
Abstract
The advent of genome-editing technologies, particularly the RNA-guided the clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated system (Cas) 9, which originates from prokaryotic CRISPR/Cas adaptive immune mechanisms, has revolutionized molecular biology. Renowned for its simplicity, cost-effectiveness, and capacity for multiplexed gene editing, CRISPR/Cas9 has emerged as the most versatile and widely adopted genome-editing platform. Its applications span fundamental research, biotechnology, medicine, and therapeutics. This review highlights recent advancements in CRISPR-based technologies, focusing on CRISPR/Cas9, CRISPR/Cas12a, and CRISPR/Cas12f. It emphasizes precision editing methods like base editing and prime editing, which enable targeted nucleotide changes without double-strand breaks. The specificity of these tools, including on-target accuracy and off-target risks, is critically evaluated. Additionally, recent preclinical and clinical efforts to treat diseases such as cancer and sickle cell disease using CRISPR are summarized. Finally, the challenges and future directions of CRISPR-mediated gene therapy are discussed, emphasizing its potential to integrate with other molecular approaches to address unmet medical needs.
INTRODUCTION
Gene therapy holds significant promise for addressing a growing number of genetic disorders. However, conventional gene therapy approaches are constrained by their inability to correct existing mutations or precisely integrate new genes into targeted genomic loci within cells. 1 The advent of genome-editing technologies, which enable targeted in situ modifications of DNA sequences, has redefined the scope of gene therapy and marked the beginning of a transformative era. 2 –5 Currently, three primary genome-editing tools are available: zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and the clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated system (Cas). 6 These technologies have significantly advanced the precision and versatility of genetic interventions.
In contrast to ZFNs and TALENs, which depend on protein-DNA interactions for target recognition, the CRISPR/Cas utilizes a guide RNA to identify specific DNA sequences. This approach allows for the targeting of diverse DNA sequences simply by modifying a short segment of the guide RNA. 7 Owing to its simplicity, cost-effectiveness, high efficiency, and ability to perform multiplexed gene editing, CRISPR/Cas has largely replaced earlier methods and emerged as the most versatile and promising genome-editing technology.
Each genetic disease possesses distinct characteristics, necessitating the development of diverse genome-editing tools capable of precise genetic modifications. In this review, we present an overview of current CRISPR-based technologies, including CRISPR/Cas9, CRISPR/Cas12a, CRISPR/Cas12f, base editing, and prime editing. We also highlight recent advancements in disease treatment achieved through CRISPR applications. Finally, we discuss the current challenges and future prospects of CRISPR-based technologies in the context of gene therapy.
CRISPR GENE-EDITING TECHNOLOGY
CRISPR/Cas serves as an adaptive immune mechanism in bacteria and archaea, providing defense against viral infections. This system comprises two essential components: Cas genes, organized into operons, and CRISPR arrays, which consist of targeting sequences (spacers) derived from invading phages, interspersed with identical repeats. 8 When a spacer matches foreign DNA, Cas proteins silence the invading genetic material. Based on the conservation and organization of Cas genes, naturally occurring CRISPR/Cas are classified into six types. 9 Among these, Cas9 from the type II CRISPR/Cas stands out for its ability to directly cleave foreign DNA at specific sites, making it the simplest and most widely adopted CRISPR system. This mechanism provides bacteria with a sophisticated and efficient defense against phages. In a groundbreaking study, Doudna and Charpentier purified Cas9 from Streptococcus pyogenes and streamlined the dual tracrRNA:crRNA into a single guide RNA (sgRNA), demonstrating that Cas9 could induce sequence-specific double-strand breaks (DSBs) in vitro. 8 Their discovery of the RNA-programmable CRISPR/Cas9 system, which earned them the Nobel Prize in Chemistry just eight years later, has revolutionized genetic engineering, providing the most precise and versatile tool in the genome-editing toolkit.
