
Abstract
Sickle cell disease (SCD) is a debilitating monogenic disease originating from mutations in the hemoglobin beta chain gene producing an abnormal hemoglobin HbS. The polymerization of HbS is responsible for the sickling of erythrocytes leading to anemia and vaso-occlusive events. Gene therapy is a promising treatment of SCD, and two different gene therapy drugs, using gene editing or gene transfer, have already reached the marketing stage. There is still a need to improve the efficacy of gene therapy in SCD, particularly when using anti-sickling beta-globin gene transfer strategies, which must outcompete the pathological HbS. One possibility is to increase transduction by inhibiting lentiviral restriction factors such as interferon-induced transmembrane proteins (IFITMs). This can be achieved by the addition of cyclosporin H (CsH) during the transduction process. This strategy was applied here in CD34+ hematopoietic progenitor and stem cells obtained from cord blood (CB). A first series of experiments with lentiviral vector coding for a green fluorescent protein (GFP) gene confirmed that the addition of CsH enhanced transgene expression levels and vector copy number per cell (VCN), while CD34+ cells remained viable and functional. Notably, the production of colony-forming cells (CFC) remained unaffected unless very high VCN values were reached. In a second step, CD34+ cells obtained from the CB of newborns with homozygous (n = 2) or heterozygous (n = 1) SCD mutations were transduced with the GLOBE-AS3 lentiviral vector coding for the HbAS3 anti-sickling beta globin. As with GFP, GLOBE-AS3 lentiviral transduction was clearly enhanced by CsH, leading to VCN > 2 and therapeutic levels of expression of the HbAS3. Moreover, the process did not affect the viability or functions of CFC. The combination of CB progenitors, the GLOBE-AS3 vector, and CsH is thus shown here to be a promising approach for the treatment of SCD.
INTRODUCTION
Sickle cell disease (SCD) is a rare genetic condition, yet widespread in many countries. 1 In the great majority of cases, a single base pair point mutation (GAG to GTG) in the first exon of the hemoglobin (Hb) subunit beta chain gene HBB leads to the replacement of a hydrophilic amino acid (glutamic acid) by a hydrophobic one (valine). The resulting mutated Hb (HbS) forms high levels of polymerized Hb tetramers, leading to the morphological alterations of red blood cells (RBC) known as sickling. 2 Circulating sickled RBC are eliminated and are responsible for vaso-occlusive crises associated with anemia, extreme pain, strokes, and associated morbidities. 2,3 Several gene therapies have been developed with the objective of curing SCD with limited side effects. 4,5 Two drugs, exagamglogene autotemcel (Casgevy®—Vertex) and lovotibeglogene autotemcel (Lyfgenia®—Bluebird Bio), have recently received marketing approval by the Food and Drug Administration. 6,7 Exagamglogene autotemcel yielded significantly positive results in a phase 3 trial that enrolled 44 patients, with the disappearance of veno-occlusive events in 97% of them. 8 It is based on CRISPR/Cas9 editing of the fetal (HbF)/adult hemoglobin genetic switch in autologous hematopoietic stem/progenitor cells (HSPCs), therefore lowering the amount of Hb tetramers containing HbS in RBC. Lovotibeglogene autotemcel provided excellent results in the production of a modified anti-sickling Hb protein and reduction of vaso-occlusive crises in 35 patients. 9 It involves gene transfer with a lentiviral vector (LV) into autologous HSPCs to express a modified beta globin gene coding for the HbAT87Q beta globin mutant chain with reduced ability to polymerize. Both types of gene therapies must deliver high percentages of gene-corrected cells to provide clinical benefit; however, the latter approach only generates clinical benefit when high levels of transgene expression (i.e., high vector copy number per cell [VCN]) lead to sufficiently high levels of HbAT87Q chains to compete with the endogenously produced HbS to reduce its integration in Hb tetramers. While these two therapies exist, it is not clear at the moment whether they will be available for all patients with SCD. It is therefore important to continue developing effective gene therapy strategies for this disease.
