/TV; bone volume/total volume [bone + other tissues]) and Tb.N (trabecular number implies the number of traversals across a trabecular structure made per unit length on a random linear path) of AAV-treated KO mice were reduced compared with untreated KO mice. Some of the bone parts evaluated in bone in treated KO mice showed lower values than WT animals; the explanation for that effect remains unclear. Future studies need to include a more significant number of mice and explore the potential cellular and molecular mechanisms of how this effect occurs. Even though fewer mice were evaluated in this study, the AAV8 vectors with CAG constructs showed a more effective trend in pathological findings compared to those with liver-specific TBG promoter constructs (Fig. 4B). in addition, there was a noticeable trend in BMD levels in KO mice with AAV8 vectors containing CAG constructs (Supplementary Table S2). Since the number of bones analyzed is limited, especially in mice treated with CAG constructs, an increase in samples is required for future studies.
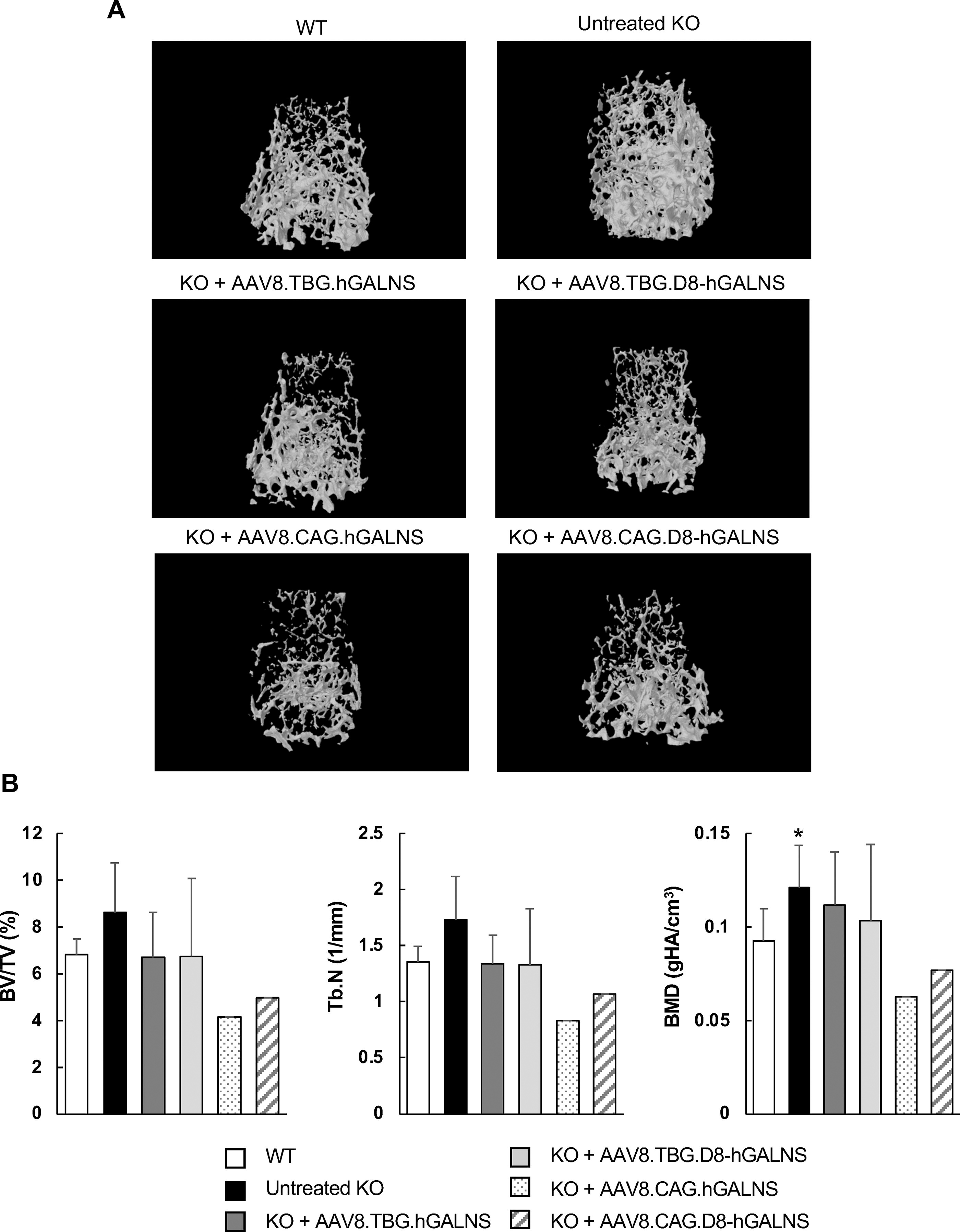
Bone architecture of MPS IVA mice treated with AAV8 vectors. Femur samples were collected from MPS IVA mice 12 weeks post-injection of AAV vectors, with or without a bone-targeting signal, to assess bone architecture using micro-computed tomography (micro-CT) analysis.