CRISPR-MEDIATED GENOME EDITING
Cas9 derived from Streptococcus pyogenes (SpCas9) cleaves double-stranded DNA, producing blunt-end DSBs, which are subsequently repaired through intrinsic cellular mechanisms, including nonhomologous end joining (NHEJ) and homology-directed repair (HDR) (Fig. 1). NHEJ, the predominant repair pathway, often introduces random insertions or deletions (indels) at the break site, leading to mutations. This approach has been widely used to disrupt gene function or facilitate targeted gene insertions. 10,11 In contrast, HDR is a high-fidelity process that can precisely repair genomic sequences when a homologous donor template is available, making it particularly appealing for clinical applications. To enhance the versatility of CRISPR-based genome editing, various Cas9 orthologs have been explored for their editing capabilities. 12 –18 For instance, Cas9 from Staphylococcus aureus (SaCas9) exhibits genome-editing efficiency comparable to SpCas9 but is significantly shorter (>1 kb smaller). This compact size allows SaCas9 and its single guide RNA (sgRNA) expression cassette to be packaged into a single adeno-associated virus (AAV) vector, facilitating its use in therapeutic applications. 12,19 These advancements have significantly expanded the CRISPR toolkit, enabling more precise and adaptable genome-editing strategies.
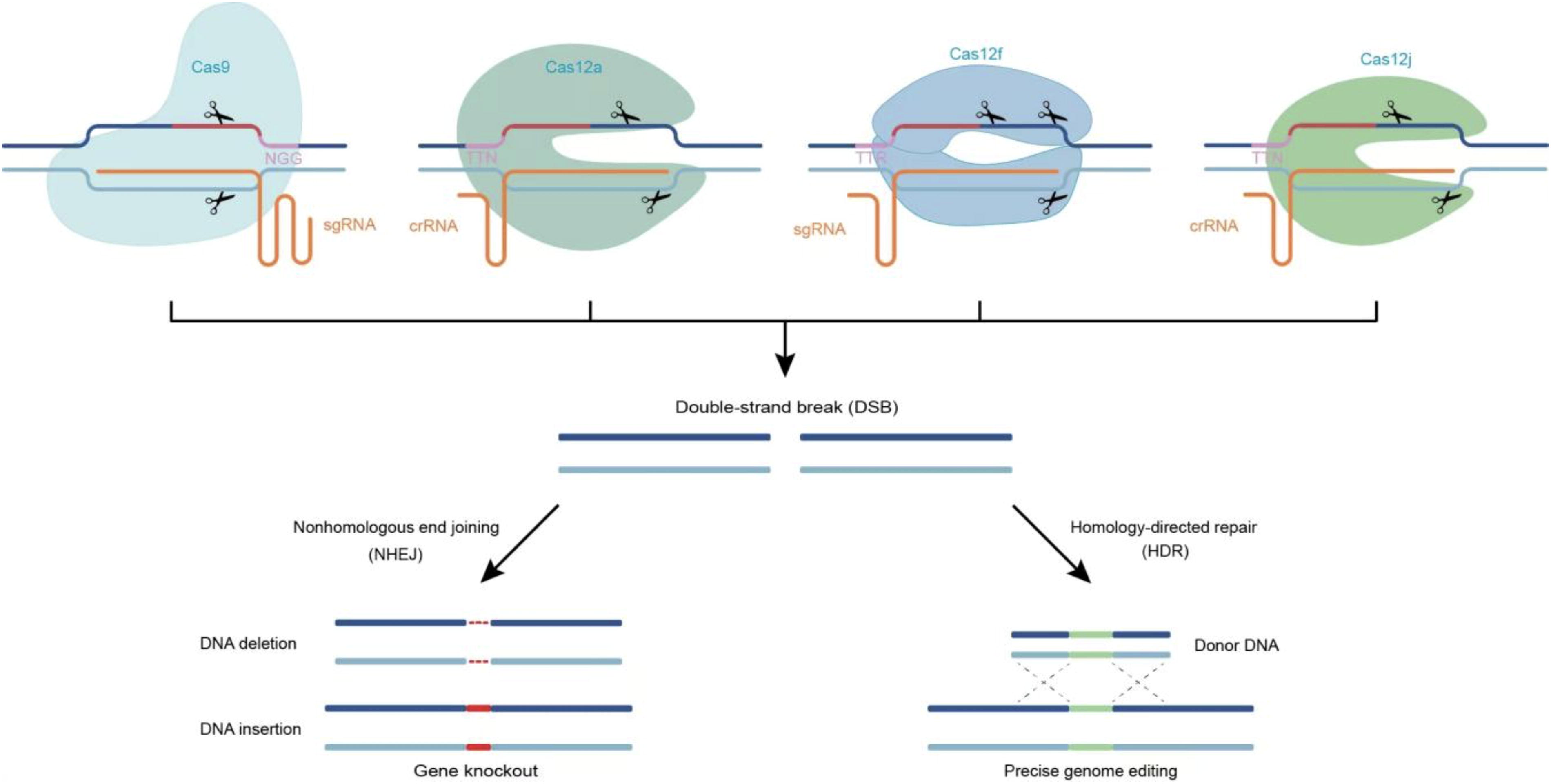
The clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated system (Cas)-based genome editing. The CRISPR/Cas9 system is composed of Cas9 and an sgRNA. The CRISPR/Cas12a system is composed of Cas12a and a crRNA. The CRISPR/Cas12f system is composed of Cas12f and an sgRNA. The spacer sequence, which is complementary to the target DNA, contains a seed region (red line) that affects the specificity of CRISPR/Cas. Guided by the spacer sequence, the Cas endonuclease cleaves DNA, generating double-strand breaks (DSBs). The repair of DSBs can occur via nonhomologous end joining (NHEJ) or homology-directed repair (HDR). NHEJ often results in insertions or deletions (indels), while HDR enables precise genome editing using a donor template.