GLOBE-AS3 is an anti-sickling LV developed for SCD gene therapy, 10,11 relying on a principle similar to that of HbAT87Q but providing a greater number of anti-sickling mutations. The GLOBE-AS3 vector codes for a mutated HbAS3 beta-globin chain carrying three such mutations (T87Q, G16D, and E22A), inhibiting axial and lateral contacts in the HbS polymer. In preclinical studies, 60–80% of mobilized CD34+ cells could be transduced by the GLOBE-AS3 vector. 10 Moreover, potentially therapeutic levels of the HBAS3 globin transgene were obtained in erythrocytes derived from the transduced cells. 10
Although these results are encouraging, the levels of HSPC transduction with LV remain a limiting factor, and solutions must be searched to improve this aspect. One option could be to inhibit the innate protection against lentiviral transduction, which is controlled by interferon-induced transmembrane proteins (IFITMs) and which has been identified as a restriction mechanism that is especially active in HSPCs. 12 IFITM restriction can be overcome by cyclosporin H (CsH), recently identified as an adjuvant enhancing the LV transduction of HSPC. 12 –14
The Drepacord study (Clinicaltrials.gov NCT 03876821) aims at evaluating the potential of cord blood (CB) HSPC for SCD gene therapy as an alternative or as a complement to the use of peripheral mobilized CD34+ cells. Here, CB collected in the Drepacord study was used to explore if CsH could enhance the transduction of CB CD34+ cells with the GLOBE-AS3 LV vector. High VCN were indeed achieved without evidence of toxicity, predicting the possibility to reach therapeutic levels of anti-sickling Hb in erythroid cells.
MATERIALS AND METHODS
Cells
The human cell lines HEK293T (Lenti-X cell line, Takara Bio Inc., Japan) and HCT116 (ATCC, Manassas, VA), respectively, used to produce and to titer LV, were maintained in the laboratory as reported elsewhere. 15 CB samples were collected following uncomplicated births at the Centre Hospitalier Sud-Francilien (CHSF) hospital (Corbeil-Essonnes, France) and used for preliminary calibration assays testing the number of transfection cycles, the dose of LV, and the impact of CsH. CD34+ CB cells were also obtained in the C18-31 Drepacord study sponsored by Inserm, which was granted approval by a local Ethics Committee on December 12, 2018, and registered in a public trials registry (Clinicaltrials.gov NCT 03876821). The Drepacord study collects, at CHSF, CB samples from individuals at risk of presenting the HbS genotype. Three such CB samples were obtained, one heterozygous (HBB A/S, MG28) and two homozygous (HBB S/S, NA27, and CT32) (Supplementary Table S1). Genotyping of samples from the Drepacord protocol was performed by sequencing HBB gene exon 1–2 and exon 3 sequences amplified by PCR (see primers in Supplementary Table S2). For all CB collected and used for transduction experiments, CD34+ cells were prepared by magnetic bead cell sorting according to the manufacturer’s instructions (Miltenyi Biotech, Bergisch Gladbach, Germany).
Lentiviral vectors
A lot of LV coding for the green fluorescent protein (GFP) gene from the EF1 short promoter and pseudotyped with vesicular stomatitis virus G (VSVG) was prepared in preclinical grade conditions relevant to the conduct of clinical trials. This LV was produced by transient transfection of HEK293T cells, concentrated and purified by several membrane and chromatography steps, as described elsewhere 16 resuspended in TSSM (tromethamine 20 mM, NaCl 100 mM, sucrose 10 mg/mL, and mannitol 10 mg/mL) buffer, and cryopreserved before use. The LV titer, measured on HCT116 cells using digital droplet polymerase chain reaction (ddPCR) and provirus-specific primers, and probes 15 was 5E + 09 infectious genome (IG)/mL.
The GLOBE-AS3 LV described elsewhere, 10 encodes the HbAS3 transgene under the control of the beta-globin promoter and of a reduced version of the beta-globin locus-control region and is pseudotyped with VSVG. A lot of the GLOBE-AS3 LV was produced at Genethon using preclinical conditions as reported 17 and provided for this study. Briefly, production of the vector was performed in a 200 L suspension of adapted HEK293T cells in a bioreactor with serum-free medium. The vector was then purified by ion exchange chromatography and tangential flow filtration as reported, 16 formulated in X-VIVO 20 medium (Lonza, Basel, Switzerland), and titrated at Genethon on HCT116 cells at 1.3E + 09 IG/mL, a titer confirmed using the same method as with the GFP LV above.
