
Abstract
Mucopolysaccharidosis type II (MPS II) is an X-linked lysosomal storage disorder caused by pathogenic variants in the IDS gene encoding iduronate-2-sulfatase (IDS), which hydrolyzes sulfate groups in dermatan sulfate and heparan sulfate. The current treatment for MPS II includes enzyme replacement therapy and hematopoietic stem cell transplantation (HSCT). Both therapies have shown limited penetration through the blood-brain barrier. Anecdotal cases have been reported with the HSCT benefit to treat neurological problems in MPS II. Herein, we generated an MPS II mouse model using CRISPR/Cas9 to examine the effectiveness of CNS-directed, adeno-associated virus (AAV)2/9-mediated human IDS gene transfer in expressing sustained IDS and improving behavior performance in this model. The intracerebroventricular administration of AAV2/9-hIDS showed higher IDS activity in the central nervous system and better auditory function compared with those by intravenous administration. The results provide a strong proof of concept for the clinical translation of our approach to treating patients with MPS II and cognitive impairment.
INTRODUCTION
Mucopolysaccharidosis type II (MPS II), also known as Hunter syndrome, is an X-linked recessive lysosomal storage disease caused by a deficiency of iduronate-2-sulfatase (IDS). It is the most common type of MPS in China and other East Asian regions, accounting for approximately half of forms of MPSs. First described by Dr. Hunter in 1917, 1 the deficiency of IDS leads to pathological accumulation of dermatan sulfate (DS) and heparan sulfate (HS) within tissues, 2 resulting in a spectrum of symptoms. These symptoms include coarse facial features, skeletal deformities and joint stiffness, growth retardation with short stature, hepatosplenomegaly, upper airway obstruction, recurrent ear infections, and cardiac valve abnormalities. Hearing loss is a prominent disease feature. 3 –6 The neuronopathic MPS II clinical form, with significant central nervous system (CNS) involvement, accounts for approximately 60%, characterized by normal development in children before 18–24 months, after which global neurodevelopmental arrest, delay, and subsequent neuroregression are noticeable. 7
Current treatments for MPS II include enzyme replacement therapy (ERT) and hematopoietic stem cell transplantation (HSCT). The ERT replaces the deficient or absent enzyme with a functional recombinant IDS administered intravenously (IV) weekly. 8 The ERT reduces the urinary glycosaminoglycans (GAGs) levels and liver/spleen volume and increases musculoskeletal mobility in patients with MPS II. The inability to cross the blood-brain barrier (BBB) impedes ERT from being efficacious in treating the neurological manifestations of MPS II. In addition, the ERT is associated with significant anti-drug antibody formation. Another critical limitation of ERT lies in the disease burden it places on families as patients require lifelong IV weekly infusions. Early HSCT can ameliorate MPS II symptoms, but limited donor availability and graft-versus-host disease remain a concern. 9,10 Thus, addressing how to improve neurological symptoms in patients with MPS II is urgently needed.
Previous reports of improving neurological symptoms in MPS II mice have been through IV-ERT administration. Sonoda et al. developed a BBB-penetrating fusion protein, JR-141, composed of an anti-human transferrin receptor antibody and full-length hIDS. IV administration of JR-141 enabled entry into the brain parenchyma of cynomolgus monkeys and mice, reducing GAG levels in the brain, although no naked IDS enzyme was detected in the brain. 11 In subsequent clinical trials, JR-141(Pabinafusp Alfa) significantly reduced HS levels in the cerebrospinal fluid of patients, with most showing either improved or stabilized cognitive function. 12,13
Gene therapy aims to restore gene function by providing exogenous genes. A significant advantage of gene therapy is that it can provide long-term, sustained treatment without constant repeat administration. Gene therapy can be delivered directly to the CNS to treat the neurological symptoms of MPS II. Various viral vectors have been widely used in preclinical and clinical gene therapy trials for different diseases, among which adeno-associated virus (AAV) is the most used viral vector. AAV9 has been widely used in therapeutic studies of neurological disorders because of its unique pro-neurological properties. 14,15 Recently there have been significant advances in both clinical and preclinical development of genetic therapies for MPS II. Smith et al. investigated the efficacy of different doses of AAV9.CB7.hIDS (RGX-121) through IT and IV administration. The study showed that a dose of 1 × 1010 vector copies (VC) IT/IV route was sufficient to normalize metabolic and skeletal outcomes, while at least 1 × 1011 VC IT/IV route was required to achieve neurological improvement. 16 The corresponding clinical trials (NCT03566043, NCT04571970; https://clinicaltrials.gov/) are ongoing. Laoharawee et al. demonstrated that intravenous administration of 1.51 × 1012 VC rAAV9-hIDS resulted in a reduction of GAG levels of the brain, although IDS enzyme activity reached only 4–50% of wild-type (WT) levels. 17 Additionally, Rigon et al. designed a tailored nanomedicine approach based on brain-targeted polymeric nanoparticles (g7-NPs). Following weekly intravenous administration, significant reductions in GAG deposits were observed in the liver and brain parenchyma of MPS II mice after 6 weeks, accompanied by decreases in neuroinflammatory biomarkers (GFAP, CD68, and LAMP-2). 18 Gene editing has also been applied in the treatment of MPS II. AAV8 with albumin locus-targeting zinc finger nucleases were administered IV to MPS II mice. The treatment resulted in elevated IDS enzyme activity in the blood and peripheral tissues, as well as a decrease in GAG accumulation in visceral organs and the brain. 19
In this study, we delivered AAV2/9 carrying the human IDS gene into newly constructed MPS II mice by intracerebroventricular (ICV) injection and compared its efficacy with that of other routes of administration (e.g., IV). The results showed that ICV and IV routes of administration were efficacious in improving the CNS and peripheral symptoms of MPS II mice. Still, ICV was more effective in increasing the activity of IDS enzymes in the CNS.
