
Abstract
Gene therapy using adeno-associated virus (AAV) vectors is currently expanding to broad clinical applications. As the presence of a neutralizing antibody (NAb) against AAV capsids significantly restrains their efficacy, an accurate evaluation of NAb status is crucial for selecting appropriate candidates for gene therapy. Notably, cell-based NAb assays may not be sufficiently sensitive for detecting low-titer NAb, and few assays can evaluate multiple AAV serotypes using a commonly available cell. In this study, we developed a sensitive NAb assay against various AAV serotypes using commonly available HEK293 and Huh-7 cells. We found that adding glucose efficiently enhanced transgene expression across various AAV serotypes without causing cell damage. In addition, by combining a highly sensitive reporter gene, NanoLuc, the necessary dose of AAV vector was significantly reduced. The reduction of AAV dose resulted in the increased sensitivity of NAb detection as low as 100 vector genomes/cell. At the lower vector doses, sensitivity improvement was not observed regardless of serotypes, suggesting the limit of assay sensitivity of the cell-based NAb assay. These findings provide a highly sensitive methodology for assessing NAb titers and offer insights into conditions to attain maximal sensitivity in the cell-based NAb assay.
INTRODUCTION
Gene therapy using adeno-associated virus (AAV) vectors is an area of active research worldwide with expanding clinical applications for various diseases. 1 AAV vectors have a high safety profile and transduce various cells, and long-term transgene expression is expected. 1 Wild-type viruses of AAV are widely distributed in nature, and a significant portion of the human population harbors neutralizing antibodies (NAbs) against AAV capsids. 2 Early exposure to AAV 3,4 and the prevalence of NAb are well-documented to be in approximately half of the human population. 5 The presence of NAb significantly reduces the transduction efficiency of AAV vectors, leading many clinical trials to exclude NAb-positive patients. 2,6 Various strategies are being explored to conquer the action of NAb and proceed with the application of AAV-mediated gene therapy. 6 –9
Accurate NAb level measurement is crucial to identifying suitable AAV-mediated gene therapy candidates. Several methods have been studied internationally, and optimal techniques have been employed in clinical trials. 5,10,11 These methods include cell-based NAb assays, 10 enzyme-linked immunosorbent assay (ELISA)-based assays, 12 quantitative polymerase chain reaction (qPCR) assays to assess AAV binding to cells, 13 and in vivo inhibition assays. 14 Among these, cell-based NAb assays are widely used because they are easy to establish, directly assess NAb activity, and provide relatively consistent results. 2,5,15 These assays measure AAV gene expression inhibition after preincubating the test serum with AAV vectors carrying a reporter gene. Currently, cell-based assays may lack sensitivity, owing to the lack of suitable cells for transduction, particularly in AAV5, AAV8, or AAV9, necessitating extremely high vector doses to achieve sufficient gene expression. 15,16 In addition, an assay that can evaluate multiple AAV serotypes with commonly available cells is preferred.
Cell lines, such as HEK293, Huh-7, and HeLa, are used in cell-based NAb assays, depending on the infectivity of the AAV serotype. 14 To enhance AAV infectivity, HEK293-derived cell lines, such as 2V6.11, expressing the adenovirus E4 ORF gene under an ecdysone-inducible promoter, 17,18 and AAVR-HeLa cells, overexpressing the AAV receptor (AAVR), 19 have been utilized. Other methods such as the addition of transduction-enhancing reagents, wild-type adenovirus 20 and dorsomorphin, 21 have also been explored.
As for the reporter genes, green fluorescent protein (GFP), LacZ, or luciferase have been utilized. Luciferase is superior to GFP in terms of assay sensitivity. 14 The NanoLuc reporter gene, which is expressed over 100 times more than luciferase, provides even higher sensitivity. 22,23
Osmotic stress to cells can activate intracellular signaling pathways to maintain osmotic pressure and homeostasis by regulating intracellular electrolytes, 24 and saccharide-mediated transgene expression enhancement was demonstrated in plasmid transfection, by inducing osmotic stress. 25 –27 Osmotic stress also improves the virus production capacity. 28,29 Thus, osmotic stress in cells may affect AAV-mediated transgene expression. Therefore, this study aimed to develop a sensitive NAb assay for various AAV serotypes using commonly available cell lines, by combining glucose-induced AAV vector gene expression with a highly sensitive reporter gene, the secreted NanoLuc (secNanoLuc).