The CRISPR-Cas12 family comprises a diverse array of single effector enzymes, including Cas12a, Cas12b, Cas12c, Cas12d, Cas12e, and Cas12f. Studies have demonstrated that Cas12b, Cas12c, Cas12d, and Cas12e exhibit minimal off-target effects during targeted genome modification. Among them, Cas12b (C2c1) demonstrates high specificity and stability across a wide temperature range, making it suitable for use in mammalian cells and thermophilic organisms. 20 Cas12c possesses unique RNA-guided DNA targeting capabilities and holds potential value in multiplex genome editing. 21 Cas12d (CasY), originating from uncultured archaea, can be engineered to enable efficient editing in human cells. Cas12e (CasX) has an extremely small protein size, which facilitates its delivery and packaging for in vivo applications. 22,23 Cas12a and Cas12f have been extensively studied due to their small size and high editing efficiency.
Cas12a, also known as Cpf1, is a single RNA-guided endonuclease belonging to the Class 2 Type V-A CRISPR system. Guided by a single crRNA, it recognizes T-rich PAM sequences (such as TTTV) and generates DSBs with 4–5 nucleotide overhangs. Unlike Cas9, Cas12a cleaves DNA distal to the PAM site, creating sticky ends. It cuts the targeted DNA distal to the thymine (T)-rich protospacer adjacent motif (PAM), generating DSBs with 4 to 5 nucleotide overhangs (Fig. 1), which is conducive to achieving precise genome editing. Cas12a is widely used in multiplex editing based on homology-directed repair (HDR) and shows potential in clinical applications due to its high specificity and efficiency. 24
Cas12f, also known as Cas14, is a small RNA-guided endonuclease in the Class 2 Type V-F CRISPR system. With a protein size of only 422 amino acids, it is one of the smallest known CRISPR endonucleases. Cas12f cleaves DNA outside the protospacer, creating sticky ends (Fig. 1), enabling efficient genome editing. Due to its miniaturized nature, Cas12f can be packaged into a single AAV vector, making it highly suitable for in vivo gene therapy. Cas12f has demonstrated efficient editing capabilities in mammalian cells and animal models, providing a promising platform for therapeutic genome editing. 25
CRISPR-BASED PRECISE GENOME EDITING
Single-nucleotide variations are responsible for two-thirds of human genetic diseases. Thus, to comprehensively cure a disease one must correct, rather than disrupt, point mutations in target genes. Although a variant could theoretically be repaired through HDR, the efficiency is low in most cells. Moreover, DSBs generated by nucleases can cause large deletions, chromosomal translocations, and chromothripsis. 26 –28 Therefore, it is important to develop efficient and precise genome-editing tools that do not require DSBs. With this in mind, a nascent technology called base editing was developed in 2016. 29
Base editors enable the precise installation of targeted point mutations without inducing DSBs or the need for donor DNA. 29 –31 These editors typically consist of a catalytically impaired Cas protein, Cas9n(D10A), a single-stranded DNA deaminase, and a single guide RNA (sgRNA). Guided by the sgRNA, Cas9n(D10A) localizes the deaminase to the target site, and the targeted nucleotide within the activity window is deaminated and converted to another nucleotide in a programmable manner. To date, three classes of base editors have been developed: the cytosine base editor (CBE), adenine base editor (ABE), and cytosine transversion base editor (GBE).