MATERIALS AND METHODS
Animal care and husbandry
The experimental animals were C57BL/6J mice, and their breeding and maintenance were conducted at the SPF Experimental Animal Center of Xinhua Hospital. All mice were housed under controlled conditions at 21–22°C with a 12-hour light–dark cycle. They had ad libitum access to standard rodent chow and autoclaved acidified sterile water (pH = 3.0). The animal experiments and procedures adhered to the Ethics Committee for Animal Welfare regulations of Xinhua Hospital, Affiliated with Shanghai Jiao Tong University School of Medicine. We performed injection of guide RNA (gRNA) targeting the IDS gene, donor oligonucleotides containing the p.R513Q (CGG to CAG) mutation, and into fertilized mouse eggs to generate offspring with point mutation. Mouse toes were collected for DNA extraction and genotyped using PCR (F1: 5′-GAGGACCTCGTAGAACTTGTATCTC-3′, R1: 5′-AGACCTCCATGTTGGGAGTCATTA-3′). Male MPS II mice were selected. Body weight (g) and body length (cm) (the distance from the tip of the mouse’s nose to its anus) measurements were taken on days 7, 14, 21, 28, 42, 56, 70, and 84 after the birth of the mice.
AAV vector design and production
The human IDS (hIDS) expression cassette encoding IDS was cloned between the modified inverted terminal repeats (ITRs) of AAV2 as the AAV-GOI plasmid as described previously 20 for following AAV production. The 1,653 bp wild-type human IDS coding sequence (NM_000202.8) was synthesized by Genscript and cloned between the chicken b-actin hybrid (CBh) promoter and an exogenous polyA sequence using the SacII and NheI restriction enzymes. The construct was verified by Sanger sequencing. The total genome length of this constructed ITR-CBh-hIDS-polyA cassette is approximately 3,000 bp (Fig. 1). Recombinant AAV2/9-hIDS vectors were produced by transient triple-transfection of HEK293T cells with RC plasmid encoding the AAV2 Rep protein and AAV9 Capsid, adenoviral helper plasmid encoding the E2, E4, and VA, and the constructed AAV-GOI plasmid containing the hIDS expression cassette, which has been described previously. 20 AAV particles were purified using iodixanol-based density gradient ultracentrifugation. rAAV was titrated by probe-based ddPCR (Biorad) assay and characterized by silver staining.

Adeno-associated virus (AAV) vector constructs for hIDS and expression. hIDS were transcriptionally regulated by the CBh promoter and an exogenous polyA sequence, flanked with AAV2-ITRs on both 3′ and 5′ ends. CBh, chicken b-actin hybrid; ITR, inverted terminal repeat.
AAV vector administration
Genotyped MPS II mice were administered total AAV2/9-hIDS vector doses of 1 × 1011 VC at 6–8 weeks of age. The IV injections were given in a 200 μL volume through the lateral tail vein. As previously described, the ICV injections were given to animals in a 5 μL volume. 21 Briefly, animals were injected intraperitoneally with a ketamine/xylazine mixture (100 mg/kg ketamine, 10 mg/kg xylazine) to produce deep anesthesia and then mounted in a stereotactic frame (RWD 68018) (stereotactic coordinates AP −1.5, ML −1.1, DV −4.5 mm from bregma). An incision was made to expose the cranium, a small hole was drilled as a site for the injection, and then a Hamilton syringe (Model 701) was used to carry out the infusion at approximately 1μL/min rate by hand. The syringe was left in place for an additional 8 min. The scalp was sutured after completion of the injection, and after recovery from the anesthesia, the mouse was returned to standard housing.
