
Abstract
Our previous research has demonstrated that the oncolytic vaccinia virus encoding Aphrocallistes vastus lectin (oncoVV-AVL), an oncolytic vaccinia virus engineered to carry the AVL, exhibits potent cytotoxic effects on colorectal and hepatocellular cancer cells. Based on this foundation, we undertook a series of experiments to explore its efficacy on gastric cancer (GC) cells. Our findings revealed that oncoVV-AVL significantly increased reactive oxygen species levels and suppressed the expression of nuclear factor erythroid 2-related factor 2, thereby enhancing viral replication and disrupting the cellular redox balance, ultimately leading to the demise of cancer cells. Additionally, our investigations uncovered that oncoVV-AVL reprogrammed the metabolic microenvironment to favor viral replication, culminating in the lysis of cancer cells. Furthermore, we observed that oncoVV-AVL not only regressed tumor growth but also induced tumor tissue necrosis. These promising results suggest potential new avenues for the therapeutic management of GC.
INTRODUCTION
Gastric cancer (GC), the fifth most prevalent cancer worldwide, typically diagnosed in its advanced stages, has the second highest mortality rate. 1 Each year, the global tally of GC diagnoses approaches one million, with the majority of cases concentrated in South America, Eastern Asia, and Eastern Europe. 2 Advances in early diagnosis, coupled with the continuous refinement of endoscopic techniques and surgical approaches, alongside the development of potent chemotherapeutic regimens, have collectively enhanced the survival prospects for patients afflicted by this malignancy. Nevertheless, the prognosis for those battling advanced stages of GC remains distressingly bleak. The sole current treatment offering a possible cure is the complete excision of the tumor. 3 Despite undergoing such resection, patients with extensive lymph node involvement, as indicated by surgical pathology, face a bleak 5-year survival rate of merely 7%. 4 The challenges of distant metastasis or recurrence loom large. While chemotherapy emerges as a formidable ally, bolstering the efficacy of surgical intervention, its curative potential is inherently limited. 5 The majority of GCs exhibit resistance to existing chemotherapy protocols, underscoring the urgent need for novel therapeutic strategies.
Oncolytic viruses (OVs) are novel cancer therapeutics characterized by their ability to selectively infect and effectively lyse tumor cells without harming normal cells. 6 Unlike traditional viral vectors employed in gene therapy, OV therapy serves a dual role: not only functioning as a delivery vehicle for foreign genes but also acting as an anticancer agent, directly lysing and eradicating tumor cells through boosting cell antitumor immunity, which reshapes the tumor microenvironment by multiple mechanisms. 7 This dual function of OVs positions them as a novel multimodal immunotherapy strategy for cancer, with OV-induced antitumor immunity playing a pivotal role in enhancing their overall therapeutic impact. 8 As of now, there are four approved OV products on the market, including RIGVIR (ECHO-7 virus) for patients with melanoma, Accuray (H101) for head and neck tumors, herpes simplex virus (HSV-1) for patients with advanced melanoma, and Teserpaturev/G47Δ (Delytact) for malignant gliomas. 9 –12 Beyond these, a diverse array of engineered viruses—derived from vesicular stomatitis virus, HSV, adenovirus, mammalian orthoreovirus, New Castle virus, vaccinia virus (VV), and measles virus—are in various stages of preclinical or clinical investigation, underscoring promising prospects of OV research and therapy. 13 VV, a DNA virus encoding ∼200 genes, has several advantages, including a brief life cycle and swift dissemination, robust lytic capabilities, inherent systemic tumor targeting, a capacity for expressing large exogenous genes, exceptional safety profile, and well-defined molecular biology. 14,15 It can encode several immunomodulatory genes, which aid the virus in overcoming innate and adaptive immune responses. 16 Additionally, VV replicates in the cytoplasm, avoiding the risk of integration into the host genome. 17 Some OV therapies based on VV are undergoing clinical trials, including GL-ONC1, ASP9801, and RGV004, whereas others are in preclinical research. 18 Jun et al. demonstrated that a VV artificially modified to carry human sodium iodide symporter (hNIS) gene, designated as GLV-1h153, effectively induced diminishment of gastric tumors in a murine xenograft model while also facilitating deep-tissue imaging. 19 Wang et al. revealed that a recombinant VV WR strain, engineered to express SurT34A and FilC, proved to be both safe and efficacious in combating GC. 20 These findings collectively underscore the promising safety and therapeutic efficacy of engineered VVs for cancer treatment.
Lectins, a diverse group of carbohydrate-binding proteins, are adept at recognizing and binding to specific sugar molecules present on the surface of cells. These proteins are integral to a myriad of biological processes, including immune responses, cell adhesion, and intercellular communication. Notably, aquatic lectins, such as sialic acid-binding lectins from Rana catesbeiana, and agglutinin of Eucheuma serra have been shown to exhibit cytotoxic effects against cancer cells. 21,22 In our previous research, we uncovered that oncolytic VVs engineered to express lectins from Tachypleus tridentatus, white-spotted charr, Asterina pectinifera, and Aphrocallistes vastus demonstrated potent antitumor activity. 23 Of interest, the oncolytic VV incorporating Aphrocallistes vastus lectin (oncoVV-AVL) demonstrated remarkable efficacy in combating colorectal cancer and cervical cancer. 24,25 Nonetheless, the function of oncoVV-AVL in GC cells remains unclear.
In this research, we evaluated the therapeutic potential of oncoVV-AVL as an oncolytic agent for GC. We explored the mechanism by which oncoVV-AVL inhibits the growth of GC cells through cell and animal experiments and explained how oncoVV-AVL affects the viral propagation ability in GC cells at the metabolic level.
MATERIALS AND METHODS
Cell culture
The GC cell lines NCI-N87 and MKN-45 were procured from Zhong Qiao Xin Zhou Biotechnology Co., Ltd. in Shanghai, China, and MeisenCTCC in Zhejiang, China, respectively. These cells were cultured in RPMI-1640 medium, sourced from Bio-channel in Nanjing, China, supplemented with 10% fetal bovine serum from ExCell Bio in Shanghai, China, and 1% penicillin–streptomycin also from Bio-channel in Nanjing, China. The cells were cultured in a stable environment with 5% CO2 at a temperature of 37°C.