Using a CBE, cytosines within the target sequence act as the substrates for cytidine deaminase, which catalyzes the hydrolytic deamination of cytosine to uracil. With the help of uracil glycosylase inhibitor domains (UGI), which inhibit the activity of uracil glycosylases, the CBE installs C-to-T substitutions in the target DNA (Fig. 2A). Using an ABE, adenines within the target sequence are the substrates for adenine deaminase, which catalyzes the hydrolytic deamination of adenosine to inosine. Although there are no enzymes known to deaminate adenine in DNA, a deoxyadenosine deaminase (TadA*) that accepts DNA as its substrate has been developed. An ABE installs A-to-G substitutions in the target DNA (Fig. 2B). Using a GBE, cytosines within the target sequence are the substrates for cytidine deaminase, which catalyzes the hydrolytic deamination of cytosine to uracil. With the help of uracil DNA glycosylase, which removes U and creates an apurinic/apyrimidinic (AP) site, a GBE installs C-to-G substitutions in the target DNA (Fig. 2C). Base editors have been used to install or reverse point mutations in various cell types and organisms. 7,32,33 and they have been used for disease treatment. 34
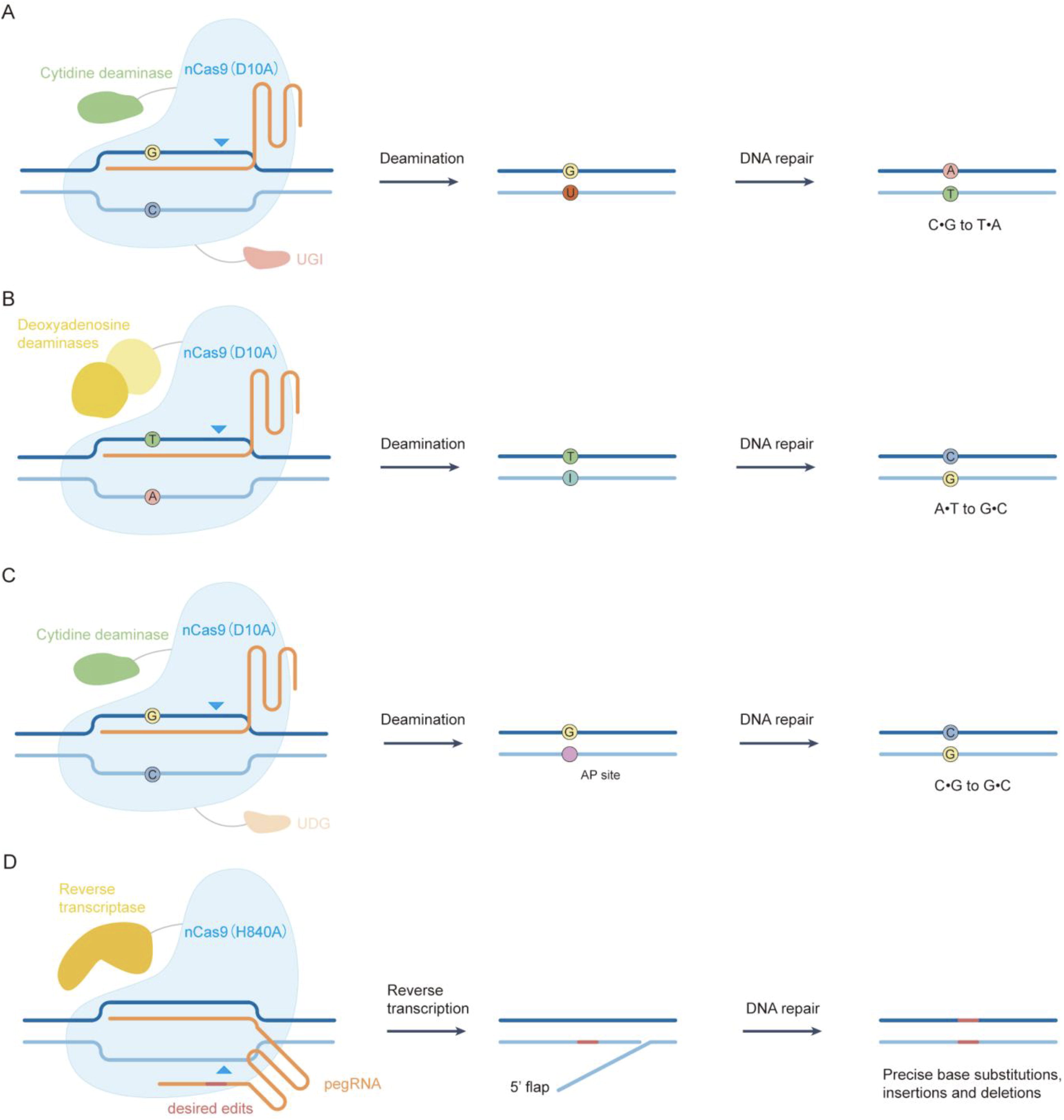
CRISPR-mediated precise genome editing.