Sample collection and preparation
A dry blood spot was obtained via the tail vein, and blood was collected by eye extraction and collected into ethylenediaminetetraacetic acid tubes (two tubes) and centrifuged at 6,300 rpm for 3 min (Thermo Scientific Sorvall Legend Micro 17R). One tube was used to extract leukocytes and stored at −20 to −80°C for leukocyte IDS assay. The other tube was used for biochemical analysis. Urine was collected and stored at −80°C until used for creatinine and GAG assay. Organs were harvested by first determining animal weight before euthanasia. The animals were perfused with 60 mL of 1× phosphate-buffered saline (PBS). The heart, lung, liver, spleen, kidney, and brain were harvested and weighed. The brain was micro-dissected into the cerebellum, cortex, hippocampus, hypothalamus, and thalamus/brainstem. The tissues were immediately snap-frozen and stored at −80°C until further processing.
For tissue processing, the cerebellum, cortex, hippocampus, hypothalamus, and thalamus/brainstem were added into preassigned 2.0 mL locked-cap microtubes (Eppendorf) containing 0.5 mm glass beads in 1 mL of ddH2O. The 50 mg of the heart, liver, spleen, lung, and kidney were added into assigned tubes containing 0.5 mm glass beads in 1 mL of ddH2O. All prepared samples in the bead tubes were then homogenized to generate tissue homogenate. Fifty microliters of tissue homogenates were transferred into 1.5 mL microtubes (Eppendorf) and stored at −20 to −80°C for Quantitative Polymerase Chain Reaction (qPCR). The remaining tissue homogenates were clarified at 13,000 rpm for 15 min at 4°C (Thermo Scientific Sorvall Legend Micro 17R). All of the supernatants (tissue lysates) were transferred into new microtubes and stored at −20 to −80°C until used for IDS, GAGs, and protein assays.
For leukocyte extraction, we added 1 mL Red Cell Lysis Buffer to the EDTA tube at room temperature, standing for 10 min, centrifuged at 2,000 rpm for 10 min (Thermo Scientific Sorvall Legend Micro 17R), and aspirated the supernatant. The above steps were repeated three times. Afterward, 800 μL 0.85% normal saline was added and centrifuged at 6,300 rpm for 3 min (Thermo Scientific Sorvall Legend Micro 17R). The supernatant was discarded, and 200 μL of ddH2O were added. The leukocyte homogenates were produced by ultrasonication.
IDS enzyme activity
The IDS assay was carried out as previously described.
22
IDS enzyme activity was measured using the substrate of 4-methylumbelliferyl-a-
GAG assay
The volume of 20 μL from each sample was transferred into a 1.8 mL mass spectrometry sample bottle. We added 20 μL of high-purity methanol-d4 (Sigma,151947), vortex-mixed it, and blew nitrogen at 30°C. Then, 10 μL of 2,2-dimethoxypropane (Sigma, D136808) and an additional 200 μL of 3N hydrochloric acid (Sigma, 90964) were added to the dried sample bottle. We quickly sealed the sample bottle and vortex-mixed it. We placed the sample in a 65°C incubator for 75 min. After completion of the reaction, the vial was removed and cooled to room temperature, and then the nitrogen was at 30°C. We added 500 μL of a mixed isotopic internal standard aqueous solution to the dried sample, vortex-mixed, filtered, and proceeded to machine injection analysis (AB Sciex Triple Quad 4500). We prepared standard solutions of chondroitin sulfate (CS) (Sigma, C6737)/HS (Sigma, H7640) at concentrations of 250/500, 100/200, 25/50, 10/20, 2.5/5, 1/2, 0.5/1 and 0.25/0.5 μg/mL and DS (Sigma, 330714W) at concentrations of 500, 200, 50, 20, 5, 2, 1, and 0.5 μg/mL. We used the detection values of the eight concentrations of standard solutions to plot the standard curves and then calculated the concentrations of CS, HS, and DS. The tissue GAG was reported as μg GAG/mg protein, and the urine GAG was reported as mg GAG/mmol creatinine. CS, DS, and HS are three types of GAGs. The LOD of CS, DS, and HS assay was 0.25, 0.5, and 0.5 μg/mL, respectively.