MATERIALS AND METHODS
Cell lines, plasmids, and AAV vector preparation
HEK293 cells and Huh-7 cells (ATCC, Manassas, VA) were cultured in Dulbecco’s modified eagle’s medium (Sigma-Aldrich, St. Louis, MO) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA). NaCl and various saccharides used in this study were purchased from Sigma-Aldrich and were dissolved in double distilled water, filtered, and sterilized to prepare stock solutions. AAV vectors encoding enhanced green fluorescent protein (EGFP), LacZ, luciferase, and secNanoLuc driven by the cytomegalovirus (CMV) promoter were prepared as previously described. 30 Titration of recombinant AAV vectors was performed using qPCR with the gene-specific CMV promoter primers and TB Green® Premix Ex Taq™ II (Takara Bio, Kusatsu, Japan) as detailed previously. 31
AAV vector transduction and measurement of transgene expression in vitro
One day before transduction, 5 × 104 HEK293 or Huh-7 cells were seeded into a 96-well culture plate. On the transduction day, 10 µL of the AAV vector encoding the reporter gene was added directly to the wells.
Firefly luciferase assays were performed as indicated by the Luciferase Assay System (Promega, Madison, WI), and NanoLuc assays were performed as indicated by the Nano-Glo Luciferase Assay Kit (Promega). The luminescence was measured using a Spark luminometer (Tecan, Männedorf, Switzerland).
β-Galactosidase activity was quantified as indicated by the β-Gal Assay Kit (Invitrogen, Carlsbad, CA), and the absorbance was measured at 405 nm using a microplate reader SpectraMax 190 (Molecular Devices, San Jose, CA).
EGFP-positive cells were observed under a fluorescence microscope IX73 (Olympus, Tokyo, Japan), and the ratio of EGFP-positive cells was calculated in a Disposable Hemocytometer C-chip (NanoEntek, Seoul, Korea).
Assay for NAb measurement
The NAb titers were evaluated using human immunoglobulin (γ-globulin from human plasma; Fujifilm Wako Pure Chemical Corporation, Osaka, Japan), and anti-AAV mouse monoclonal antibodies (ADK1a against AAV1, A20 against AAV2, ADK5a against AAV5, ADK8 against AAV8, ADK9 against AAV9; PROGEN Biotechnik, Heidelberg, Germany). Experiments using mice and microminipigs were approved by the Institutional Animal Care and Concern Committees of Jichi Medical University and performed per the guidelines provided by the committees. Human immunoglobulin was adjusted to 50 mg/mL, and mouse monoclonal antibodies were reconstituted to 50 µg/mL.
The NAb assay was performed as previously described. 32 Briefly, 5 × 104 HEK293 or Huh-7 cells were seeded in a 96-well plate the day before transduction. On the transduction day, 10 µL of the test sample was diluted serially with FBS, and 10 µL of AAV vector encoding the reporter gene was mixed, incubated at 37°C for 1 h, and then added to the wells. After AAV transduction, the NaCl or saccharide solution was added to the culture medium, and the concentration, including that in the medium, was determined. After 48 h, AAV gene expression was measured. A sample was considered positive for neutralizing activity if it inhibited gene expression by over 50% compared with the control FBS. The NAb titer was reported as the final serum dilution in the culture medium that exhibited neutralizing activity.
Trypan blue exclusion assay and in vitro cell counting assay
HEK293 or Huh-7 cells were seeded in a 96-well plate at a density of 5 × 103 cells/well and transduced with AAV2 at 1000 vector genomes (vg)/cell. Glucose solution was added to each culture medium 1 h after AAV transduction. Then, 48 h post-transduction, the cells in a trypan blue solution (0.4%) were placed on a Disposable Hemocytometer C-chip (NanoEntek), and the percentages of live and dead cells were calculated using a fluorescence microscope IX73 (Olympus). Cell numbers were evaluated as indicated by the Cell Counting Kit-8 (Dojindo Laboratories, Kumamoto, Japan), and the absorbance was measured at 450 nm using a microplate reader SpectraMax 190 (Molecular Devices).
Total anti-AAV antibody detection
Total anti-AAV antibodies were quantified by ELISA as previously described. 33 To detect human immunoglobulin, mouse monoclonal antibody, or pig immunoglobulin that binds to AAV capsids, anti-human IgG (Abcam, Cambridge, United Kingdom), anti-mouse IgG (Proteintech group, Rosemont, IL), or anti-pig IgG (Sigma-Aldrich) conjugated with horseradish peroxidase were used, respectively.
Statistical analysis
Statistical analyses were performed with GraphPad Prism 9.5.0 (GraphPad Software, San Diego, CA). Student’s t-test was used to compare EGFP expression levels. Statistical comparisons between groups were conducted using a one-way analysis of variance with a post hoc Tukey test. Statistical significance was set at p < 0.05.