Beyond the three established classes of base editors, recent studies have introduced glycosylase-based editors, including gGBE, AYBE, gCBE, and gTBE. These innovative tools harness the unique properties of glycosylases to enable precise base editing without inducing DSBs. For example, gGBE achieves C-to-G transversion by excising uracil residues 35 ; AYBE facilitates A-to-Y editing, thereby expanding the repertoire of therapeutic tools; 36 gCBE integrates cytidine deaminase with glycosylase activity to achieve C-to-T conversion across a wider range of genomic contexts, overcoming limitations of conventional CBEs; and gTBE employs thymine DNA glycosylase (TDG) to excise thymine, enabling T-to-C conversion through endogenous repair mechanisms. 37 These advancements significantly enhance the precision and versatility of base editing technologies, broadening the scope of nucleotide conversions and offering promising new avenues for correcting disease-associated point mutations in genetic disorders.
Transversion mutations, targeted deletions (e.g., removal of the 4-bp duplication that causes Tay-Sachs disease), and targeted insertions (e.g., a 3-bp insertion that corrects the most common cause of cystic fibrosis) are difficult to install or correct efficiently. Recently, a revolutionary technology called prime editing that can overcome these problems was developed in 2019. A prime editor contains a catalytically impaired Cas, Cas9n(H840A), fused to an engineered reverse transcriptase, and a prime editing guide RNA (pegRNA), which specifies the target site and encodes the desired edit. Guided by the pegRNA, Cas9n(H840A) localizes reverse transcriptase to the target site and the primer-binding site pairs with the Cas9n(H840A)-nicked strand, priming reverse transcription and incorporating the desired edits into the genome. Therefore, prime editing writes new genetic information directly into the targeted DNA (including all 12 types of point mutations, targeted insertions, targeted deletions, and combinations of these edits) in human cells without DSBs or donor DNA. 38,39 In principle, prime editing could correct about 89% of known pathogenic human genetic variants (Fig. 2D).
SPECIFICITY OF CRISPR AND CRISPR-BASED TECHNOLOGIES
CRISPR and CRISPR-based technologies are RNA-guided genome editors. Cas nucleases can tolerate mismatches between the RNA and target DNA; thus, unwanted off-target effects are possible. For instance, sgRNA-dependent off-target edits can be induced by a tolerance for mismatched nucleotides in target-like sequences in human cells. 40 As with CRISPR/Cas genome-editing technologies, base editors have the potential to introduce sgRNA-dependent off-target effects; moreover, they can induce sgRNA-independent off-target effects. For example, a CBE can induce genome-wide off-target effects via the long-term expression of cytidine deaminase. 41 Recently, it was confirmed that CBEs and ABEs cause transcriptome-wide off-target effects by editing RNA transcripts. 42,43 Unlike Cas nucleases and base editing, prime editing utilizes three checkpoints for complementary base pairing; therefore, off-target genomic changes are rare. 44,45
Reducing the off-target effects of genome-editing tools is of utmost importance for gene therapy. Fortunately, off-target effects can be mitigated by selecting highly specific target sites and by using truncated sgRNAs, RNPs, or rationally engineered deaminase variants. 46,47 Moreover, through phage-assisted evolution and structure-guided engineering, researchers have developed several high-fidelity Cas variants with reduced off-target editing activity. 48 –55 For example, SpCas9-HF1 is a high-fidelity variant that harbors alterations designed to reduce nonspecific DNA contacts. 49 However, these variants exhibit reduced on-target activity as well. Therefore, identifying CRISPR systems with robust on-target efficiency and low off-target activity continues to be an important topic.