Circulating anti-hGALNS antibodies
To assess humoral responses to hGALNS transduction, we performed ELISA on plasma samples from treated and untreated MPS IVA and wild-type mice. Treated KO mice showed the presence of significantly higher levels of circulating anti-hGALNS antibodies than untreated KO mice (0.63 ± 0.47, 0.39 ± 0.31, or 0.50 ± 0.40 optical density [OD] units for KO mice treated with AAV8.TBG.hGALNS, AAV8.TBG.D8-hGALNS, or AAV8.CAG.hGALNS) (Fig. 5). The variability in the antibody levels seen between different treatment cohorts could be due to the assay artifacts where the circulating hGALNS levels influenced the detectable levels of the unbound anti-GALNS antibody levels. This could be the reason for lower OD levels for the AAV8.CAG.D8-hGALNS group with high circulating levels of hGALNS in the plasma. The detection of antibodies in KO mice could be due to the immune response to a foreign protein hGALNS enzyme as these mice are deficient or to the human sequence. As expected, in treated MTOL mice, no anti-GALNS antibodies were detected since this mouse model expressed an endogenous nonfunctional mutated GALNS and is tolerant to hGALNS inherently. 53
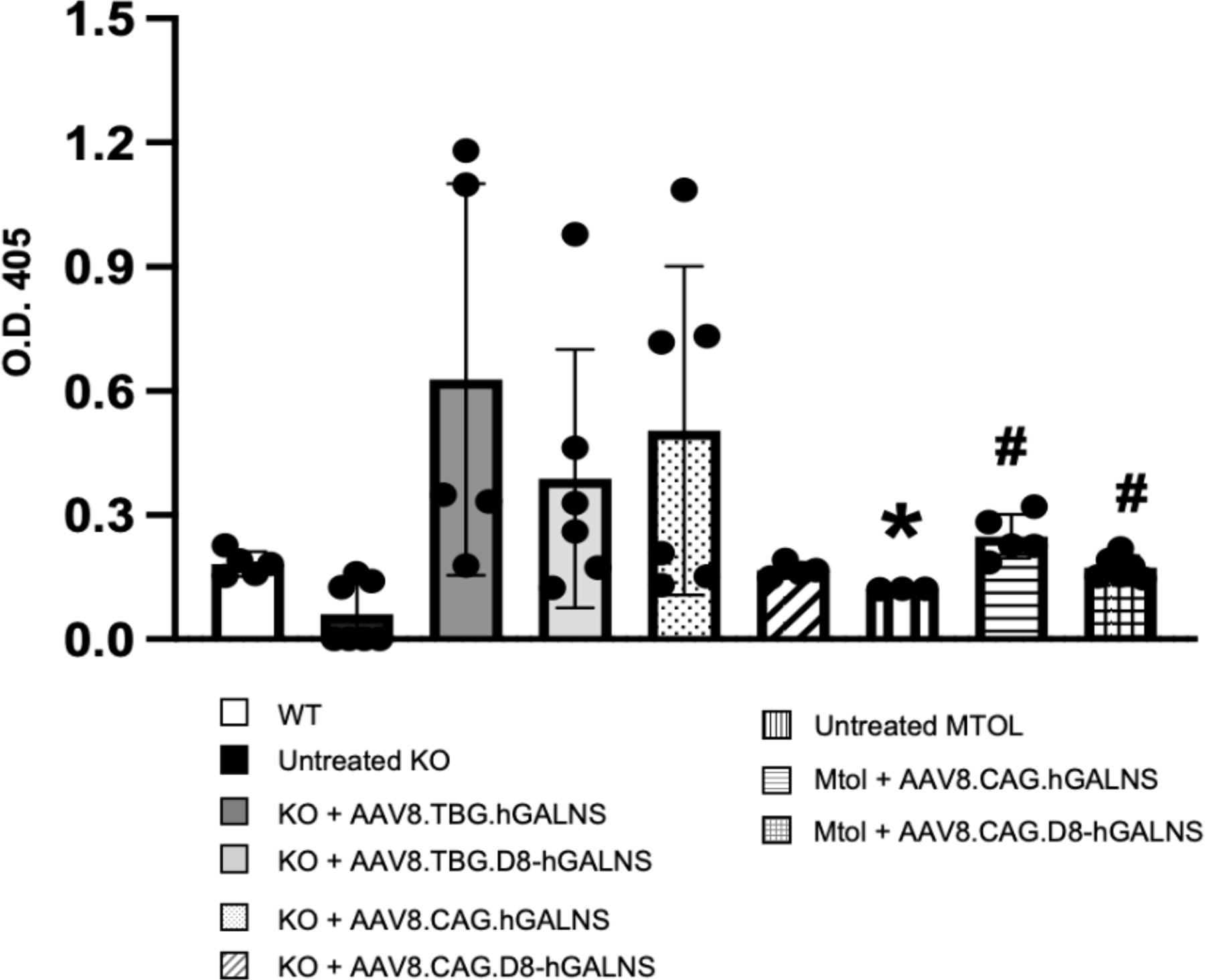
Anti-hGALNS IgG development in plasma of MPS IVA mice treated with AAV8 vectors. Plasma was collected from MPS IVA mice 12 weeks postinjection of AAV8 vectors under the control of TBG (2.0 × 1014 GC/kg dose) or CAG promoter (5.0 × 1013 GCs/kg dose), with or without a bone-targeting signal. Circulating anti-hGALNS antibody titers were detected by indirect ELISA assay, and optical density (OD) values at 405 nm were measured in a microplate spectrophotometer. n = 4–8. One-way ANOVA analyzed statistics with Bonferroni’s post hoc test. Data are presented as means ± SDs. *p < 0.05 AAV8 vectors under the control of TBG or CAG promoter vs. wild type (WT), # p < 0.05 AAV8 vectors under the control of TBG or CAG promoter vs. untreated., open bar; untreated KO, black bar; KO+AAV8.TBG.hGALNS, dark gray bar; KO+AAV8.TBG.D8-hGALNS, gray bar; KO+AAV8.CAG.hGALNS, dotted bar; KO+AAV8.CAG.D8-hGALNS, oblique striped bar; untreated MTOL, longitudinal striped bar; MTOL+AAV8.CAG.hGALNS, lateral striped bar; KO+AAV8.CAG.D8-hGALNS, cross-striped bar.
DISCUSSION
In a previous study, we found that transducing the hGALNS gene using AAV vectors controlled by a liver-specific TBG promoter moderately impacted bone and cardiovascular issues in mice with MPS IVA. 44,45 Our early research also showed that male mice treated with AAV vectors under liver-specific and ubiquitous promoters had a low antibody response against the gene product and improved overall bone pathology compared to females. 44 In this study, we tested a higher dose (2.0 × 1014 GC/kg) of AAV vectors expressing hGALNS, with or without a bone-targeting signal, controlled by a liver-specific TBG promoter. We assessed their effects on bone lesions in two established MPS IVA mouse models representing human patient genotypes. We also developed a micro-CT analysis for MPS IVA mice to evaluate therapeutic effectiveness.
Systemically administering AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS into MPS IVA model mice increased enzyme activity levels in multiple tissues, including bone, and partially improved skeletal pathology 12 weeks postinjection. Our initial hypothesis was that MTOL mice have higher or at least similar enzyme activity levels in plasma and liver compared to KO mice. Consequently, we observed differences in GALNS activity levels between the two mouse models when the vectors were controlled by the CAG promoter. Further analyses are needed, including measuring neutralizing antibodies that can inhibit the GALNS enzyme in plasma and tissues. 70