qPCR for IDS vector sequences
DNA was isolated from the tissue homogenates by FastPure Cell/Tissue DNA Isolation Mini Kit (Vazyme, DC102-01). Reaction mixtures of 10 μL contained 1uL cDNA, 5 μL of 2× ChamQ Universal SYBR qPCR Master Mix (Vazyme, Q711), 200 nM each of forward and reverse primers, and 3.6 μL ddH2O. A C1000 Touch Thermo Cycler (Bio-Rad) equipped with CFX manager software v3.1 was used for qPCR reaction. The qPCR conditions were 95°C for 30 s, followed by 40 cycles of 95°C for 10 s and 60°C for 30 s. IDS primers used were: forward primer: 5′-TGTGCCCAGTACATGACCTT-3′; IDS reverse primer: 5′GGAGAGTGAAGCAGAACGTG-3′. A standard curve was generated by serial dilution of the plasmid. ddH2O was used as a negative control. qPCR results from the analysis were expressed as VC per genome equivalent.
Histopathology
A graded series of alcohols processed formalin-fixed tissues. For hematoxylin and eosin(H&E) analysis, tissues were embedded in paraffin, and 4-mm sections were cut and stained. For (alcian blue) AB staining, 4-mm sections were cut, deparaffinized, and hydrated to distilled water. Sections were immersed in a 3% acetic acid solution for 3 min and stained with AB for 30 min. After rinsing with running tap and distilled water, the slides were counterstained with nuclear fast red.
Auditory brainstem responses
The auditory thresholds were assessed for each group at 6 months of age. The experiment utilized short sounds (click) and short pure tones at 4k, 8k, 11k, 16k, 22k, and 32 kHz frequencies as stimuli (1 ms rise/fall time, 4 ms plateau). The stimuli were bandpass-filtered within the range of 300–3,000 Hz, with 1,024 repetitions and a scanning time of 10 ms. The intensity started from the maximum stimulus intensity (90 dB SPL) and gradually decreased in 10 dB steps until no repeatable auditory brainstem responses (ABR) waveform was detected. Then, it was increased by 5 dB until repeatable ABR waveforms were detected, determining the threshold as the lowest stimulus intensity at which a repeatable ABR wave III could be discerned.
Modified Morris water maze
A pool measuring 1,200 mm in diameter was filled 12 cm deep with water (23 ± 2°C) mixed with non-toxic white acrylic paint. Three-dimensional, multi-colored visual cues were attached to the walls of the pool according to the cardinal directions (North, South, East, and West), and a platform was placed at the water surface in the northwest quadrant of the maze (l day with the platform visible and 4 days with the platform hidden). Age-matched, 6-month-old mice were released in the maze facing the southern wall of the pool. The mice were given 60 s to explore the maze, and the total time spent swimming before locating the submerged platform was recorded. If a mouse did not reach the platform within 60 s, the animal was guided to the platform and given 20 s to explore. Animals were given four trials daily for 5 days, with 15 min between each trial. On day 6, spatial exploration testing was conducted with the platform removed. The exploration time was extended to 120 s, and the number of times each mouse crossed through the original platform area was recorded. 23
Y-maze spontaneous alternation
The Y-maze assessed spatial learning and working memory. 24 Animals were placed in a testing room and allowed to habituate for 15–30 min in low light. The Y-maze apparatus consists of three identical arms (zones), each with a distinct visual cue. The mouse was placed in one of the arms and allowed to explore the maze freely for a single 8-min trial. Zone choices were recorded sequentially. Spontaneous alternation was determined when the animal explored an overlapping triplet of three distinct zones. Percent spontaneous alternation was scored as (of alternations/[total zone entries − 2]) × 100.
Mouse skeletal imaging
Skeletal evaluations were conducted using plain radiography. Animals were radiographed under tribromoethanol anesthesia (240 mg/kg) with an INSIGHT VET DXA X-ray imaging system, operating at 3.0 mAs and 60 kVp, with a source-to-image distance of 55 cm. Images were analyzed and processed using a DICOM Viewer. Skeletal anatomical structures were subjected to micro-CT scanning using a CT scanner (NMC-200). The scans were performed in continuous scanning mode at medium magnification with standard X-ray tube settings and exposure parameters (90 kV, 0.04 mA, 200 ms). Raw images were reconstructed using the 3D reconstruction software Recon. Data analysis software Avatar was used to analyze the target region of interest, ensuring the same area was selected for analysis across all samples.