RESULTS
Glucose effectively enhanced the transgene expression of the AAV vector in HEK293 and Huh-7 cells
We investigated whether simultaneous administration of NaCl and various saccharides (glucose, galactose, sucrose, trehalose, sorbitol, mannitol, and xylitol) in a culture medium of HEK293 and Huh-7 cells improves transgene expression by the AAV2-CMV-LacZ vector and found that transgene expression was effectively induced at glucose concentrations ranging from 150 to 200 mM in both HEK293 and Huh-7 cells (Fig. 1a). As for the optimal timing of glucose administration, five time points ranging between 3 h before and after AAV transduction were tested. Although statistically insignificant, the transgene expression was slightly higher at 1 h after AAV addition in both cells (Fig. 1b). Therefore, we decided to add 175 mM glucose at this point in subsequent experiments. When HEK293 and Huh-7 cells were transduced with AAV2-CMV-EGFP, 175 mM of glucose significantly increased the number of EGFP-positive cells (Fig. 1c).
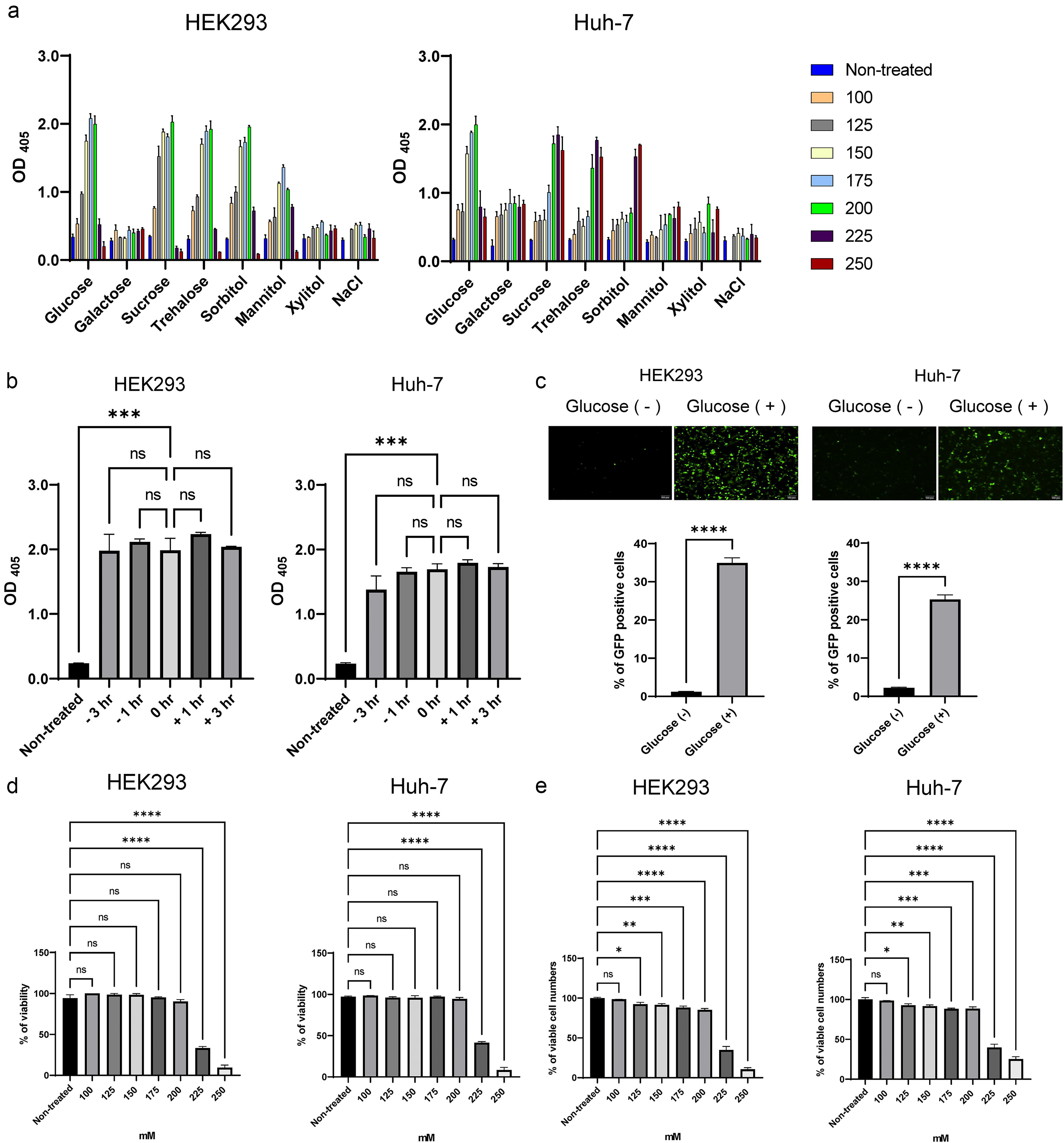
Evaluation of the effect of glucose on AAV gene expression.