APPLICATIONS OF CRISPR IN GENE THERAPY
CRISPR and CRISPR-based genome-editing technologies enable precise modifications of complex genomes. Since its adaptation for use in mammalian cells, CRISPR has emerged as a transformative tool, offering unprecedented opportunities for the treatment of various genetic disorders. 56 For diseases such as specific cancers and blood disorders, an ex vivo approach is appealing given that blood cells are good targets for ex vivo editing. 2,57 In contrast, for diseases such as Leber congenital amaurosis (LCA) direct in vivo editing makes sense because the target tissue is accessible. 13 To date, most clinical trials have relied on ex vivo editing using CRISPR, with the subsequent infusion of the modified cells into patients (Fig. 3). These approaches have proven to be highly effective. In the following section, we summarize attempts to treat different diseases with CRISPR gene-editing tools (Table 1).

CRISPR therapy may be done ex vivo or in vivo. The ex vivo approach involves the extraction of target cells, delivery of the CRISPR components to yield gene-modified cells, expansion of the edited cells, and their infusion into patients. In the in vivo strategy, CRISPR components can be locally delivered to the target tissue. Subsequent editing of the target genes produces edited cells in vivo to provide a therapeutic benefit.
Applications of CRISPR in Cancer, Sickle Cell Disease, β-Thalassemia, In Vivo Disease Treatment, Genetic Diseases, and Immune Diseases
Cas, CRISPR-associated system; CRISPR, clustered regularly interspaced short palindromic repeats; CBE, Cytosine base editing; CF, Cystic Fibrosis; HBB, hemoglobin β subunit gene; SCD, Sickle Cell Disease; SCID, Severe Combined Immunodeficiency; TDT, β-Thalassemia; TTR, Transthyretin.
ATTEMPTS TO TREAT CANCER
Cancer, a disease with high incidence and mortality rates, is a public problem that seriously threatens human life and health. Presently, surgery, radiotherapy, and chemotherapy are the main approaches for treating cancer. 57 However, the latter two treatments engender serious side effects. 58 Chimeric antigen receptor (CAR)-T cell therapy can have remarkable effects on patients with refractory or relapsed malignancies; 2,57 thus, it has become the fourth major treatment modality. The development of genome-editing technologies has opened a window to accelerate the production of the next generation of CAR-T cells.
T cells play a significant role in the therapeutic effect of CAR-T cells. However, T cells are continually exposed to antigens in patients with malignant tumors, resulting in the expression of multiple inhibitory receptors, including PD-1, CTLA-4, and LAG3. These receptors inhibit T cell proliferation and cytokine production, leading to immune escape. Thus, the disruption of these inhibitory receptors could improve the potency of CAR-T cells. One study reported that anti-CD19 CAR-T cells with a CRISPR-mediated double (TRAC/B2M) or triple (TRAC/B2M/PD-1) knockout had increased antitumor functions in vitro. 59 Another study involving an EGFRvIII CAR-T cell product with a CRISPR-mediated triple (TRAC/B2M/PD-1) knockout demonstrated enhanced antitumor activity in a preclinical glioma model, 60 warranting further investigation in patients with glioblastoma.
Rapid cell proliferation is a distinct feature of cancer development. Although photothermal therapy is used to ablate tumors, it is usually accompanied by undesirable damage to healthy organs due to the high temperatures that are used (tumors can tolerate temperatures of up to 50°C). Researchers have designed an elegant strategy utilizing CRISPR/Cas9 technology for tumor-targeted gene editing. 3 They constructed a hypoxia-responsive gold nanorod-based nanocomposite of CRISPR/Cas9 to knock out HSP90a and reduce the thermal resistance of cancer cells. Following the entry of the complex into tumor cells, CRISPR/Cas9 RNP was released and HSP90a was knocked out; effective tumor ablation was achieved under mild photothermal therapy both in vitro and in vivo.
The first in-human clinical trial of CRISPR/Cas9 was conducted by Lu et al. 61 at West China Hospital to treat lung cancer in 2016. Using electroporation, Cas9 and sgRNA plasmids were cotransfected into T cells derived from patients, generating PD-1-edited T cells. Then, they were expanded ex vivo and injected back into the patients. Edited T cells were detected in peripheral blood samples after the infusion. Further data showed that PD-1 expression was significantly reduced in the gene-edited T cells. Follow-up studies showed that none of the patients had treatment-related severe adverse effects. Thus, this study demonstrates the safety and feasibility of CRISPR-mediated gene therapy.