Statistical analysis
GraphPad Prism 8.0 was used for all plotting and statistical analyses. Error bars on the graphs represented either group mean ± standard deviation or standard error of the mean. Morris water maze data were analyzed for multiple group comparisons using a two-way analysis of variance (ANOVA). The Mann-Whitney U test was employed for non-parametric comparison analysis of Y-maze spontaneous alternation data. For IDS activity, GAG levels, and vector biodistribution, two-way ANOVA followed by Tukey’s multiple comparisons test was used. Biochemical and radiological data were grouped by mouse treatment type and evaluated using one-way ANOVA. A p-value of <0.05 was considered statistically significant for all tests.
RESULTS
Disease-relevant phenotypes in a new MPS II mouse model
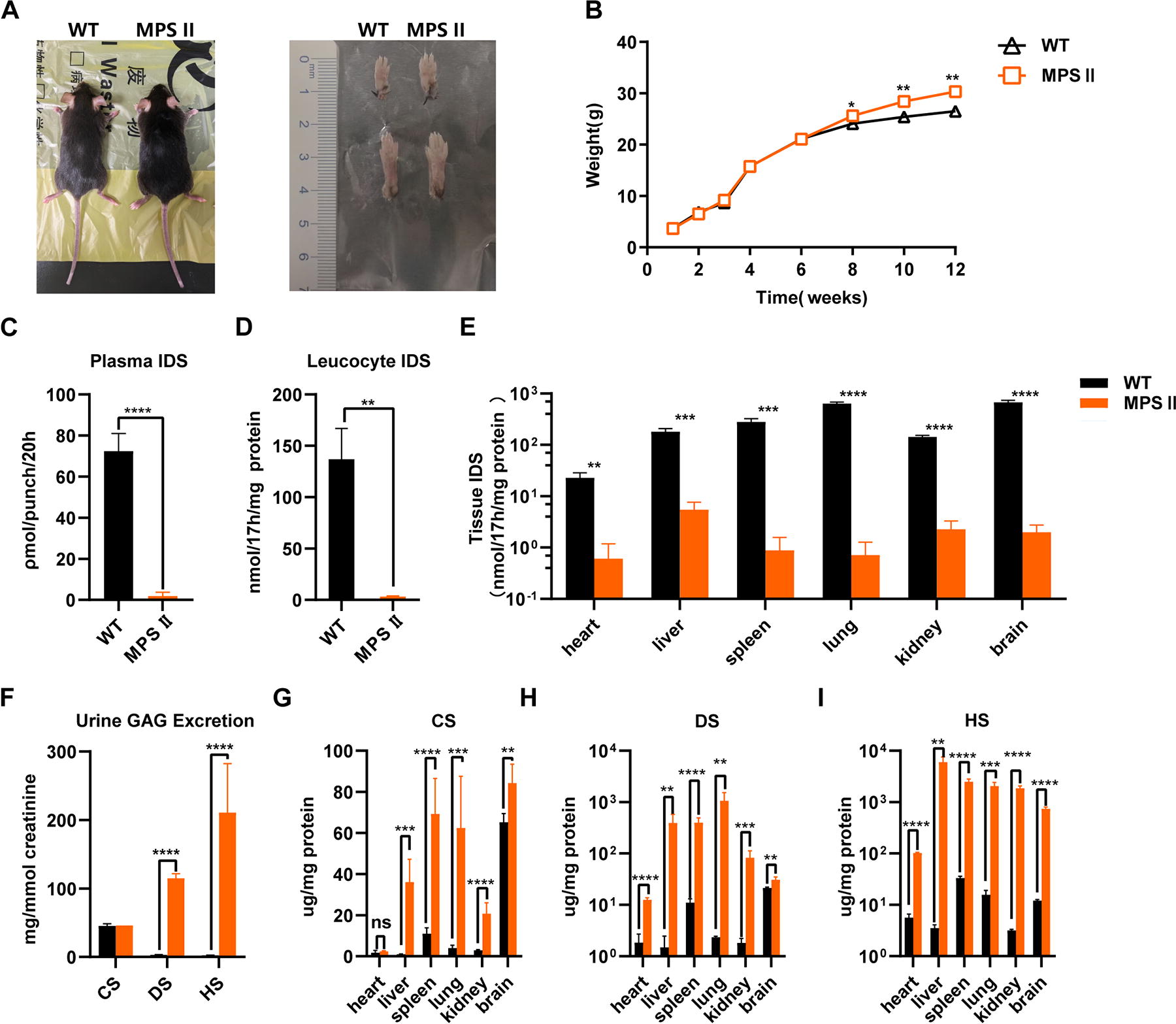
Starting from 8 weeks of age, MPS II mice exhibited significantly increased body weight, rough fur, and thicker toes, and toes were typically in a curled position compared with wild-type mice of the same background (Fig. 2A, B). This mouse model shows minimal residual IDS enzyme activities in plasma and leukocytes as in affected patients with MPS II. Almost all tissues showed IDS enzyme activity below 5.51 nmol/17h/mg. Some samples were undetectable or below our LOD (Fig. 2C–E). Consistent with the deficiency in IDS enzyme activity, tissues from MPS II mice showed the expected elevation in GAGs, with peripheral tissues and urine showing elevated levels of DS and HS. In contrast, HS levels were elevated mainly in the brain, with a less significant increase in DS. These findings are consistent with the Tanaka et al. study (Fig. 2F–I). 25
The primary finding of the microscopic examination of tissue sections taken from MPS II mice was the presence of foamy, vacuolated cell types infiltrating multiple organs and tissues (Fig. 3). Alcian Blue staining revealed elevated GAG levels, as shown by intense staining in various tissues, including the liver, lung, kidney, and brain (Fig. 4).