Cell viability was not affected at glucose concentrations of 100–200 mM, whereas it was suppressed above 225 mM in trypan blue exclusion assay (Fig. 1d). In the in vitro cell counting assay, viable cell numbers were decreased, especially above 225 mM in both cells (Fig. 1e). Although viable cell number was significantly reduced at 175 mM of glucose (Fig. 1e), cell death was not induced (Fig. 1d), suggesting that this glucose concentration suppressed cell growth without causing cell death.
Addition of glucose reduced necessary vector doses of AAV5, AAV8, and AAV9, leading to increased sensitivity of the NAb assay
Next, we tested whether we could improve the sensitivity of the NAb assay by reducing the vector dose. As a result, gene expression for each serotype revealed that the addition of 175 mM glucose effectively induced the AAV5, AAV8, and AAV9 expressions with both the luciferase and secNanoLuc genes (Fig. 2a, b). Given that a stable NAb assay requires a relative light unit (RLU) threshold of 10,000, 18,22 we tested viral vector doses that achieved this RLU level. Glucose-induced gene expression allowed sufficient luciferase gene expression ranging from 10,000 to 100,000 vg/cell for AAV5 and from 1,000 to 10,000 vg/cell for AAV8 and AAV9. Using highly sensitive reporter gene secNanoLuc, a sufficient expression level was attained from 100 to 1,000 vg/cell for AAV5 and from 10 to 100 vg/cell for AAV8 and AAV9 (Fig. 2a, b). Based on these results, we used the AAV-CMV-secNanoLuc vector in the assays at doses at which gene expression exceeded 10,000 RLU. To evaluate NAb sensitivity in cell-based assays, we measured NAb titers using human immunoglobulins and anti-AAV mouse monoclonal antibodies. The NAb titer was expressed as the final dilution ratio of the antibody that achieved 50% neutralizing inhibition of transgene expression (ND50). 22 For this purpose, mean ND50 values (n = 5) with and without glucose were compared.

Evaluation of NAb assay for AAV5, AAV8, and AAV9 capsids.
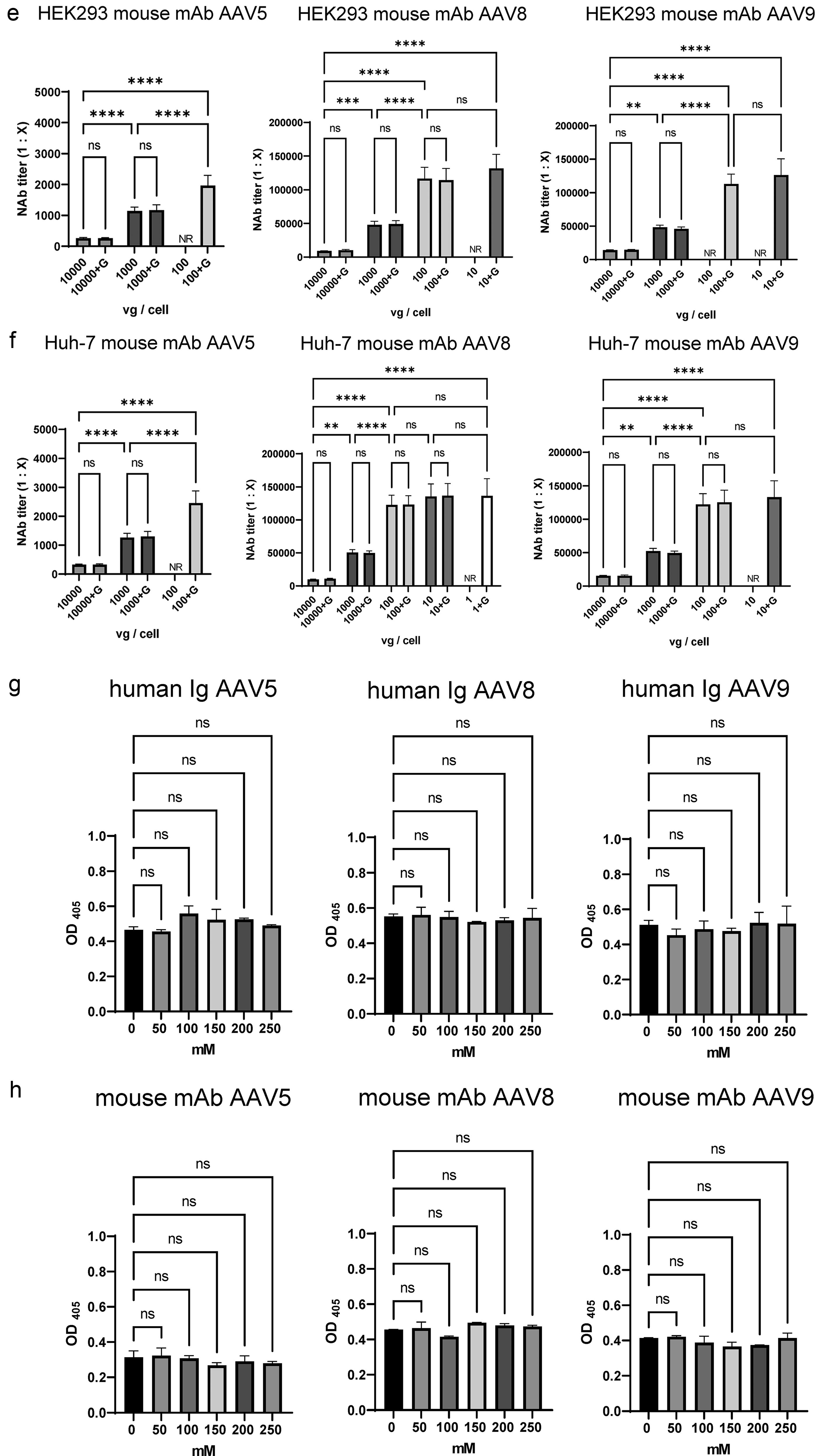
(Continued).
In HEK293 cells, the increased sensitivity was observed as the AAV vector dose decreased until 100 vg/cell (Fig. 2c). The addition of glucose in AAV5 and AAV9 at 100 vg/cell resulted in transgene expression levels beyond 10,000 RLU, allowing the determination of the NAb titer of human immunoglobulin (Fig. 2b, c). The same was true with anti-AAV mouse monoclonal antibodies specific for AAV5, AAV8, and AAV9 (Fig. 2b, d).