The effective delivery of active enzymes into bone lesions, especially avascular cartilage, remains challenging when using AAV-mediated gene therapy. While systemic delivery of AAV vectors with a TBG promoter has improved bone length in MPS VI feline and rat models, 42,43 the ideal AAV vector constructs for treating bone and cartilage lesions in MPS IVA remain unknown. Therefore, an AAV construct with more efficient transduction and higher promoter activity in bone should be advanced for clinical development. Recently, Lee et al. reported that the ubiquitous CAG promoter constructs packaged into AAV8 vectors provided higher levels of transgene products in the mouse bone than other AAV vectors with different serotypes or AAV8 vectors under the control of a bone-specific promoter. 71 Likewise, a study where an AAV9 vector under the CAG promoter in the MPS IVA rat model showed an increased GALNS level, preventing skeletal dysplasia and cartilage deterioration. 72,73
Yang et al. showed that different AAV serotypes had different transduction efficiencies in bone cells, including osteoblasts and chondrogenic cells. 74 This shows that AAV vectors can potentially deliver transgene products to avascular cartilage lesions from adjacent bone cells, which provides a promising approach for treating systemic skeletal disorders. Additionally, AAV vectors containing TBG constructs can deliver secreted proteins to the bone from hepatocytes through cross-correction. Moreover, a study by Chen et al. revealed that AAV2, when administered locally via intra-articular injection, showed high transduction efficiencies in chondrocytes located in the deep layer of the articular cartilage. 75 As such, there is a need for further studies to optimize the administration route of AAV vectors and identify appropriate AAV serotypes for the treatment of bone and cartilage lesions in MPS IVA mice. 44,45
In our current research, we examined the effectiveness of AAV vectors at a higher dose of 2 × 1014 GC/kg of AAV8.TBG.D8-hGALNS or AAV8.TBG.D8-hGALNS, which is four times higher than the previously used vector dose of 5 × 1013 GC/kg. This high dose is roughly equivalent to the highest vector dose used in many AAV-based gene therapy clinical trials for patients with spinal muscular atrophy, Duchenne muscular dystrophy, X-linked myotubular myopathy, and limb–girdle muscular dystrophy. 76 Our findings are as follows: 1) AAV GC numbers increased dose-dependently in the livers of MPS IVA mice treated with AAV8 compared with our previous results with the AAV8-TBG.hGALNS or AAV8.TBG.D8-hGALNS vector; 2) hGALNS enzyme activity levels dose-dependently increased in the bones of MPS IVA mice treated with the AAV8.TBG.hGALNS or AAV8.TBG.D8-hGALNS vector; 3) enzyme activities in the bones of MPS IVA mice treated with the AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS vectors were significantly higher than those receiving a 4-fold increased dose of the AAV8.TBG.hGALNS or AAV8.TBG.D8-hGALNS vectors; and 4) improvement of bone lesions was greater in MPS IVA mice treated with AAV8.CAG.hGALNS or AAV8.CAG.D8-hGALNS. Our data suggest a potential improvement in bone pathology with the aspartic acid octapeptide (D8). However, more mice are required to confirm the potential of the dual effect of D8-hGLANS. This discovery instills hope for the future of MPS IVA treatment. Therefore, it is crucial to delve into the uptake level of D8 in bone cells compared to the native GALNS enzyme. Another avenue worth exploring could be integrating D8 into the vector capsid protein to directly target the bone. 77