Transduction and efficacy assessment
CD34+ cell transduction was performed as previously reported. 18 Briefly, CD34+ cells (1E + 06 cells/mL) were cultured in X-VIVO15 serum-free medium without phenol red (Lonza) supplemented with penicillin (100 U/mL)/streptomycin (100 µg/mL) (Gibco, Thermo Fisher), ckit-ligand (50 ng/mL), FLT3-ligand (100 ng/mL), interleukin-3 (20 ng/mL), and thrombopoietin (50 ng/mL) (Miltenyi Biotech). After 18–24 h of culture, transduction was initiated by adding the LV at the indicated concentration in the presence of protamine sulfate (8 µg/mL) (Sigma Aldrich, Saint Louis, MO) and with or without CsH (Sigma Aldrich) used at 8 µM as described elsewhere. 12 Six hours later, the medium was changed, and cells were either plated in methylcellulose (see below) or cultured in X-VIVO15 medium containing cytokines and 10% heat-inactivated fetal bovine serum (FBS) (Cytiva, Pasching, Austria) for several days to measure cell viability or transgene levels. When two cycles of transduction were performed, the cells that were washed after the first cycle of transduction were transduced again in the same conditions but incubated for 18 h (overnight), then were washed and cultured either in methylcellulose or in a medium supplemented with FBS as above.
HBB S genotype was determined by PCR amplification (primers described in Supplementary Table S2) and sequencing of the PCR product (Genwiz, Leipzig, Germany).
VCN were measured by ddPCR using provirus-specific primers and probes as reported elsewhere. 15
Cell viability was evaluated by trypan blue staining on a hemocytometer by microscopic observation.
The ability of transfected cells to differentiate into lineage-specific colony-forming cells (CFC) was evaluated after seeding on methylcellulose plates (Methocult optimum, Stemcell Technologies, Vancouver, Canada). Automated CFC counting was performed with the Stemvision® enumeration system (Stemcell Technologies) and analyzed with the “Human Cord Blood 14-day” software. The different categories of lineage-differentiated CFC, burst-forming units erythroid (BFU-E), CFU-granulocyte erythrocyte monocyte megakaryocyte (CFU-GEMM), and CFU-granulocyte monocyte (CFU-GM) were aspirated to analyze VCN in the pooled colonies of each CFC category. Genomic DNA from the different pools of CFC was extracted using the Maxwell RSC cell DNA kit and instrument (Promega, Madison, WI, USA). The production of Hb in BFU-E was assessed by high-performance liquid chromatography (HPLC) as previously reported. 19
Flow cytometry
Flow cytometry was conducted on an Attune NxT cytometer (Thermofisher Scientific, Courtaboeuf, France), analyzing data with the FlowJo V10.7.2 analysis software (Becton Dickinson, Ashland, OR).
The expression of GFP or markers was analyzed on fresh cells, and dead cells were excluded by 7AAD staining. Cell surface markers were detected with fluorochrome-conjugated monoclonal antibodies used at the indicated dilutions: human CD45-BV421 (1/20) and human CD3-BV605 (1/20) (Biolegend, San Diego, CA); human CD19-PE-Vio770 (1/50), human CD4-Vio-Bright667 (1/50), and human CD8-APC-Vio770 (1/50), Miltenyi Biotec.
Intracellular IFITM3 expression was measured on fixed and permeabilized cells using the fixation/permeabilization solution kit (Invitrogen, ThermoFischer) then staining with a monoclonal mouse antihuman IFITM2/3 antibody (clone 2E8D12) used at 1/200 dilution (Proteintech, Manchester, UK) revealed by a goat anti-mouse IgG—Alexa Fluor 488 used 1/200 (Invitrogen).
Statistics
Statistical analyses were performed with the Prism 9.3.1 software.