Cell viability assay
The MKN-45 and NCI-N87 cells, at a density of 8 × 103 cells per well, were seeded into 96-well plates, each with six repetitions, and were allowed to adhere to the plate walls overnight. oncoVV-AVL and oncoVV both were recombinated from the VV WR strain; the former deleted only the thymidine kinase (TK) gene, whereas the latter inserted the AVL gene and deleted the TK gene. 25 The formula for calculating the multiplicity of infection (MOI) is as follows: MOI = Total PFU of virus (used)/Number of cells. The infectivity titer is quantified using plaque-forming units (PFU), with the unit of infectious titer typically expressed as PFU/mL. The virus at MOI of 0.5, 1, 2, and 5 was introduced into the cultures. Following incubation periods of 24, 48, or 72 h, 20 μL of 5 mg/mL MTT solution (supplied by Beyotime Biotechnology) was added to each well, and the plates were incubated at 37°C for 4 h. Subsequently, the supernatant was replaced with 150 μL dimethyl sulfoxide to fully dissolve the formazan crystals. The absorbance at 570 nm was then measured using a microplate reader (Multiskan, Thermo Fisher Scientific) to assess the cell viability.
Viral yields assay
The MKN-45 and NCI-N87 cells, at a density of 1 × 105 cells, were seeded in a 24-well plate. After incubating for 12 h, the cells were treated with oncoVV and oncoVV-AVL (2 MOIs). The cells were then collected after 24, 36, and 48 h and stored at −80°C. Following three cycles of freeze-thawing, viral yields were measured by the TCID50 assay in HEK293A cells.
Flow cytometry
The MKN-45 and NCI-N87 cells, at a density of 5 × 105, were seeded into a 6-well plate. After an incubation period of 12 h, the cells were treated with oncoVV or oncoVV-AVL at a specified MOI. After 48 h, the cells were collected, washed twice with phosphate-buffered saline (PBS), and then stained with the FITC Annexin V Apoptosis Detection Kit (BD Biosciences). Flow cytometry analysis was performed using an appropriate volume of 1 × binding buffer. To detect reactive oxygen species (ROS) levels, the cells were incubated with 10 μM DCFH-DA (Solarbio, CA1410) in 1 mL of serum-free medium at 37°C for 20 min. For autophagy investigation, cells were stained with an MDC (dansylcadaverine) probe. Subsequently, the cells were washed three times with serum-free medium and resuspended in 200 μL of PBS. The data were collected from the flow cytometry (BD Biosciences) and analyzed using FlowJo10 software.
Western blotting assays
MKN-45 and NCI-N87 cells were treated with oncoVV or oncoVV-AVL for 36 and 48 h. Posttreatment, the cells were collected, lysed, and subjected to SDS-PAGE electrophoresis for protein separation, followed by transfer to a PVDF membrane. The immunoblotting analysis involved the use of primary antibodies diluted 1:1000, which included rabbit polyclonal antibodies against GAPDH, PI3K, phosphorylated PI3K, Akt, phosphorylated Akt, Yes-associated protein (YAP), phosphorylated YAP, microtubule-associated proteins light chain 3 (LC3), and nuclear factor erythroid 2-related factor 2 (NRF2), all from Cell Signaling Technology. Additionally, a rabbit polyclonal antibody against A27L was sourced from Abcam Plc. The secondary antibodies, diluted 1:4000, consisted of goat antirabbit and goat antimouse antibodies, obtained from ABclonal. The immunoblots were subsequently visualized using a chemiluminescence image system with the ECL enhanced plus kit (ABclonal, RM00021) for detection.
Animal experiment
The female BALB/c nude mice, aged 6 weeks (Shanghai SLAC Laboratory), were inoculated with MKN-45 cells at a concentration of 3 × 106 cells per mouse, which were suspended in 100 μL of 0.9% NaCl solution and subcutaneously injected into the right flank. When the tumor size reached approximately 200 mm3, the mice were randomly assigned to three distinct groups. Each group received a different intratumoral injection: 100 μL of 0.9% NaCl, oncoVV, or oncoVV-AVL (1 × 107 PFU). The tumor growth was monitored and recorded every 5 days, with the tumor volume calculated using the formula: Tumor volume (mm3) = 1/2 × length (mm) × width2 (mm). After 30 days, the mice were euthanized, and the tumors were harvested for further analysis.
Hematoxylin–eosin staining and immunohistochemistry
Immunohistochemistry (IHC) and hematoxylin
Targeted metabolome analysis
Targeted metabolome analysis was conducted by Shanghai BAYP Biotechnology Co., Ltd. In brief, cell samples were mixed with a methanol/acetonitrile/water (2:2:1) solution at a 20-fold volume ratio, followed by centrifugation at 14,000 g for 20 min to remove proteins and particles. The analytical phase used a triple quadrupole mass spectrometer (QTRAP 5500, AB SCIEX) in conjunction with an ACQUITY UPLC BEH amide column (1.7 μm, 2.1 × 100 mm) on a Shimadzu Nexera X2 LC-30AD system for LC/MS analysis. A 2 μL aliquot of each sample was introduced into the LC autosampler for sequential analysis. The ACQUITY UPLC BEH amide column was maintained with a flow rate of 300 μL/min at 45°C, utilizing a gradient of 5 − 45−60% acetonitrile containing 20 mmol ammonium acetate. Prior to the analysis, each metabolite standard (50 mg/mL) underwent LC-MS/MS to establish optimal MRM transition parameters, facilitating the construction of a metabolite MRM library. Subsequently, the retention time for 40 energy-related metabolites was determined by measuring their corresponding MRM(q1/Q3) transitions one by one. A series of diluted solutions containing reference standards were prepared for LC-MS analysis of 40 energy-related metabolites. Calibration curves were obtained by least-squares fitting based on the peak area of each standard metabolite in the corresponding concentration of the cascade dilution reference standard solution. The MultiQuant software was used to process the raw MRM data files, including peak finding, alignment, extraction, and filtering. Fisher’s exact test was used for enrichment analysis, and FDR correction was used for multiple tests.
Data analysis
Statistical analysis was conducted using GraphPad Prism 8.0 (GraphPad Software). One-way analysis of variance (ANOVA) and Student’s t-test were employed (the t-test is a method that determines whether two populations are statistically different from each other, whereas ANOVA determines whether three or more populations are statistically different from each other). The results were displayed in mean ± standard error of the mean, whereas the multiple measurements of tumor volume growth were displayed in mean ± standard deviation, and *p < 0.05 or **p < 0.01 was considered statistically significant.