The first in-human phase 1 clinical trial of CRISPR in the United States opened in 2018 with the intent to edit T cells for cancer immunotherapy against melanoma, synovial sarcoma, and multiple myeloma. 59 T cells derived from patients were engineered with CRISPR/Cas9 to knock out genes encoding the endogenous T cell receptor (TCR) chains TCRα (TRAC) and TCRβ (TRBC), as well as PD-1. Then, the cells were transduced with lentivirus particles carrying a gene encoding a TCR specific for NY-ESO-1 antigen. 60 Following gene editing and transduction, the NY-ESO-1-redirected CRISPR-edited T cells were expanded ex vivo and injected back into the patients. The edited T cells persisted for up to 9 months, and there were no clinical toxicities. These results demonstrate the safety and feasibility of multiplex editing using CRISPR/Cas9 in human T cells.
The first off-the-shelf CAR-T cell therapy with a PD-1 knockout was conducted by Caribou Bioscience 61 to treat aggressive non-Hodgkin’s lymphoma. They used CRISPR/Cas9 to insert a CD19-specific CAR into the targeted TRAC gene and knock out PD-1 in human primary T cells derived from healthy people. These modified CAR-T cells, which exhibited uniform CAR expression and enhanced tumor rejection, 62 were expanded ex vivo. No graft-versus-host disease was detected in patients that received 40 × 106 of the CAR-T cells. Therefore, this strategy may overcome durability concerns and help create therapies that are as effective as autologous anti-CD19 CAR-T cell therapies but which are much less expensive.
CRISPR-mediated genome editing in human cells is not without risk. Cas9 induces off-target cleavage, and chromosomal translocations have been detected in edited T cells. 63 Base editing, which creates premature stop codons without inducing DSBs, has greater potential in gene therapy than Cas9. Moreover, it has been investigated in patients with T-cell acute lymphoblastic leukemia (T-ALL). Researchers used a CBE to introduce stop codons to inactive three genes (TRBC, CD7, and CD52) in healthy volunteer donor T cells. Then, the cells were transduced with lentivirus particles carrying a gene encoding a chimeric antigen receptor that recognizes CD7 (CAR7). The base-edited CAR7 cells were infused into patients. It worked in a 13-year-old girl who experienced successful immunological reconstitution and ongoing leukemic remission. 34 A similar study using a CBE to produce multiplex base-edited T-cells for T-ALL was done in the United States. 4 These findings underscore the potential of base editing to advance immunotherapy development.
To date, many trials have used CRISPR to engineer immunotherapies for cancer. 62 The strategies used in these trials involved the inactivation of immunosuppressive factors by CRISPR or the integration of a CAR element into a specific locus for CAR-T cell engineering. Both strategies have been shown to promote the effectiveness of tumor-specific cytotoxic T cells; thus, they both hold promise in cancer treatment. 64,65
ATTEMPTS TO TREAT SICKLE CELL DISEASE (SCD) AND β-THALASSEMIA (TDT)
SCD and transfusion-dependent TDT are severe monogenic diseases caused by mutations in the hemoglobin β subunit gene (HBB). 66,67 Patients with SCD are usually treated with blood transfusions and iron chelation while patients with TDT are usually treated with hydroxyurea, pain relievers, and blood transfusions. However, neither treatment fully ameliorates the manifestations of the disease or addresses the underlying causes. Allogeneic bone marrow transplantation has been used to cure both SCD and TDT, but matching is difficult. 68 BCL11A is a transcription factor that represses the expression of fetal hemoglobin and γ-globin. 69 Individuals whose fetal globin is not silenced following birth are largely protected from SCD and TDT. 70 Therefore, disrupting the activity of BCL11A would increase γ-globin expression and alleviate the symptoms of SCD and TDT. CRISPR-based BCL11A editing offers hope for a cure for SCD and TDT.