RESULTS
Optimization of CD34 CB HSPCs transduction with a purified GFP LV and CsH
In the literature, fluorescent microscopy and Western immunoblotting techniques were used to show the expression of the lentiviral restriction factor IFITM3 in CD34+ cells and its reduction by CsH. 12 To obtain a more quantitative measure, we used intracellular flow cytometry with an already reported monoclonal antibody 20 recognizing both IFITM2 and IFITM3 that are highly similar proteins displaying the same antiviral activities. The flow cytometry staining technique was validated on IFITM3-transduced 293T cells treated or not with CsH (Supplementary Fig. S1) and used on CD34+ cells cultured in conditions mimicking a double transduction protocol. As shown in Figure 1A and B, a high percentage of cytokine preactivated CD34+ cells express IFITM2/3 at the start of the experiment (0 h time point), and levels remain high throughout the culture period without significant changes over time. The percentage of IFITM2/3+ cells is drastically reduced by CsH within 6 h (from 70 ± 12% to 6 ± 0.8%, respectively), which is the period of time that would correspond to a first transduction cycle (first LV would be added at 0 h time point). The levels remain significantly reduced between 6 h and 24 h ending with a 50% reduction in IFITM2/3+ cells in CsH-treated conditions compared to untreated cells (53 ± 14% vs. 93 ± 4%, respectively), and corresponding to the time frame of a second transduction cycle (second LV would be added at the 6 h time point) (Fig. 1A and B). The effects of CsH on the lentiviral transduction of CD34+ CB HSPCs were then assessed by testing various transduction conditions using 1 or 2 cycles of transduction and different concentrations of a GFP LV. The significant enhancement of transduction levels by CsH is exemplified in Figure 2A and B. A vector dose-dependent enhancement of transgene levels is observed, reaching almost 100% of GFP+ cells after a double transduction (2x) with 5E + 07 IG/mL, followed by a plateau effect (Fig. 2A). The effects of CsH also translate into higher VCN in all conditions tested (Fig. 2B). Interestingly, 10 VCN or more could be obtained with two transduction cycles. This is a strikingly high level, considering that 2 VCN is the expected minimal level required for SCD disease correction with LV. As shown in Figure 2C, neither the number of cycles of transduction (1 vs. 2), nor the vector dose or CsH presence impacted cellular viability, as measured. Different types of CFC, respectively, GEMM, GM, and BFU-E, were generated by CD34+ cells transduced in the presence or absence of CsH (Fig. 2D). While a statistical analysis revealed no significant differences in CFC formation between all conditions examined globally, focusing only on high transduction conditions showed that CsH slightly reduced CFC counts when a double transfection with the highest dose of vector (1E + 08 IG) was used (Fig. 2D). This effect is coherent with the very high VCN obtained in these conditions.
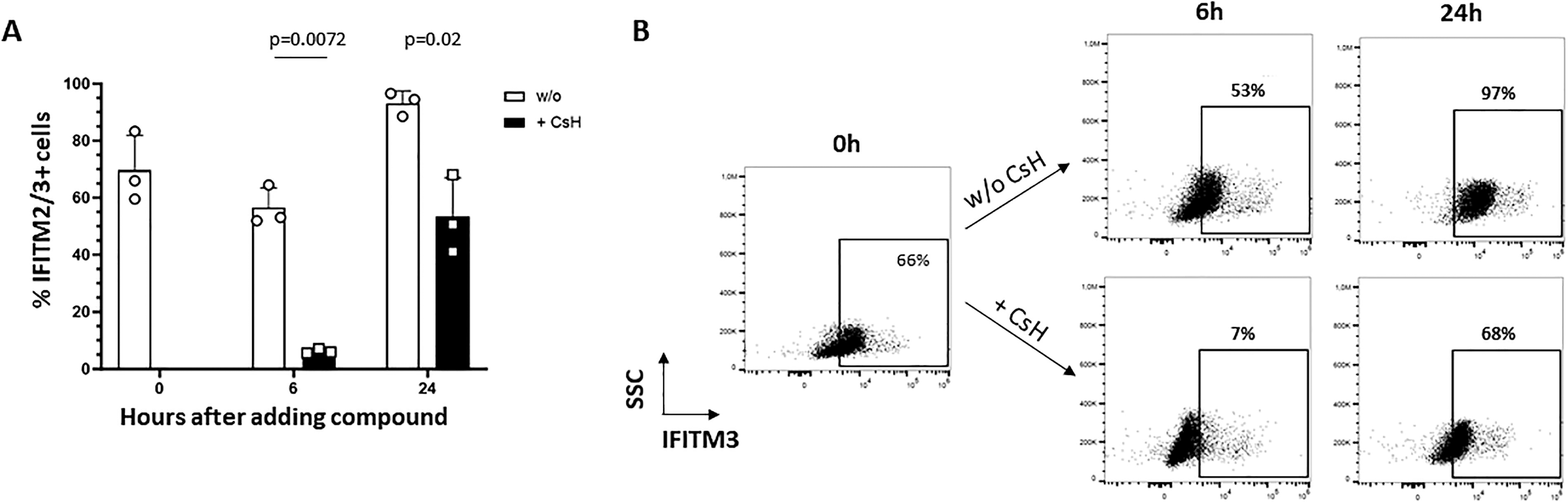
CsH decreases the percentage of IFITM2/3+ CD34+ cells. Human CB CD34+ cells (3 donors) were prestimulated overnight with a cocktail of cytokines and then cells were stimulated or not with CsH (8 µM) for 0, 6, and 24 h. At the indicated time points, IFITM2/3 expression was assessed by intracellular flow cytometry. Background staining with the secondary antibody only was <2%.