In Huh-7 cells, the increased sensitivity of the NAb assay was also observed with decreasing AAV vector doses until 100 vg/cell (Fig. 2b, e). Transgene expression of AAV5 at 100 vg/cell exceeded 10,000 RLU with glucose addition, improving the NAb assay sensitivity (Fig. 2e). This trend is consistent with that observed for the anti-AAV mouse monoclonal antibody (Fig. 2b, f). No significant differences in NAb sensitivity were observed in both cells for every serotype tested by the addition of glucose.
To evaluate the action of glucose, an ELISA was conducted at various glucose concentrations. The results indicated that the glucose concentrations used in the NAb assay did not adversely affect AAV capsid binding to anti-AAV antibodies (Fig. 2g, h).
Cell-based NAb assay using glucose worked well with animal serum
After demonstrating the utility of the NAb assay with purified human immunoglobulins and anti-AAV mouse monoclonal antibodies for AAV5, AAV8, and AAV9, we evaluated its effectiveness in other species, using serum from mice and microminipigs. We conducted experiments using animal sera in which NAb titers were previously measured without glucose addition. Sera were obtained from Fabry disease model mice administered AAV9 with NAb titers estimated at 1:44,800 or 1:89,600 in a previous study. 34 The assay using AAV at a reduced dose of 100 vg/cell showed a significantly higher NAb titer than that obtained at 1000 vg/cell (1:83,263 to 1:170,141 in HEK293 and 1:83,904 to 1:181,936 in Huh-7) (Fig. 3a). In microminipigs, sera were obtained from untreated animals with low NAb titers, which were estimated at 1:14 in a previous study. 33 Again, significantly increased NAb titers were obtained at the dose of 100 vg/cell than that obtained at 1000 vg/cell (1:20 to 1:54 in HEK293 and 1:16 to 1:52 in Huh-7), suggesting that this assay is effective and that reducing the vector dose could increase NAb sensitivity in different animal sera. ELISA confirmed that the glucose concentration used did not affect AAV capsid binding to anti-AAV antibodies (Fig. 3c, d).