RESULTS
oncoVV-AVL enhanced cytotoxicity and apoptosis of GC cells
To assess the cytotoxic effects of oncoVV-AVL on GC, we conducted treatment on two GC cell lines, MKN-45 and NCI-N87, with oncoVV and oncoVV-AVL at MOIs of 0.5, 1, 2, and 5. Our results revealed a gradual decrease in the survival rates of both MKN-45 and NCI-N87 cells following treatment with oncoVV-AVL, as opposed to the oncoVV control (Fig. 1A). Additionally, we observed a marked increase in the apoptosis rate of MKN-45 cells infected with oncoVV-AVL, reaching 34.1%, whereas only 7.6% of cells in the oncoVV control-exhibited apoptosis, indicating that the incorporation of AVL into oncoVV significantly enhances its antitumor efficacy. Consistent with this finding, the apoptosis rates of NCI-N87 cells infected with oncoVV-AVL were found to be eight-fold higher than those treated with oncoVV (Fig. 1B). In summary, our findings elucidate the augmented anticancer activity of oncoVV-AVL in gastric carcinoma.

The antitumor activity of oncoVV-AVL in human GC cells in vitro.
oncoVV-AVL improved viral replication in GC cells
To evaluate the replication capacity of the viruses, we used the TCID50 assay. Our findings demonstrated that the viral yield of oncoVV-AVL escalated from 1.78 × 105 PFU to 6.43 × 106 PFU, whereas the viral copies of oncoVV stagnated at 9.58 × 104 PFU without a significant rise, illustrating that oncoVV-AVL substantially boosted viral propagation in MKN-45 cells (Fig. 2A). Analogous outcomes were observed in NCI-N87 cells (Fig. 2C). To further substantiate these observations, the expression of A27L, a VV membrane protein, was examined via Western blot analysis. The results revealed a marked elevation in A27L expression in both MKN-45 and NCI-N87 cells posttreatment with oncoVV-AVL, as compared with oncoVV (Fig. 2B, D). These results collectively suggest that oncoVV-AVL significantly enhances virus replication within GC cells.
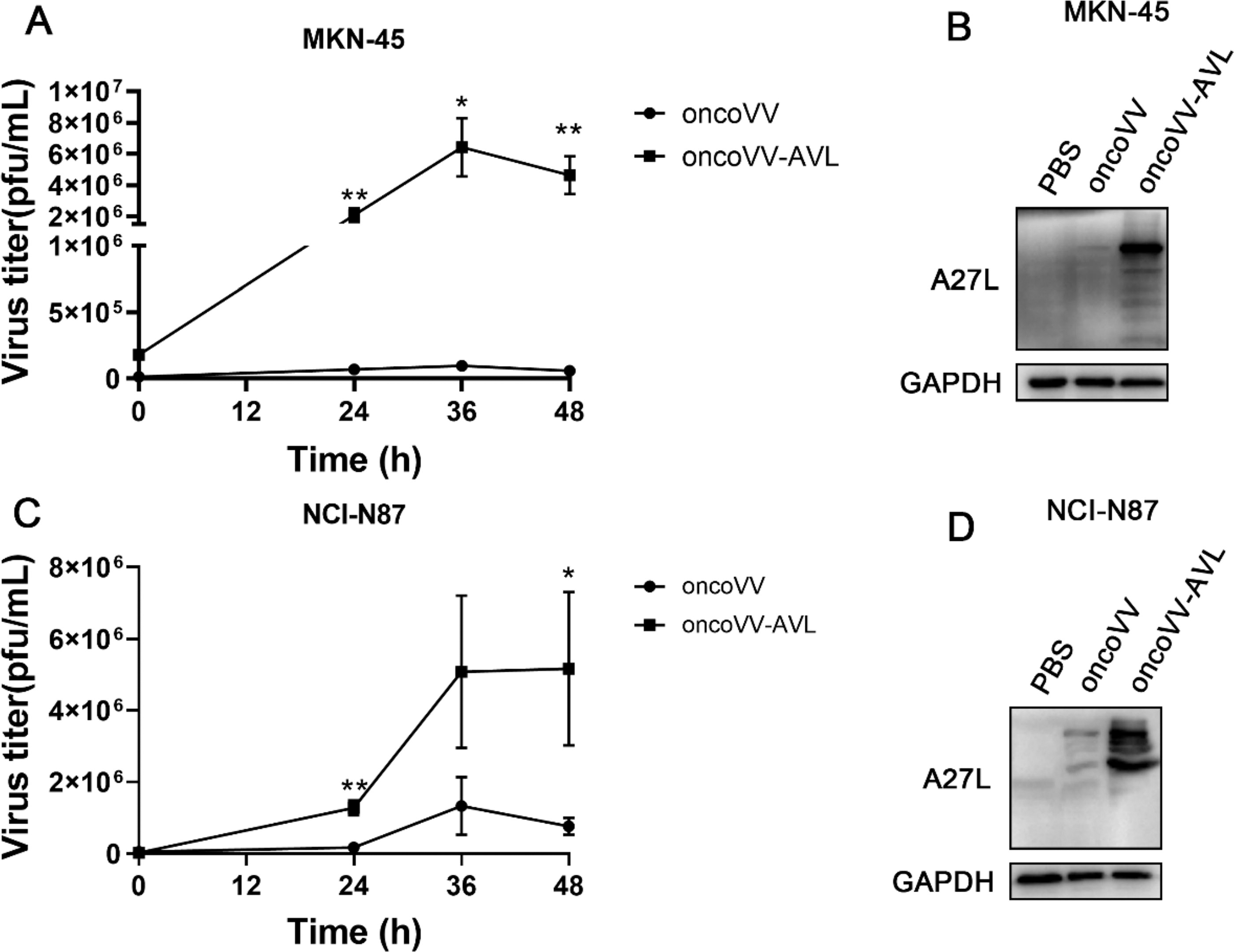
Enhanced viral replication capacity of oncoVV-AVL in GC cells. Viral copies of oncoVV-AVL in MKN-45
oncoVV-AVL regulated signaling pathway
Signaling pathways within tumor cells are frequently dysregulated, serving as critical factors in the genesis and advancement of cancer. Targeting these pathways with specific treatments is a key strategy in cancer therapy. The PI3K/Akt signaling pathway is one of the most frequently deregulated in cancer, and the suppression of its activities has been implied to repress tumor proliferation and angiogenesis. 26,27 The phosphorylation status is used to evaluate the activation of the PI3K/Akt signal pathway. The results indicated that the concentrations of phospho-PI3K and phospho-Akt were markedly reduced in MKN-45 cells following treatment with oncoVV-AVL, as compared with the oncoVV and PBS controls (Fig. 3A). The level of phospho-Akt obviously decreased in NCI-N87 cells treated with oncoVV-AVL compared with controls, whereas the level of phospho-PI3K was unchanged as compared with the oncoVV control, indicating oncoVV-AVL may display antitumor activity through regulating Akt but not PI3K in NCI-N87 (Fig. 3A). The Hippo pathway, a significant player in cancer biology, is predominantly composed of mammalian sterile 20-like protein kinase 1/2 and the YAP. In this pathway, phospho-YAP is predominantly localized in the cytoplasm, where it is subjected to protein degradation, whereas YAP enters in the nucleus, where it forms a complex with the transcription factor TEA domain family member, resulting in promoting cell proliferation and survival genes. 28 The data presented herein indicated that YAP and phospho-YAP undergo protein degradation (Fig. 3B), indicating that oncoVV-AVL exerts its antitumor effects through activating the Hippo pathway.