Optimization of the transduction of CB CD34+ cells with a GFP vector, using CsH and repeated transduction. A preclinical grade purified lentiviral vector coding for GFP was used at the indicated concentrations (corresponding to MOI of 10, 50, and 100, respectively) to transduce CB CD34+ cells (n = 3 donors in two experiments) during 1 (1x) or 2 (2x) cycles of transduction, in the presence (+) or absence (w/o) of CsH.
Optimization of CD34+ HSPCs transduction with the GLOBE-AS3 LV
Based on results obtained with the GFP LV, the GLOBE-AS3 vector was tested using two cycles of transduction, and results are shown in Figure 3. Transduction levels were lower than with the GFP vector, due to the larger size of the GLOBE-AS3 provirus (6.3 kb vs. 3.6 kb for the GFP LV). CsH significantly enhanced the levels of transduction with the GLOBE-AS3 vector (Fig. 3A). Two cycles of transduction with CsH were needed to reach the 2 VCN target therapeutic level with 3- and 5E + 07 IG/mL of vector (Fig. 3A). As expected from the GFP results, transduction impacted neither cell viability (Fig. 3B) nor CFC generation (Fig. 3C) considering the levels of transduction obtained. The transduction-enhancing effects of CsH and lack of negative impact of CsH on viability and clonogenicity were also confirmed with one cycle of transduction but these conditions could only reach a maximum value of VCN 1, which is suboptimal for gene correction (Supplementary Fig. S2).

Transduction of CB CD34+ cells with the GLOBE AS3 LV. The purified GLOBE-AS3 LV was used at the indicated concentrations to transduce twice CB CD34+ cells of 2–4 donors in 2–3 experiments, in the presence or absence of CsH.
Test of the GLOBE-AS3 vector on patient samples from the Drepacord study
The Drepacord study provided 3 CB samples with either heterozygous A/S (MG28) or homozygous S/S HBB genotype (NA27, CT32) (Supplementary Table S1), and the CD34+ cells were transduced with the GLOBE-AS3 vector. As shown in Figure 4A, the addition of CsH enhanced transduction levels in all of these samples. Higher transduction levels were obtained after 2 cycles of transduction, especially in sample CT32, in which 5 VCN were reached. Figure 4B confirms that CsH leads to stable higher levels of VCN in erythroid precursor as measured in BFU-E 14 days post-transduction. With MG28 cells, Figure 4A and B also confirm the greater efficacy of two cycles of transduction by comparing the effects on the same sample. Figure 4C and D confirms that transduction with the GLOBE-AS3 and CsH does not impact cell viability and CFC generation even with two cycles of transduction in the conditions that were used. Of note, the lower efficiency noted in NA27 cells relates to the fact that these homozygous S/S cells were transduced only once, due to an experimental constraint. The induction of the HBAS3 transgene by the GLOBE-AS3 LV was confirmed by DNA sequencing of the transduced erythroid cells (Supplementary Fig. S3). In addition, the various Hb expression profiles were examined by HPLC in erythroid cells differentiated from transduced CD34+ cells from the three patients, confirming the detection of HbAS3 chains in transduced samples (Fig. 5 and Supplementary Table S3). As shown in Figure 5, the levels of therapeutic HbAS3 (which competes with HbS) become at least greater or equal to those of HbS when at least a VCN of 2, and ideally of 4, is achieved in the cells after two cycles of transduction (MG28 and CT32) (Supplementary Table S3). As shown with the HbS/S CT32 sample, two cycles of transduction with CsH provide more than 70% of anti-sickling Hb of all beta globin expressed in erythroid cells, values expected to have therapeutic effects.