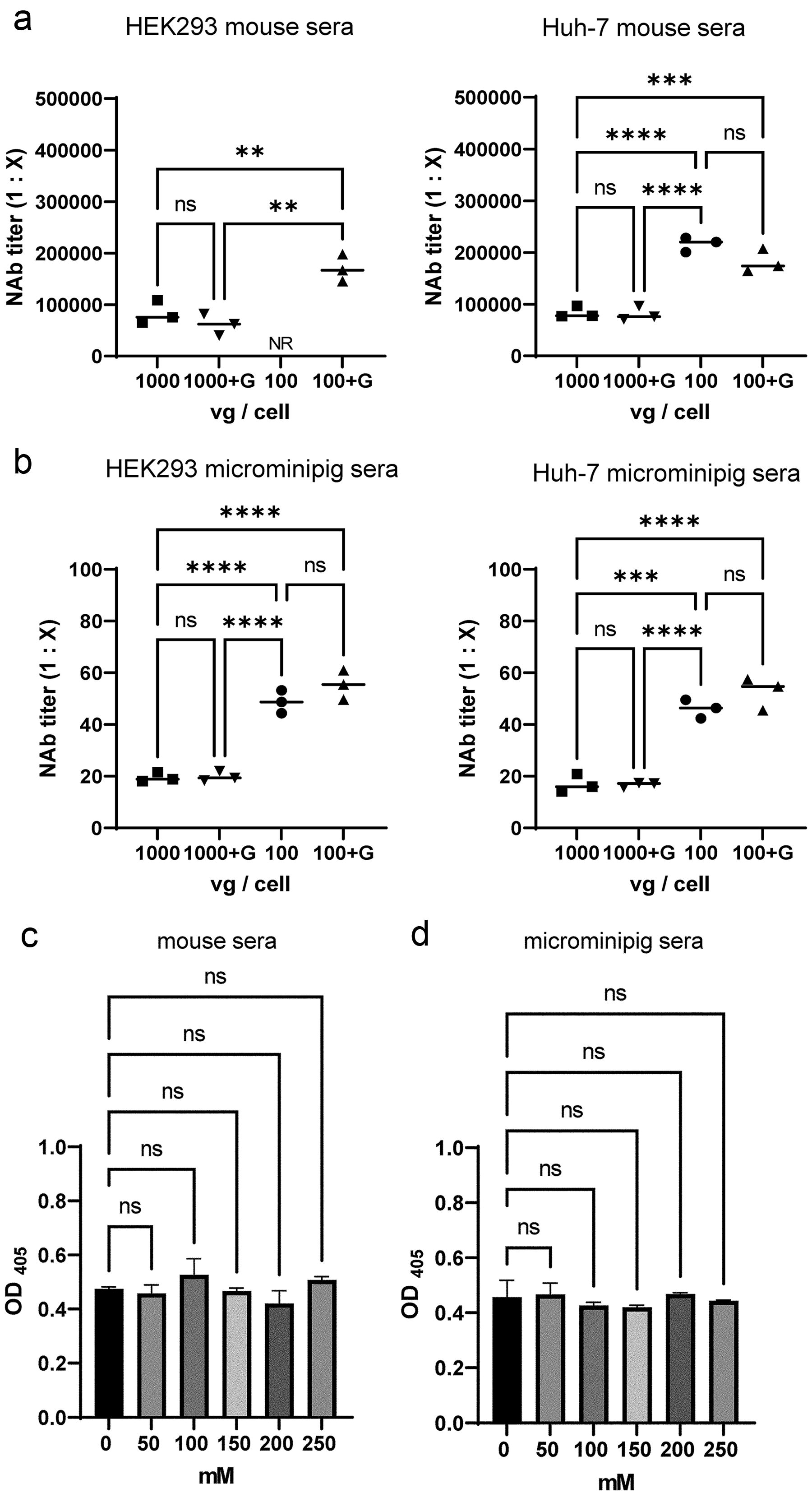
Evaluation of NAb assay for mice and microminipig sera.
Assay sensitivity reached the maximum at 100 vg/cell, regardless of AAV serotypes
Given that the NAb assay sensitivity did not increase even when viral vector doses were reduced to ≤100 vg/cell for AAV5, AAV8, and AAV9, we investigated whether similar results could be obtained by NAb assays with AAV1 and AAV2, which have much higher infectivity in HEK293 and Huh-7 cells. Transgene expression from AAV1 and AAV2 encoding Luciferase or secNanoLuc was also enhanced with 175 mM glucose (Fig. 4a, b). Therefore, cell-based assays were performed at doses achieving 10,000 RLU, as low as 1 vg/cell. In HEK293 cells, increased sensitivity was observed as the AAV vector dose decreased until 100 vg/cell (Fig. 4c, e). Interestingly, at the lower vector doses, no improvement of assay sensitivity was observed in both serotypes (Fig. 4c, e). This trend was consistent with Huh-7 cells (Fig. 4d) and anti-AAV mouse monoclonal antibodies (Fig. 4e, f). Overall, the NAb assay sensitivity did not increase to <100 vg/cell, regardless of the AAV serotype or antibody in this study. As other serotypes tested, glucose concentration did not affect the binding of AAV capsids to anti-AAV antibodies in AAV1 and AAV2 by ELISA (Fig. 4g, h).