Tissue analysis after AAV2/9-hIDS treatment was performed using HE staining. Arrows indicated hippocampal neurons; triangles indicated hepatic macrophages. Scale bar = 20 μm.

Tissue analysis after AAV2/9-hIDS treatment was performed using Alcian Blue staining. Positive Alcian Blue staining suggested that GAG is stored in the tissue. Scale bar = 20 μm.
Previous MPS II mouse models are usually accompanied by skeletal dysplasia, 26,27 so we obtained skull and whole-body X-ray images from the MPS II mice. Compared with WT mice, MPS II mice showed significantly shortened femurs and elongated zygomatic bones, with no statistical difference in tibia length between the two groups (Supplementary Fig. S1A–C). In the MPS II mice, severe calcification of the calcaneus tendon was noted, and severe periosteal bone formation was seen at the lateral aspect of the distal tibia (Supplementary Fig. S1D).
To evaluate the neurocognitive deficits in MPS II mice, mice from each group underwent Morris water maze and Y-maze behavioral analyses at 6 months of age. MPS II mice had a longer latency to escape the maze compared with WT mice in the Morris water maze (particularly on the third day, the latency of MPS II mice was 48.55 s compared with 25.7 s in WT mice (p < 0.0001) (Supplementary Fig. S2A). On the sixth day, the WT mice showed a significant increase in the number of times crossing the platform within the same time frame compared with MPS II mice (p = 0.0023) (Supplementary Fig. S2B). Their swimming speed during the visible platform phase was significantly slower than that of WT mice (p < 0.0001) (Supplementary Fig. S2C). In the Y-maze experiment, the average spontaneous alternation rate for MPS II mice was 46.76%, while that for wild-type mice was 68.55%. The difference between the two groups was significant, with a difference of 21.79% (p < 0.0001) (Supplementary Fig. S2D). Therefore, we conclude that MPS II mice have significant neurocognitive deficits compared with WT littermates.
Collectively, these results suggest that this model possesses the pathological manifestations of human MPS II disease.
Prevention of CNS and peripheral symptoms in MPS II via ICV injection of AAV2/9-hIDS vector
In MPS II male mice aged 6–8 weeks, injection of 1 × 1011 VC AAV2/9-hIDS via ICV/IV resulted in widespread vector distribution. Following treatments, plasma IDS activity in MPS II mice was significantly higher than in WT mice, reaching levels up to 120-fold, and this expression persisted throughout the experiment (16 weeks post-injection) (Fig. 5A). In addition to the elevation in plasma IDS activity, significantly increased IDS activity in leukocytes was observed in both treatment groups of mice after 16 weeks of treatment (21-fold in the ICV group and 26-fold in the IV group) (Fig. 5B). Urine collection was conducted periodically to evaluate the long-term impact of IDS expression on urinary GAGs in treated mice. Both ICV and IV groups showed significant reductions in DS and HS levels in urine, which decreased to WT levels (p < 0.01)(Fig. 5E–G).

Plasma
At 6 months of age (16 weeks post-injection), all mice were euthanized, and organs were harvested for analysis. The IDS activity in different brain regions of mice injected with AAV2/9-hIDS via the ICV was approximately 95–157% of WT levels (Fig. 5C). However, supraphysiological levels of IDS activity were detected in all tested peripheral organs, with the highest observed being in the heart (250-fold) (Fig. 5D). These results of the IDS activity profile indicate that the ICV-administered AAV2/9-hIDS vector leaks into the systemic circulation, which is absorbed and expressed by peripheral organs. The mice treated with AAV2/9-hIDS administered by IV route did not reach WT levels in the CNS (6.68–66.27% of WT levels), while it was still sufficient to degrade accumulated GAG in the brain to normal levels. Similar IDS activity was detected in the peripheral tissues of the IV group, with no difference compared with the ICV group.