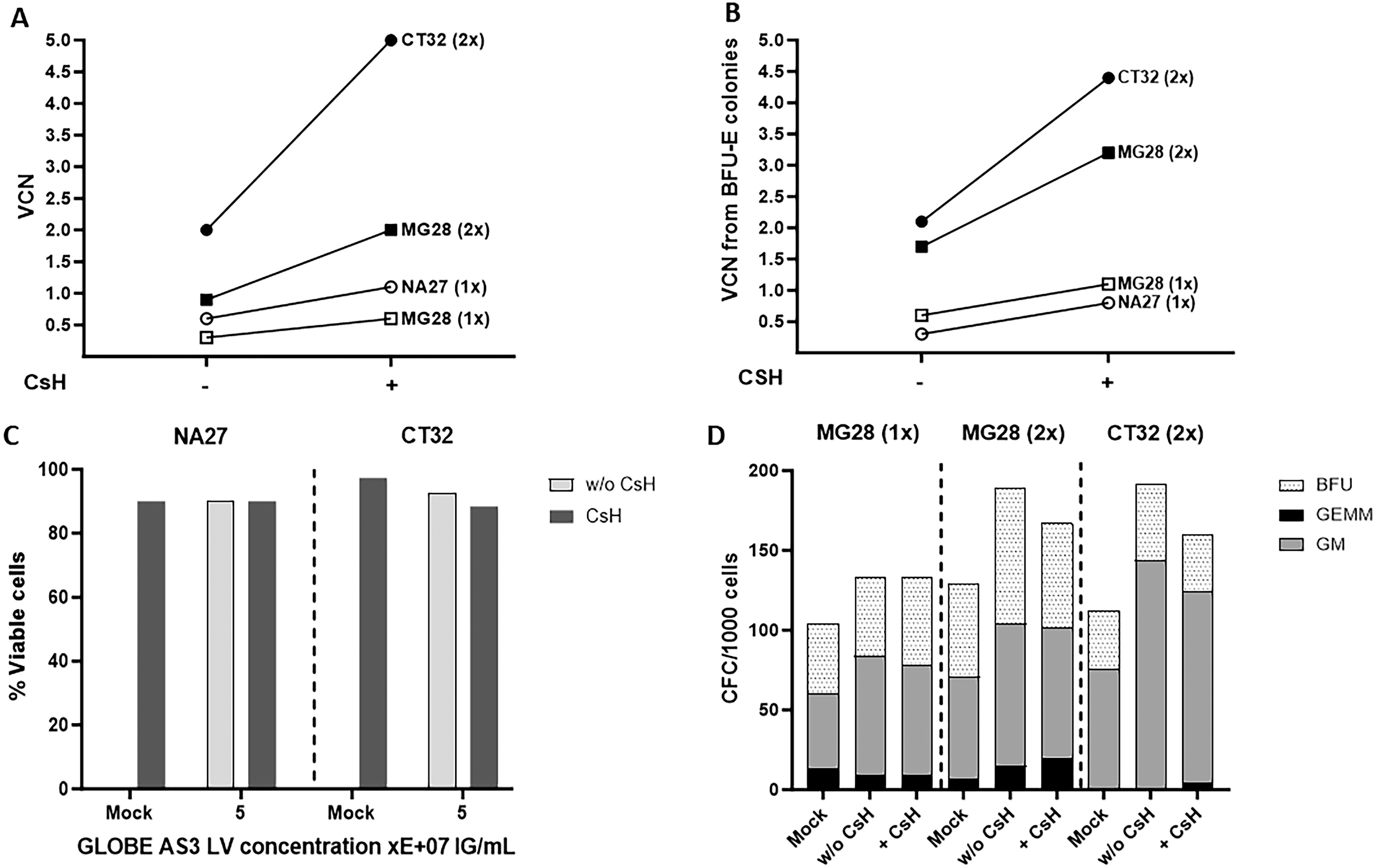
Transduction of CD34+ cells from Drepacord samples with the GLOBE-AS3 vector. The purified GLOBE-AS3 LV was used 1x or 2x at 5E + 07 IG/mL (MOI = 50) with or w/o CsH to transduce Drepacord CB CD34+ samples, as indicated. Mock corresponds to untransduced cells exposed to cytokines and CsH but not LV. Results for the indicated samples and conditions show:
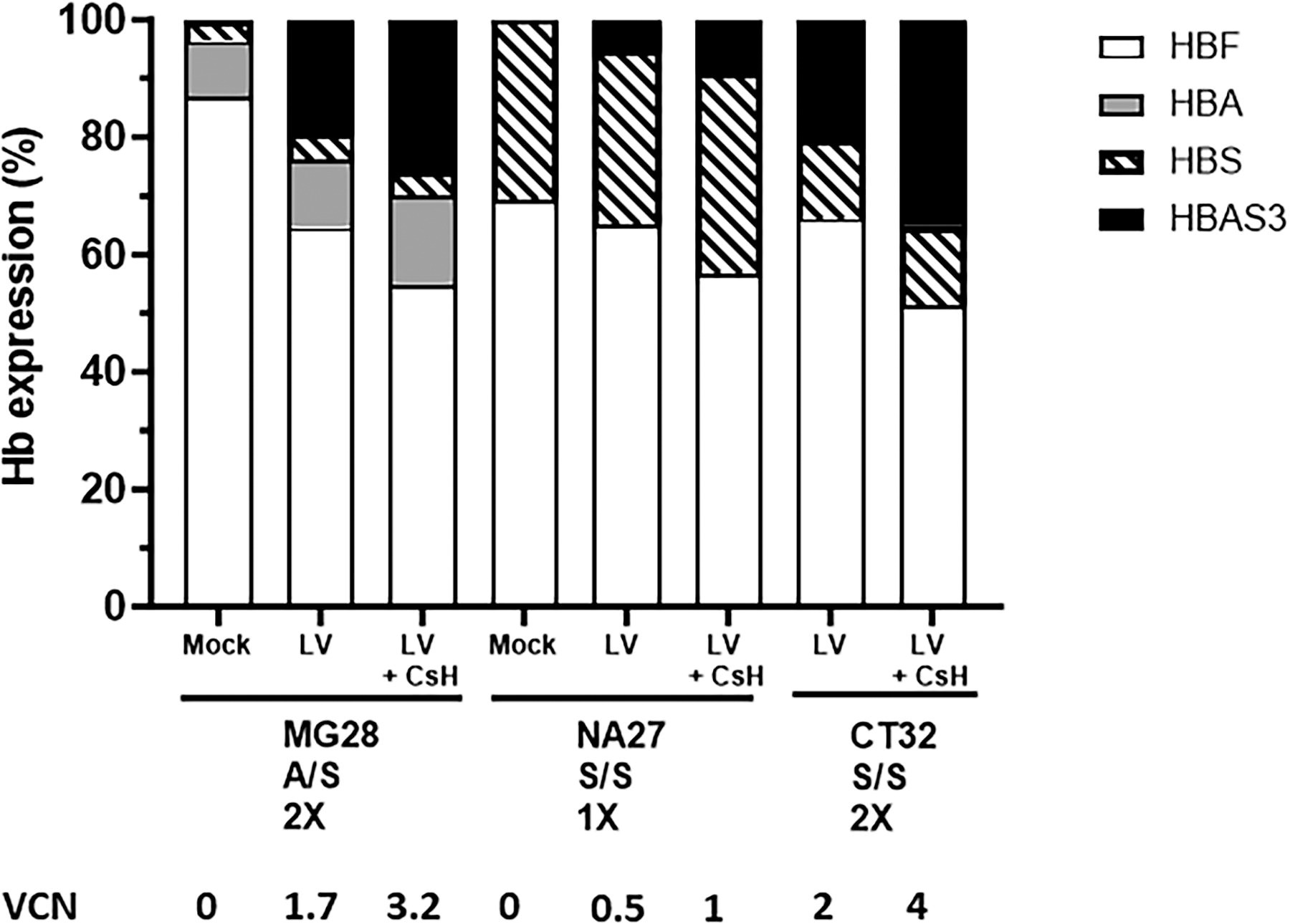
Globin expression by CD34+ cells from the Drepacord protocol and transduced with the GLOBE-AS3 vector. The indicated samples were transduced with 1 or 2 cycles of 5E + 07 IG/mL (MOI = 50) of the GLOBE-AS3 LV in the presence or absence of CsH and cultured in methylcellulose for 14 days. Bulk BFU-E were collected and pooled to measure the expression of the various types of hemoglobin using HPLC. VCN were measured by ddPCR in day 14 bulk BFU-E genomic DNA.
DISCUSSION
The Drepacord study enabled us to test a combination of approaches to improve the efficiency of gene correction in CB HSPCs for the gene therapy of SCD. Here, the anti-sickling GLOBE-AS3 LV vector was used in combination with the CsH adjuvant, demonstrating the possibility to improve the efficacy of transduction to reach values of at least two copies of transgene per cell using less vector, values that provide putatively protective levels of anti-sickling Hb. A first series of experiments in CD34+ cells confirmed the strong reduction of cells expressing the antiviral IFITM2/3 proteins obtained with the CsH adjuvant, enabling their enhanced transduction with impressive VCN and limited toxicity. CsH moreover allowed to use lesser amounts of LV vector by displaying a plateau effect. This confirmed data reported by several groups 12,14 in a different context, highlighting the global efficacy of CsH in counteracting innate mechanisms of antiviral protection for efficient transduction of quiescent cells. Our results are consistent with the implication of IFITM3 to restrict lentiviral transduction in CD34+ cells but do not exclude the participation of other innate restriction factors.