Advanced micro-CT analysis showed that some parameters in bone morphology are distinguishable between untreated, WT, and treated mice, which could be a potential quantitative assessment method for bone in MPS IVA mice. The number of animals analyzed in micro-CT is a limitation in interpreting the data in the present study. Therefore, in future studies, the number of individuals per group should increase, and bone mechanical strength assays must be performed to confirm the effect of the bone pathology reported here. In this study, the intravenous administration of AAV8 vectors with a CAG constitutive promoter has also demonstrated significant therapeutic potential for treating MPS IVA bone and cartilage lesions, offering a beacon of hope for the future of MPS IVA treatment. However, it is important to note that our study has limitations, including the limited number of experimental mice and the lack of an anti-AAV antibody assay. Further research is needed to understand the potential of AAV vectors in fully treating MPS IVA.
CONCLUSIONS
This preclinical study offers valuable insights into selecting the appropriate vector dose and promoter for future AAV-mediated gene therapy for humans with MPS IVA. Additional research is necessary to address and overcome the challenges presented by immune responses against the produced enzyme and/or the AAV vector to successfully implement this approach in clinical settings.
Footnotes
ACKNOWLEDGMENTS
The authors thank the support of the Austrian MPS society, A Cure for Robert, Inc., The Carol Ann Foundation, Angelo R. Cali & Mary V. Cali Family Foundation, Inc., The Vain and Harry Fish Foundation, Inc., The Bennett Foundation, Jacob Randall Foundation, Nemours Children’s Health, and REGENXBIO Inc. Special thanks to Olivier Danos, Joseph T. Bruder, and Subha Karumuthil-Melethil for supporting the study.
AUTHORS’ CONTRIBUTIONS
K.S.: Investigation in the mouse models, data curation, formal analysis, writing—original draft manuscript. K.S., M.S., S.K., M.P., and J.V.A.: Investigation. M.S., J.V.A., and S.T.: Investigation and data curation of pathological assessments of the murine models. A.M.H.P.: Writing—review and editing, formal analysis, and data curation. K.S., and S.T.: Conceptualization, methodology, and project administration. S.T. and A.M.H.P.: Visualization. S.T.: Supervision and resources.
DATA AVAILABILITY STATEMENT
The data used and/or analyzed during this study are available from the corresponding author on reasonable request.
AUTHOR DISCLOSURE STATEMENT
S.T. and K.S. declare competing financial interests: they have U.S. Provisional Patent Application nos. 62/711,238, 62/756,880, and 62/799,834.
FUNDING INFORMATION
This work was supported by grants from the Austrian MPS society, A Cure for Robert, Inc, The Carol Ann Foundation, Angelo R. Cali & Mary V. Cali Family Foundation, Inc., The Vain and Harry Fish Foundation, Inc., The Bennett Foundation, Jacob Randall Foundation, and Nemours Funds. S.T. was supported by an Institutional Development Award from the Eunice Kennedy Shriver National Institute of Child Health & Human Development of the National Institutes of Health (NICHD) (
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.