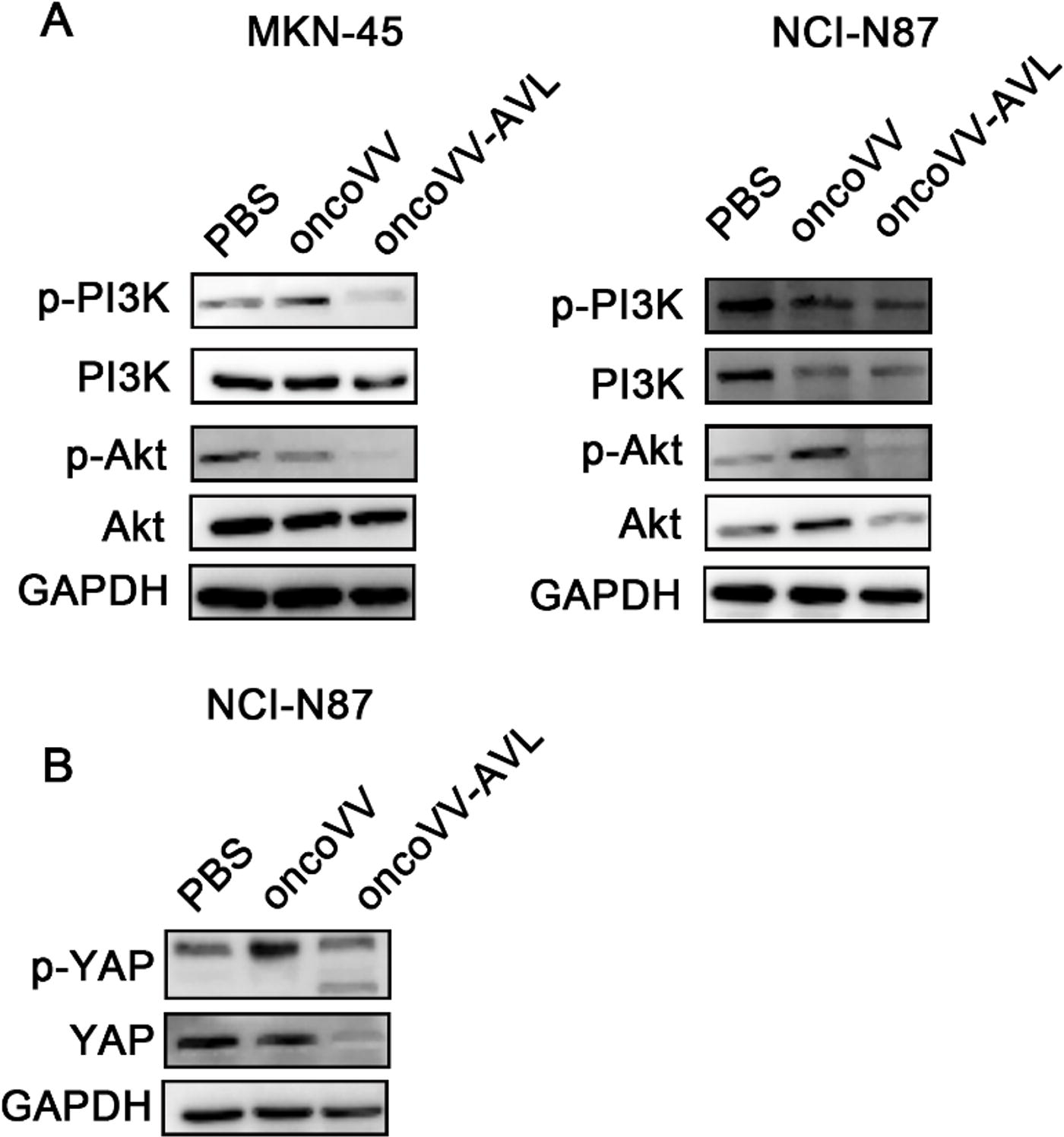
oncoVV-AVL regulates signaling pathways.
oncoVV-AVL increased ROS levels in GC cells
The replication ability of OVs plays a crucial role in oncolytic therapy. The viral reproduction experiments indicated that AVL promotes viral replication in GC cells. Increasing evidence has highlighted that heightened levels of ROS can actually promote viral replication. 29 To assess ROS levels in MKN-45 and NCI-N87 cells, we employed DCFH-DA staining followed by flow cytometry analysis. Our findings indicated that ROS levels were significantly higher at 36 h following inoculation with oncoVV-AVL compared with oncoVV-infected cells under the same MOI in both MKN-45 and NCI-N87 cells (Fig. 4A, B). NRF2 orchestrates the enhancement of gene expression in response to oxidative stress, aiding in neutralization of ROS levels. 30 Elevated NRF2 expression can also promote cancer progression by activating the expression of enzymes within signaling pathways, the products of which are essential for cancer cell growth and proliferation. 31 In order to determine whether oncoVV-AVL downregulates NRF2 protein expression leading to high ROS levels in cells, the expression of NRF2 protein was evaluated in GC cells. The expression of NRF2 in the oncoVV-AVL group displayed a downward trend relative to the PBS and oncoVV groups, indicating that oncoVV-AVL indeed induces elevated ROS levels (Fig. 4C, D). In summary, the data suggest that oncoVV-AVL downregulates NRF2 expression to suppress cancer proliferation and concurrently elevates ROS levels to enhance viral yields.