Due to the increased IDS activity, both ICV and IV treatment significantly reduced DS and HS levels in all tested tissues. However, there was no significant difference in DS and HS levels between WT mice and the two treatment groups (p > 0.05), indicating normalization of GAGs in all tested tissues at the time of sampling (Fig. 5H–M).
To assess post-treatment pathology, we performed Alcian Blue and HE staining of tissues (brain, liver, lung, and kidney). Significantly less GAG storage was observed in the tissues of ICV/IV mice compared with untreated MPS II mice (Fig. 4); in the MPS II mouse model, GAG accumulation results in neuronal or cellular vacuolization formation. Vacuolization of both hippocampal pyramidal cells and peripheral tissue cells was significantly reduced in treated mice (Fig. 3).
DNA was isolated from the same tissue homogenates, and vector distribution was assessed using qPCR. Low copy numbers were detected in most tested peripheral tissues, while high copy numbers were observed in the liver. This indicated that AAV2/9-hIDS primarily transferred to the liver, produced IDS, and released it into circulation, where it was subsequently absorbed by various tissues, consistent with previous reports (Fig. 6A). 28,29

In addition to increased IDS activity and decreased GAG levels, mice in the treatment cohorts exhibited a decelerated rate of weight gain, eventually showing no discernible distinction in weight compared with WT mice by the 6-month-old (Fig. 6B). The relative proportions of the brain, heart, lungs, and kidneys did not exhibit notable variances across groups. However, untreated MPS II mice displayed a liver weight 45% larger than WT mice (constituting 6.01% and 4.13% of total body weight, respectively). Mice undergoing IV and ICV for MPS II demonstrated a slight reduction in liver weight, with 4.85% for the ICV group and 4.46% for the IV group (Fig. 6C).
Sustained expression of IDS in the CNS prevents neurocognitive deficits and improves hearing in ICV-treated MPS II mice
At 6 months of age (4 months after injection), untreated MPS II mice, AAV2/9-hIDS-treated MPS II mice, and WT mice were evaluated for neurocognitive function using the Morris water maze and Y-maze. On day one, all mice demonstrated initial maze learning skills across four trials, but there were no significant differences in escape latency and speed between the groups. WT mice showed a shortened escape latency on the fourth day (21.88 s), whereas untreated MPS II mice exhibited significant deficits in learning this task (latency to escape reduced to 33.24 s). In contrast, ICV group mice showed a significant reduction in escape latency (17.45 s), outperforming MPS II mice (p = 0.0102). Mice in the IV group also showed a reduction (30.60 s) but without significant difference compared with untreated MPS II mice (p = 0.6380) (Fig. 6D). On the sixth day, the underwater platform was removed. Compared with the untreated group, the IV group showed a significant increase in the number of times crossing the platform within the same time frame. The ICV group showed an increase as well without a significant difference (Fig. 6E). Additionally, the swimming speed of mice in the IV group was significantly faster than that of the MPS II group (p = 0.0016), while no statistical difference was observed between the ICV group and the MPS II group (p = 0.2634) (Fig. 6F). The ICV and IV groups showed higher proportions of time spent and distance traveled in the quadrant where the platform was located than untreated MPS II mice (Fig. 6G, H). The spontaneous alternation rates of mice in the ICV group and IV group reached 62.33% and 66.12% (p = 0.3774, ICV vs. IV), respectively, much higher than the average spontaneous alternation rate of MPS II mice (48.7%) (p < 0.01) and close to that of WT mice (63.6%) (Fig. 6I). Therefore, ICV resulted in sustained, normal-level IDS activity in the CNS that preventing further neurocognitive impairment in MPS II mice.
Remarkably, following 4 months of treatment, the ICV group exhibited significantly decreased hearing thresholds at frequencies of 8, 16, and 32 kHz compared with MPS II mice (p < 0.001), whereas the IV group did not show a significant reduction in hearing thresholds (Fig. 6J).
DISCUSSION
In this study, we developed an MPS II murine model and validated its phenotype to create an in vivo platform for investigating novel therapies. We use this new MPS II murine model to examine a gene therapy approach by administering the AAV2/9-hIDS to treat both CNS and somatic disease phenotypes, comparing the ICV with the IV administration routes. Both treatments effectively increased IDS enzyme activity and reduced the pathological accumulation of GAGs in MPS II mice. Notably, sustained expression of IDS could effectively improve behavior performance in the Morris water maze.