The Drepacord study provided several CB CD34+ cell samples with HBB S/S homozygous or A/S heterozygous genotype, demonstrating that the large size GLOBE-AS3 vector combined with CsH could provide a satisfactory level of 3–4 VCN, allowing to reach potentially therapeutic levels of HbAS3 production. Excessively high VCN values were not attained with CsH and the globin vector, therefore not posing a concern for toxicity and complying with usual safety recommendations for cell and gene therapy. Our results in CB S/S cells with CsH showed that VCN of four provide 70% of HbAS3 over total beta globin and therefore compare favorably to those of a previous preclinical study using the GLOBE-AS3 LV in adult mobilized peripheral blood cells or bone marrow in which at most 2 to 3 copies of vector were obtained and provided about 25–30% of HbAS3 over total beta globin translating into 35% nonsickling erythrocytes derived from these transduced cells. 10 Thus, therapeutic efficacy could be reasonably expected from HSPC in which VCN of 2–4 are reached with anti-sickling GLOBE-AS3 vector.
In addition to its ability to enhance the efficiency of transduction, CsH has previously been reported to preserve cell viability. 14 Our data confirmed published studies showing that HSPC treated with CsH appear to retain normal hematopoietic differentiation potential. 12,13 With the GFP vector, CsH generated high VCN but the hematopoietic differentiation potential of transfected cells to form all types of CFC was not altered unless very high levels of VCN were attained, in the range of 10 copies of vector per cell or more. It is generally considered that VCN values above 5 may introduce a risk of genotoxicity and loss of hematopoietic activity; therefore, robust transduction protocols must be developed to obtain VCN values between 2 and 5, a therapeutic range for anti-sickling beta-globin vectors. In that range of VCN, we obtained preliminary data confirming that CsH-treated cells can engraft in immunodeficient mice to generate multiple lineages of human cells as shown in Supplementary Figure S5 with a GFP vector. This confirms the lack of gross hematopoietic toxicity by CsH reported by others who also showed that CsH does not affect primitive HSPC, which can generate secondary transplants. 12 Moreover, the group of Kajaste-Rudnitski even further improved CD34+ cell transduction by adding deoxynucleotides to enhance quiescent hematopoietic stem cell transduction, preserving their long-term engraftment potential. 13 This suggests that additional optimization may still be possible to further improve the outcome of SCD gene therapy. To our knowledge, CsH is not yet available in a good manufacturing production grade for clinical use. However, this reagent would be useful as it improves transduction efficacy and allows for a reduction in the amount of vector used, yielding even less risk for toxicity and subsequently improving manufacturing feasibility. It also decreases production costs.
In conclusion, this preclinical study provides highly encouraging data for an improved efficiency of HBAS3 transduction and production and could be further developed within the regulatory recommendations for human applications of gene therapy in SCD.
Footnotes
ACKNOWLEDGMENTS
The authors would like to thank the participants of the Drepacord study (Clinicaltrials.gov NCT 03876821) and the staff of the Centre Hospitalier Sud-Francilien (CHSF) maternity unit, notably Delphine Lavergne, Emmanuelle Quargnolo, and Ludivigne Roux, for their contribution to this study. Thanks also extend to the Inserm Clinical Research Department for sponsoring the Drepacord study. The authors are grateful to Genethon for providing the GLOBE-AS3 vector. They are also thankful for the technical support from their colleagues Axel Gosseye, Antoine Biek, and Ajanthan Nesarajah. Writing for this article was assisted by MC Béné (MPIYP, Paris, France).
AUTHORS’ CONTRIBUTIONS
M.M.: Methodology, investigation, and formal analysis. L.R.: Resources. A.C.: Investigation. A.M.: Methodology, investigation, and supervision. C.F.: Investigation. S.P.: Investigation. M.D.: Resources and investigation. A.G.: Conceptualization, methodology, formal analysis, resources, writing original draft/review and editing, supervision, and project administration.
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
This study has been funded by Inserm, by the French National Research Agency (
SUPPLEMENTARY MATERIAL
Supplementary Data S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.