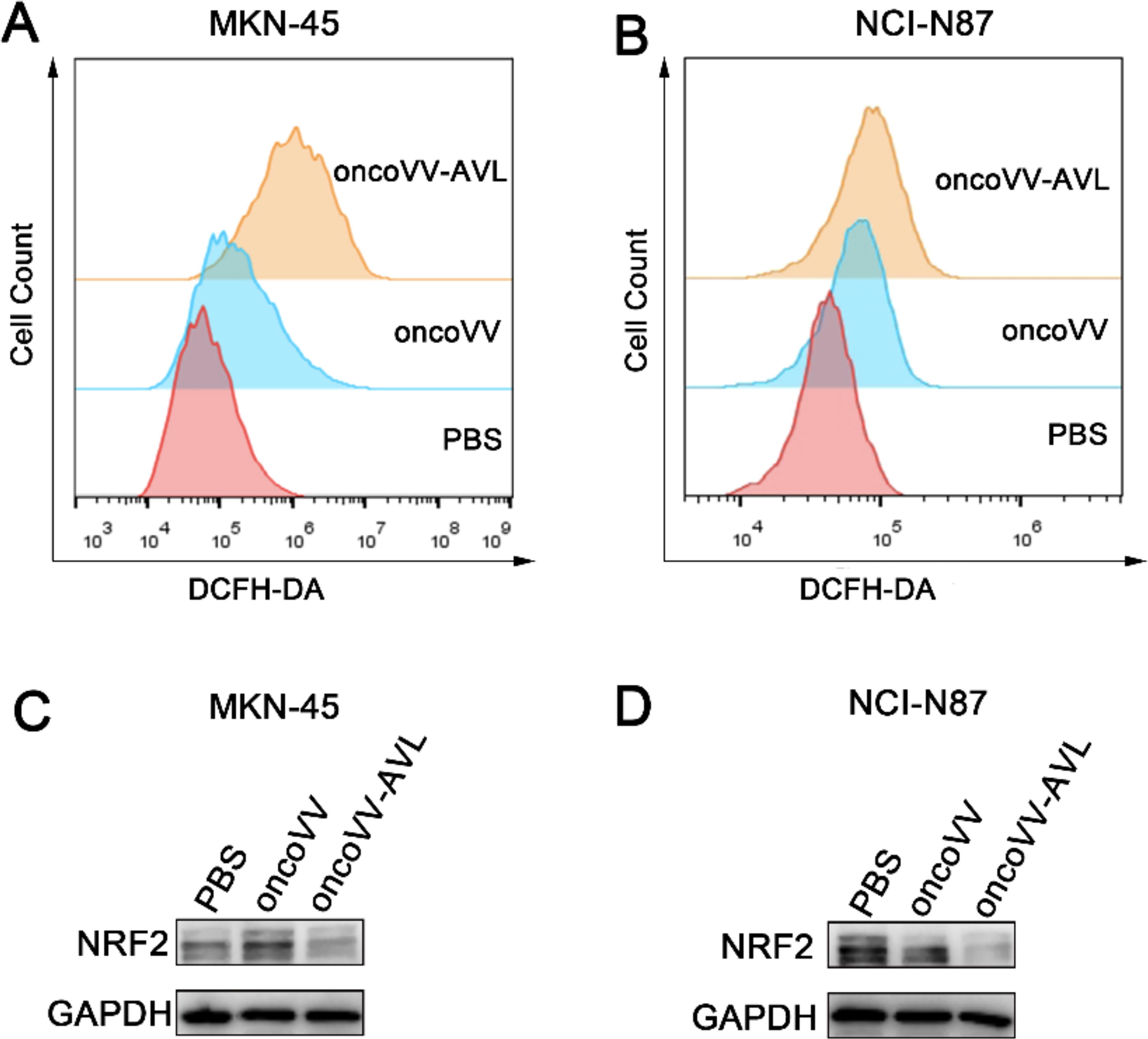
Elevated levels of ROS in GC cells infected with oncoVV-AVL. ROS levels in MKN-45
oncoVV-AVL increased apoptosis and autophagy through enhancing ROS accumulation in GC cells
ROS has been documented as a vital stimulus, not only triggering the initiation of both the apoptotic and autophagy pathways but also facilitating viral reproduction. 28 To further ascertain whether ROS accumulation indeed influences viral reproduction, we first assessed the impact of ROS levels on viral replication. NADPH is essential for preserving the intracellular redox balance and functions as a scavenger for ROS. In the presence of NADPH, the expression levels of A27L were diminished in oncoVV-AVL treatments in GC cells, indicating that heightened ROS levels indeed support viral replication (Fig. 5A). Furthermore, we explored whether virus-induced ROS accumulation leads to apoptosis (Fig. 5B). As illustrated in Fig. 5B, the results revealed that the apoptosis rates in MKN-45 cells for the PBS, oncoVV, and oncoVV-AVL groups were 2.88%, 5.33%, and 57%, respectively. However, upon the addition of NADPH (100 μM), the apoptosis rates for the NADPH, oncoVV+NADPH, and oncoVV-AVL+NADPH groups were 3.48%, 3.93%, and 36.3%, respectively. Notably, there was no significant variation in apoptosis rates between the PBS and PBS+NADPH groups, nor between the oncoVV and oncoVV+NADPH groups. Conversely, the apoptosis rate in the oncoVV-AVL+NADPH group was notably lower in NCI-N87 cells, compared to the oncoVV-AVL group. Collectively, these findings demonstrate that oncoVV-AVL-induced ROS accumulation promotes apoptosis in GC cells. In the presence of NADPH, the oncoVV-AVL group exhibited a reduction in autophagy within MKN-45 cells, whereas only minimal alterations were observed in the PBS and oncoVV treatments, suggesting that oncoVV-AVL infection triggers ROS accumulation, leading to enhanced autophagy (Fig. 5C). Consistent with this observation, we noted a diminished LC3II/I ratio in NCI-N87 cells treated with oncoVV-AVL combined with NADPH, compared with the oncoVV and PBS controls (Fig. 5D).