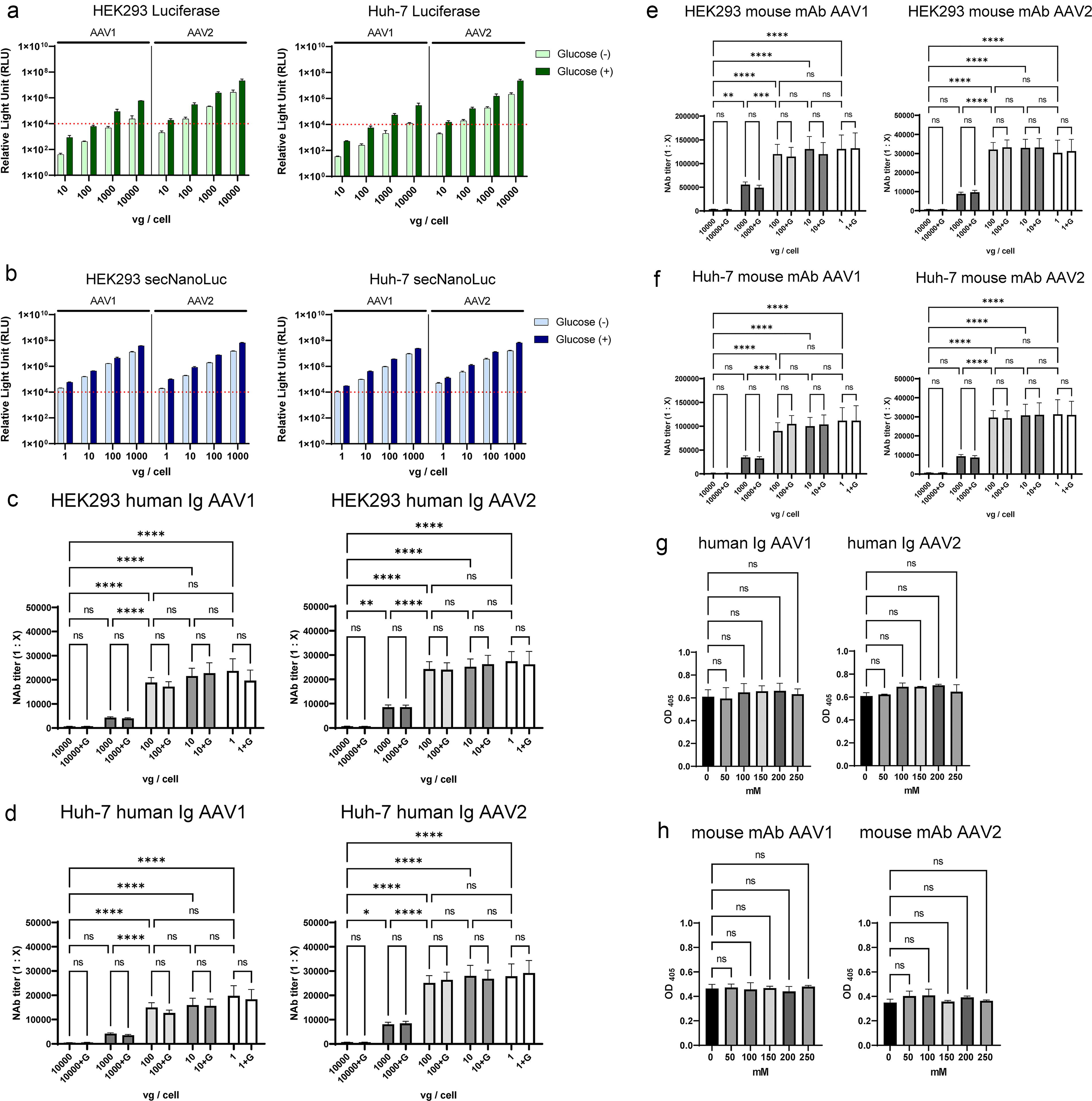
Evaluation of NAb assay for AAV1 and AAV2 capsids.
DISCUSSION
The presence of NAb against AAV capsid presents a significant challenge in gene therapy, particularly during systemic administration. Many clinical trials have excluded NAb-positive patients. Therefore, an accurate NAb status assessment before AAV vector administration is crucial for selecting appropriate candidates. 2,10,11 Cell-based NAb assays are valuable because they can directly reflect AAV transduction inhibition. However, AAV serotypes that lack sensitive cells for transduction often require extremely high vector doses to achieve sufficient gene expression, which can limit the detection of low-titer NAb against these capsids.