We established a novel p.R513Q MPS II mouse model with an IDS pathogenic variant analogous to the human IDS p.R468W and p.R468Q variants. 30,31 The codon 468 position is a “hot” spot in Asian patients with MPS II and is often associated with the severe form of the disease. This is the first MPS II mouse model engineered with a point mutation. The p.R513Q MPS II mouse exhibits significantly reduced IDS activity in plasma, leukocytes, and several organs and tissues and markedly elevated GAG levels in urine and tissues (Fig. 2C–I). In addition, the p.R513Q MPS II mouse demonstrates neurocognitive deficits in spatial learning and memory in the Morris water maze and working memory in the Y-maze (Supplementary Fig. S2). Overall, the p.R513Q MPS II mouse phenotype is consistent with previously established MPS II mouse models. 27,32 Future studies on therapeutic strategy development can utilize this model.
In our study, regardless of the route of administration, MPS II mice treated with ICV and IV showed significantly higher plasma IDS levels than the WT mice group (93-fold increase in the ICV group and 126-fold increase in the IV group), indicating successful administration of ICV/IV. Motas et al. (in coronal sections of the brain) 33 and Hinderer et al. (the whole brain) 34 reported comparable changes in IDS activity, where they observed approximately 20–40% of WT levels in the CNS. We conducted a more in-depth analysis of various brain regions; IDS enzyme activity in all five brain regions of the ICV group approached normal levels. This may be due to the CBh strong promoter used in the vectors from these previous studies. The CBh promoter provides strong expression in ex vivo rat hippocampal slices and long-term, widespread expression in the CNS, thereby promoting the expression of IDS in the CNS. 35,36 Although the IV group showed lower IDS levels in brain regions (6.68–66.27%), the extent of GAG reduction exceeded expectations (Fig. 5C, D). Desnick et al. and Polito et al. suggested that correction of GAG pathological accumulation may require less than 5% of WT lysosomal enzyme levels. 37,38 Our results are consistent with this hypothesis.
Unlike previous studies that used ELISA to measure total GAG, we employed mass spectrometry to detect this biomarker, which can analyze three types of GAGs (CS, DS, and HS). GAG accumulation in the CNS was mainly composed of HS, while in the peripheral tissues and urine, it was mostly DS and HS. After treatment, DS and HS decreased to levels comparable to those in WT mice. This method allows for a more sensitive detection of variations in various GAGs.
Deposits of GAGs in mucosal linings, connective tissue, cartilages, bones, and the CNS led to many components contributing to hearing loss. Nearly all patients with MPS II manifest with pure sensorineural hearing loss, often characterized by progressive deterioration. 39,40 In addition to the accumulation of GAGs in the neurovascular structures of the inner ear, which damages the organ of Corti, spiral ganglion, and stria vascularis, Chimelo et al. found that the prolonged wave latency of brainstem auditory evoked potentials in patients with MPS II, indicating widespread impairment in the higher/lower brainstem functions between the distal auditory nerves and the cochlear nucleus. 41 In our study, the increased IDS enzyme activity in the CNS of mice in the ICV group promoted a reduction in GAG levels, improving the auditory function in the murine MPS II model. IV group did not show a significant decrease in hearing thresholds. This may be due to the high expression of IDS in the CNS of mice after ICV injection, which improved the brainstem auditory pathways.
In conclusion, our study demonstrates the efficacy of MPS II treatment using AAV2/9-hIDS, which induces mice to reduce neurological deficits. In the ICV group, IDS enzyme activity in the CNS increased significantly and reached WT levels. We anticipate that this study will contribute to the field’s efforts to develop a long-term effective treatment that will provide neurological benefits to patients with MPS II.
Footnotes
ACKNOWLEDGMENT
The authors thank Caygen for providing the p.R513Q MPS II mouse.
AUTHORS’ CONTRIBUTIONS
G.C. and G.M. drafted the article. H.Z. designed the study and revised the article. G.C. and X.Z. performed the experiments and statistical analyses. Y.C. and Q.L. performed the vector testing. J.Y. and S.H. performed the ABR testing. H.L. and M.Y. modified the figures. All data were generated in-house, and no paper mill was used. All authors agree to be accountable for all aspects of this study, ensuring its integrity and accuracy.
AUTHOR DISCLOSURE
The authors declare no competing interests.
FUNDING INFORMATION
This work was supported by the Shanghai Municipal Health Commission [No.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.