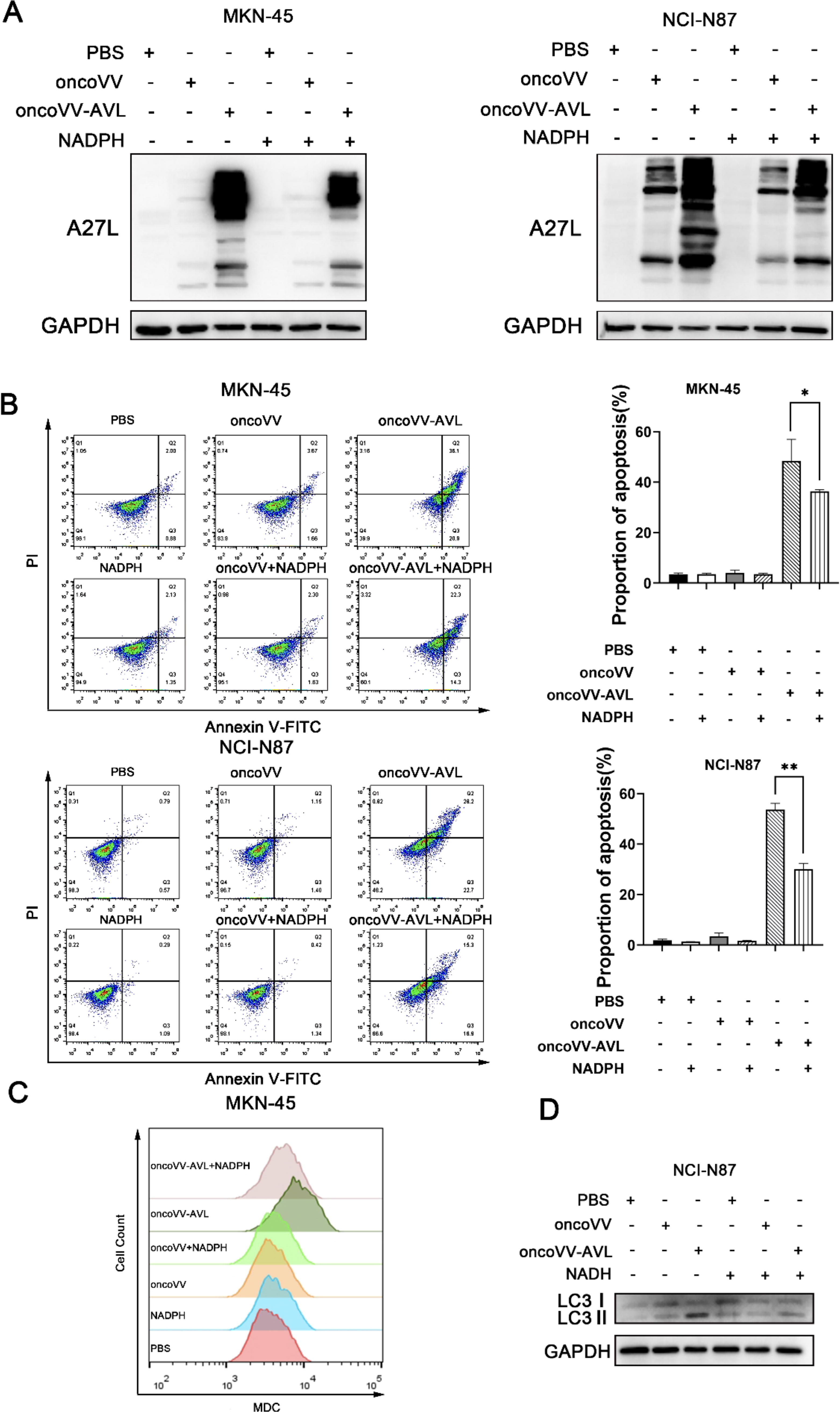
Viral replication, cell apoptosis, and autophagy are influenced by elevated levels of ROS in GC cells infected with oncoVV-AVL.
oncoVV-AVL reprogrammed GC cell metabolism to promote viral replication
ROS are generated as by-products during various cellular processes, including cellular respiration in the mitochondria and metabolic processes. 32 To ascertain that oncoVV-AVL enhances ROS accumulation through modulating metabolism, a metabolomic analysis was performed. The targeted metabolomic analysis of MKN-45 cells treated with oncoVV-AVL revealed significant reprogramming of cellular metabolism, which is indicative of the viral strategy to manipulate host resources for its propagation and survival. The observed changes in metabolite levels, as illustrated in Fig. 6A, indicate a profound interference with host cell metabolism by oncoVV-AVL. The reduction of NADH, fumarate, pyruvate, and UDP glucose (Fig. 6B) suggests a disruption to both glycolysis and the tricarboxylic acid (TCA) cycle—fundamental metabolic pathways. The decline in ribulose-1,5 bisphosphate, a key component in both the Calvin cycle of photosynthesis and the pentose phosphate pathway, could result in a decrease in NADPH production. In contrast, a marked increase in adenosine diphosphate glucose (ADPG) suggests a reallocation of metabolic substrates toward viral replication, as ADPG is a precursor in glycogen synthesis and could potentially be utilized by the virus for its own needs. These metabolic pathways are vital for cellular energy generation and biosynthesis, and their disruption can curtail the supply of critical metabolites required for normal cellular functions. Collectively, the data imply that oncoVV-AVL impairs cancer cells by sabotaging key metabolic pathways, which not only hampers the host cell’s viability but may also unwittingly assist the virus in its replication.