Our study revealed that adding 175 mM glucose in the culture medium enhanced gene expression and thus reduced the AAV vector dose in both HEK293 and Huh-7 cells. This is particularly beneficial for AAV serotypes with low infectivity to the cells, such as AAV5 (Fig. 2c–f) and AAV9 (Fig. 2c, e).
To our knowledge, this is the first study to demonstrate the utility of glucose to effectively induce AAV gene expression in cell-based NAb assays. If glucose-mediated enhancement applies to in vivo settings, there may be a great impact on AAV-mediated gene therapy. Nonetheless, as blood glucose levels are kept less than 10 mM in animals and humans, attaining a 175 mM level seems challenging. Therefore, the application of this phenomenon may be confined to in vitro usage.
In this study, the mechanism through which saccharides induce AAV gene expression remains unclear. Saccharide-induced osmotic stress activates various signaling pathways involved in cellular homeostasis. 24 Osmotic stress improves antibody production by increasing metabolism, transcription, and translation. 25 –27,35 Research suggests that high osmolarity conditions can enhance the specific antibody productivity of plasmids used in antibody production. While the exact mechanism is not fully understood, these conditions are believed to improve mRNA transcription rates 35 and significantly increase protein translation. 26 Similar mechanisms may be involved in the AAV-mediated gene expression. Further research on the intracellular effects of glucose may help elucidate these mechanisms.
Notably, decreasing the AAV vector dose <100 vg/cell did not improve the assay sensitivity, regardless of the AAV serotype or antibody species. This trend was also observed in a previous study with AAVR-expressing HEK293 cells using AAV8 vectors. 22 In that study, the coefficient of variation of the assay worsened with reduced viral doses, concluding that the optimal viral dose for the cell-based assay was 100 vg/cell. 22 In this study, we also obtained reliable results at the lower vector doses, and assay sensitivity remained constant. Although the reason for this phenomenon is currently unclear, we speculate that at extremely low vector doses, the property of the antibody itself is the limiting factor of the assay sensitivity. In this case, the amount of antibody (Ab) that binds to the antigen (Ag) correlates to the inverse of the dissociation constant (KD) value (Ab + Ag ⇆ AbAg, [AbAg]/[Ab][Ag] = 1/KD). When the amount of AAV per cell was sufficiently small and the amount of antibody was constant, the NAb titer depended on the KD value. Based on our results, the NAb titer for each AAV serotype at 100 vg/cell with glucose may correlate with the inverse KD value for each mouse monoclonal anti-AAV antibody (Table 1), suggesting that 100 vg/cell in this cell-based NAb assay reaches close to the detection limit of NAb.
List of mouse monoclonal antibodies used in this study
The product names of anti-AAV mouse monoclonal antibody.
AAV, adeno-associated virus; KD, dissociation constant as listed in product sheets; mAb, anti-AAV monoclonal antibody; NAb, neutralizing antibody; N.A., not available.
This study provides a sensitive approach to assessing NAb titers when selecting suitable candidates for clinical and preclinical trials. This approach can help accurately identify patients with low NAb titers and study the range of low NAb titers capable of AAV gene expression by intravenous administration and will expand the number of candidates eligible for AAV-mediated gene therapy.
CONCLUSIONS
Our study identified adding glucose-enhanced gene expression in AAV serotypes with low sensitivity of transduction in HEK293 and Huh-7 cells. By combining secNanoLuc, we developed a sensitive cell-based NAb assay with reduced vector dose across various serotypes. The assay sensitivity demonstrated improvement until the dose of 100 vg/cell. As the sensitivity remained constant at lower doses, 100 vg/cell may be a practical goal for developing NAb assay with maximal sensitivity. Further testing with human samples will confirm the utility of this assay for AAV-mediated gene therapy.
Footnotes
ACKNOWLEDGMENT
The authors thank Yoko Haruta, Momoko Aoyama, Hiroki Daimon, and Akiko Ogasahara (Jichi Medical University) for their excellent technical assistance.
AUTHORS’ CONTRIBUTIONS
R.W. and H.M. conceived and designed the study. R.W. performed the experiments, analyzed the data, and wrote the article. K.O., Y. Sehara, Y.H., Y. Saga, M.U., T.O., and H.M. contributed to the concept, analyzed the results, and revised the article. All authors read and approved the final version of the article.
AUTHOR DISCLOSURE
The authors have no conflicts of interest to declare.
FUNDING INFORMATION
This research was supported by the Jichi Medical University Young Investigator Award, the