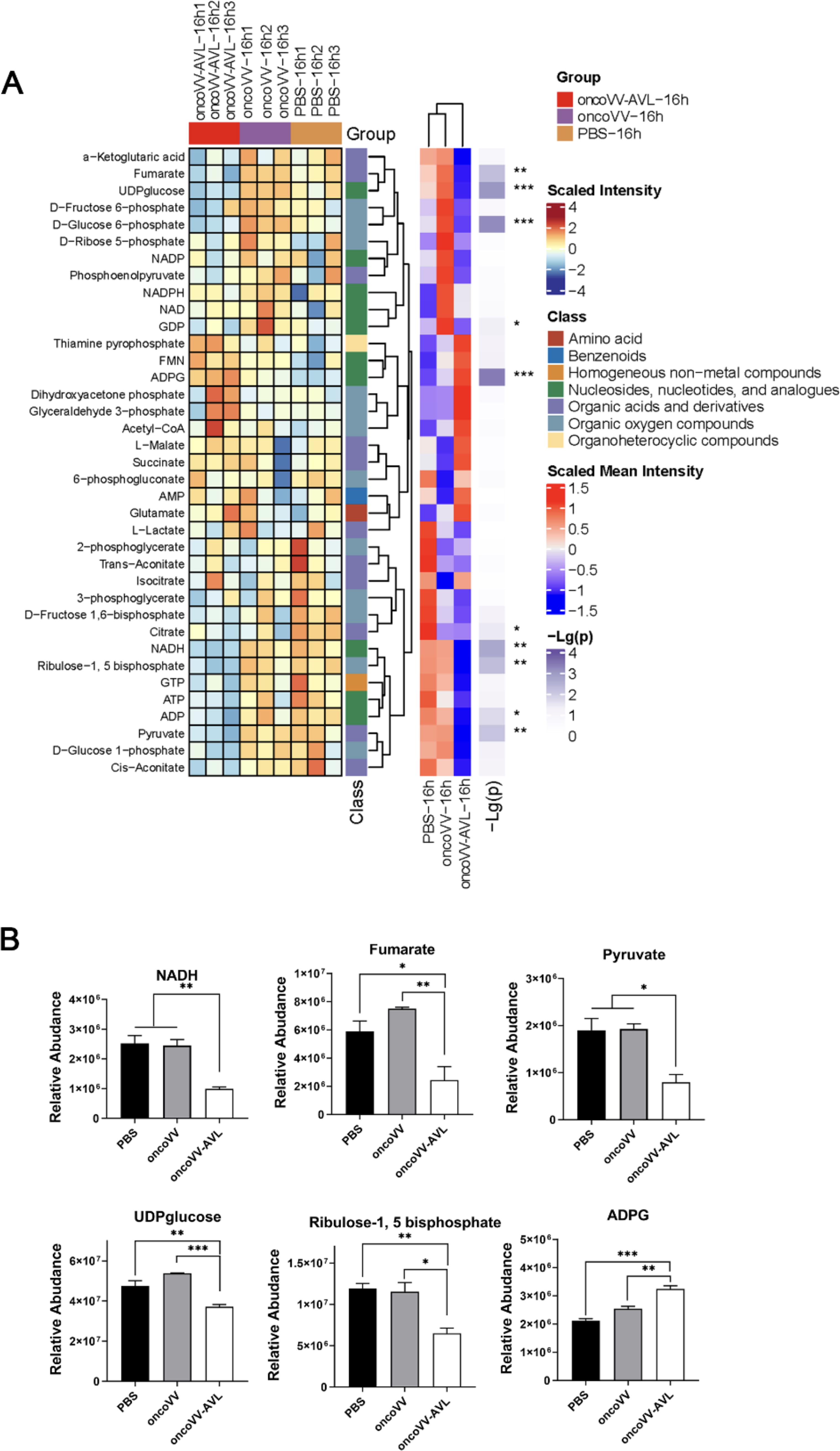
Cellular metabolism modified by oncoVV-AVL.
oncoVV-AVL repressed tumor growth in vivo
To evaluate the efficacy of oncoVV-AVL in antitumor in vivo, a gastric xenograft model was established using 6-week-old female nude mice, each implanted with 3 × 106 MKN-45 cells. The tumor-bearing mice were divided into three groups based on average tumor volume, with seven mice in each group. Prior to treatment, the average tumor volumes were 190.16 mm3, 190.83 mm3, and 190.96 mm3, respectively. After intratumoral injection and up to 30 days (before sacrifice), all three groups of mice survived. The average tumor volumes for the 0.9% NaCl, oncoVV, and oncoVV-AVL groups were 834.37 mm3, 945.29 mm3, and 395.62 mm3, respectively (Fig. 7A). The analysis of the tumor growth curve indicated that the average tumor volume following intratumoral injection of oncoVV-AVL was significantly lower than in both the 0.9% NaCl and oncoVV groups, demonstrating the ability of oncoVV-AVL to effectively restrain the growth of MKN-45 xenograft tumors. Tumor tissues harvested from the mice were then subjected to Western blot analysis to assess A27L protein expression. The oncoVV-AVL group showed a marked increase in A27L levels, signifying robust replication of oncoVV-AVL within the tumor tissue (Fig. 7B). H&E staining of the tumor tissue (Fig. 7C) showed that the cellular nuclei in the 0.9% NaCl group were densely arranged with no discernible changes in cell morphology. Conversely, the oncoVV treatment group showed slight alterations in nuclear structures and the presence of cell cavities. Remarkably, the AVL group exhibited extensive tumor cell necrosis, characterized by the presence of cytoplasmic vacuoles in most cells, suggesting that oncoVV-AVL administration triggers tumor tissue necrosis, thereby impeding tumor growth. IHC analysis (Fig. 7D) further elucidated that the intensity of brown staining in the tumor tissue correlated with the level of OV replication, with the oncoVV-AVL treatment exhibiting more pronounced staining than the oncoVV and 0.9% NaCl controls, suggesting that oncoVV-AVL can strengthen the antitumor effect by augmenting viral reproduction in GC cell MKN-45.
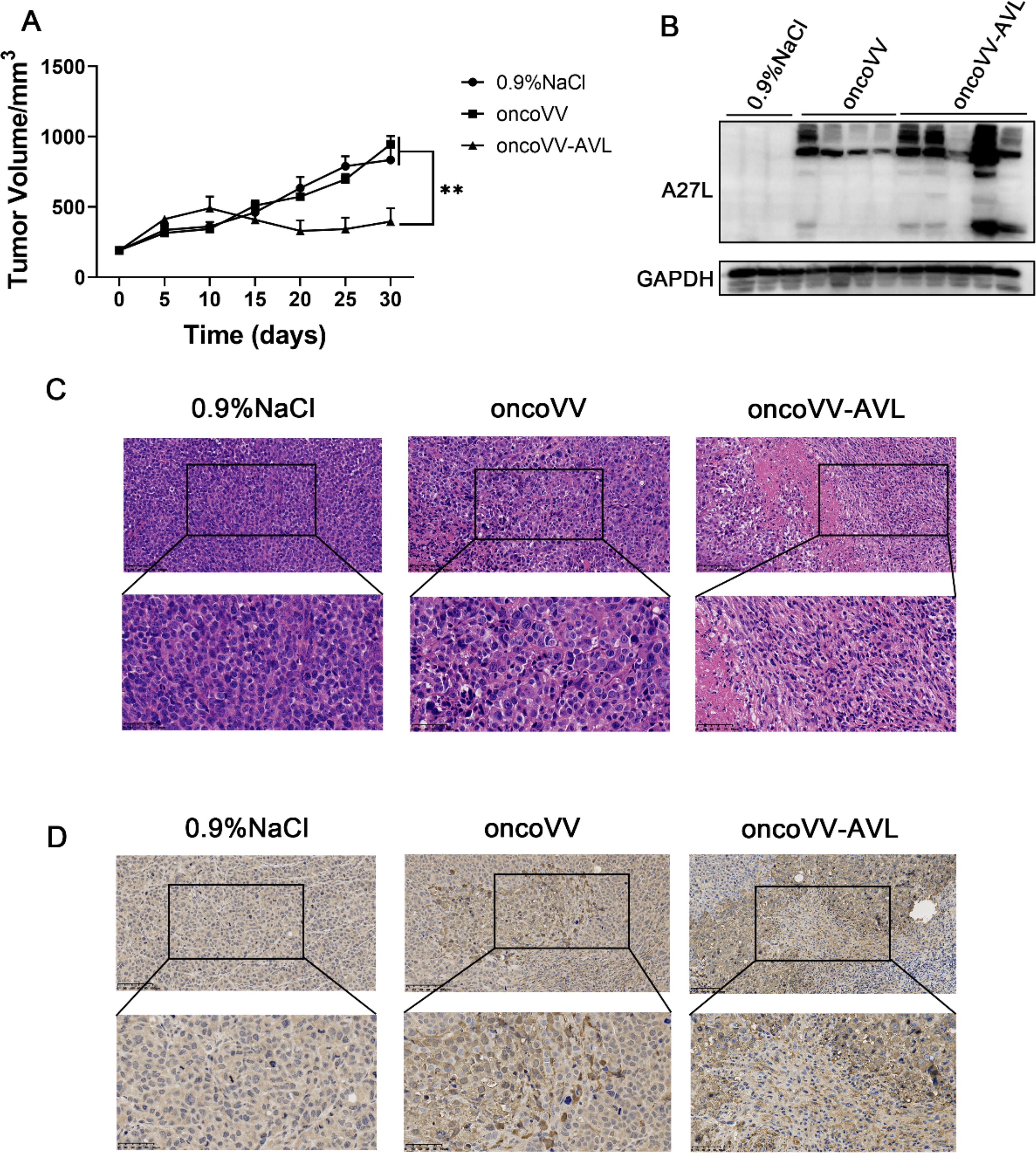
Tumor growth is repressed by oncoVV-AVL in vivo.
DISCUSSION
Currently, the clinical treatment of GC has not seen significant improvement. OVs have been designed for utilization in various types of cancer. Nevertheless, there is a scarcity of reports concerning the prospective application of OVs in GC. For instance, due to the limited expression of the coxsackie-adenovirus receptor, the infectivity of adenovirus in gastrointestinal cancer cells is generally poor. In this study, a robust anti-GC effect of recombined oncolytic VV was demonstrated both in vivo and in vitro.
OV therapy primarily attains its objective through two distinct pathways: first, by selectively infecting cancer cells and replicating within them to induce direct lysis, and second, by activating the immune system to eliminate the tumor cells. Increasing evidence has solidified that OVs can reprogram metabolic pathways to harness the metabolic byproducts generated by tumor cells, thereby supporting their own replication.
33
By inhibiting glycolytic enzymes, cancer cells are prevented from acquiring the ATP necessary for cell proliferation, resulting in a reduction of the glycolytic rate in these cells. This reduction is advantageous for OVs, as it allows them to utilize glycolytic by-products for their replication and to induce cell apoptosis.
34
Vorobyev et al. demonstrated that 2-
In GC, it has been found that targeting ROS through autophagy elicits an antitumor effect. 37 It is well-established that ROS are by-products of cellular metabolism, predominantly generated in mitochondria. Notably, viruses have been found to induce mitochondrial ROS production to facilitate their replication within cells. 29 In line with that, our finding indicated that oncoVV-AVL exhibited a significantly reduced replication capability within GC cells in the presence of NADPH, suggesting that high levels of ROS promote viral replication. Moreover, ribulose-1,5-bisphosphate is involved in the Calvin cycle of photosynthesis and the pentose phosphate pathway, with the latter being closely related to the production of NADPH within cells, which is crucial for the cellar antioxidant capacity. 38 In the current study, metabolomic analysis shows that the production of ribulose-1,5-bisphosphate is significantly decreased; we inferred that oncoVV-AVL reduces the antioxidant capacity of GC cells to promote viral reproduction. Furthermore, Sahoo et al. have proven that Zika virus can promote its replication by downregulating the expression of NRF2 protein, affecting cellular metabolism. 39 Correspondingly, the current research demonstrates that the expression of NRF2 protein is downregulated after GC cells are infected with oncoVV-AVL, indicating that oncoVV-AVL also promotes its replication by downregulating the expression of NRF2 protein. Additionally, consistent activation of NRF2 is linked to cancer progression, with overexpression of NRF2 observed in various cancers including lung, glioblastoma, pancreas, ovarian, breast, and colorectal. 40,41 In line with this, we infer that oncoVV-AVL downregulates NRF2 expression to suppress cancer proliferation. Overall, our investigation reveals that oncoVV-AVL significantly elevates ROS accumulation and suppresses NRF2 expression, thereby facilitating viral replication and lysis of GC cells.
Traditional treatments for GC have demonstrated a poor response to existing chemotherapy regimens, highlighting the urgent need for innovative therapeutic strategies. In our current research, we have found that the oncolytic VV, oncoVV-AVL, exhibits significant antitumor efficacy both in vitro and in a xenograft tumor model. These findings suggest that oncoVV-AVL may be a promising candidate for clinical trials, with further considerations required regarding its safety, feasibility, and immune-activating effects. To enhance the antitumor efficacy of oncolytic virotherapy, investigators often arm the viruses with genes such as immune checkpoint inhibitors, tumor-associated antigens, cytokines, and bispecific T cell engagers. These modifications aim to promote viral replication and boost the immune response, thereby augmenting the overall therapeutic impact. 42 Although some cytokines, such as tumor necrosis factor, interleukins, and interferons, have been used to engineer OVs, they have not been effectively applied in clinical trials, because they may elicit direct antiviral effects. 43 –45 Unlike previous strategies, we used marine lectins to arm the OV, oncoVV-AVL, which can reprogram the metabolic pathways to increase ROS levels and downregulate NRF2 expression, leading to amplified viral replication in cells. These research results broaden the scope of oncoVV-AVL application in cancer therapy.
Footnotes
AUTHORS’ CONTRIBUTIONS
T.Y. and G.L. were responsible for the conception and design of the study; B.Z., Z.H., J.Z., and J.Y. conducted the experiments; K.C. and Y.Z. were in charge of data analysis; and T.Y. prepared the article. Each author has reviewed and approved the final version of the article.
DATA AVAILABILITY STATEMENT
All primary and processed data that substantiate the conclusions of this research are accessible upon request.
AUTHOR DISCLOSURE
The authors declare no conflict of interest.
FUNDING INFORMATION
This research was funded by