Researchers have used CRISPR/Cas9 to edit the BCL11A enhancer in hematopoietic stem cells. The expression of BCL11A was reduced and fetal γ-globin was induced without significant side effects. 71 Subsequently, a clinical trial aimed at treating SCD and TDT was designed by CRISPR Therapeutics in association with Vertex Pharmaceuticals. 69 They used CRISPR/Cas9 to edit the BCL11A enhancer in CD34+ hematopoietic stem and progenitor cells (HSPCs) obtained from healthy donors. The editing efficiency was approximately 80% with no evidence of off-target editing. Subsequently, two patients, one with SCD and one with TDT, were infused with autologous CD34+-edited HSPCs. More than a year later, both patients had substantial and sustained increases in fetal hemoglobin with >99% pancellularity. The treatment was not harmless; adverse events were reported, but they were not life-threatening and resolved after treatment. In an additional eight patients infused with autologous CD34+-edited HSPCs, similar results to the first two patients were observed. Excitingly, the first regulatory authorization of the CRISPR/Cas9-based gene-edited therapy in the world, CASGEVYTM was announced for the treatment of SCD and TDT in November, 2023.
In another clinical study for treating SCD, a strategy using RNAi to knock down BCL11A was used and clinical success was achieved. 72 Recently, a clinical trial for patients with TDT was designed in China. 73 In the trial, BCL11A-edited autologous HSPCs were transplanted into two children, one with the most severe (β0/β0) form of TDT. Both patients were clinically well with multilineage engraftment, showed robust γ-globin expression, and achieved transfusion independence for >18 months after treatment. These results suggest that CRISPR/Cas9-based BCL11A editing has great potential to address the underlying causes of SCD and TDT.
SCD is caused by an A·T-to-T·A transversion mutation in HBB. Researchers corrected the SCD allele (HBBS ) to the wild-type allele (HBBA ) in HSPCs from patients using prime editing and subsequently transplanted the prime-edited SCD HSPCs into immunodeficient mice. After 17 weeks, the mice had measurable HBBA levels and displayed engraftment frequencies, hematopoietic differentiation, and lineage maturation similar to those of unedited HSPCs from healthy donors. 74 These results demonstrate a potential strategy for a one-time autologous SCD treatment that directly corrects HBBS back to wild-type HBBA without requiring DSBs or donor templates.
ATTEMPTS TO TREAT DISEASES IN VIVO
LCA type 10 (LCA10) is a debilitating retinal dystrophy caused by bi-allelic loss-of-function mutations in CEP290. Researchers developed an SaCas9-edited therapy (EDIT-101) to treat LCA10. 19 They identified a pair of sgRNAs with high efficiency and specificity to remove the CEP290 mutant intron using SaCas9. Then, they used an AAV5 vector to deliver the sgRNAs and SaCas9 into photoreceptor cells by subretinal injection to restore normal CEP290 expression. Rapid and sustained CEP290 gene editing was observed; the CEP290 expression level met the target therapeutic threshold. The first clinical use of EDIT-101 for the treatment of patients with LCA10 was done in 2020. This clinical trial is the world’s first in vivo use of CRISPR in patients. After 6 months, the patients in the mid-dose group experienced a pronounced therapeutic effect while those in the low-dose group experienced a poor therapeutic effect. Fortunately, none of the patients experienced any serious adverse effects. The safety, tolerability, and efficacy of EDIT-101 will be further assessed in approximately 18 patients.
Transthyretin (TTR) amyloidosis is an autosomal dominant disorder characterized by the accumulation of misfolded TTR in tissues—predominantly the nerves and heart. The clinical phenotypes of most patients are amyloid polyneuropathy and cardiomyopathy. Considering that TTR is synthesized almost entirely within the liver and that low expression has no significant side effects, eliminating the expression of mutated TTR is a therapeutic strategy for TTR amyloidosis. 75 Researchers designed an in vivo gene-editing therapy named NTLA-2001. 76 It utilized a lipid nanoparticle delivery system with liver tropism, which carried mRNA encoding SpCas9 and an sgRNA targeting TTR. After intravenous infusion, only mild adverse events were observed in three of the six patients. By day 28, treatment was associated with mean TTR reductions of 52% in the group that received a low dose (0.1 mg/kg) and 87% in the group that received a high dose (0.3 mg/kg). This finding indicates that the effects of NTLA-2001 were dose-dependent, and they underscore the potential for in vivo gene editing as a therapeutic strategy for monogenic diseases.
ATTEMPTS TO TREAT GENETIC DISEASES AND IMMUNE DISEASES
Cystic fibrosis is a severe genetic disorder. Mutations in the cystic fibrosis transmembrane conductance regulator (CFTR) gene lead to abnormal ion channel function, resulting in abnormal mucus secretion in multiple organs such as the lungs and pancreas. This, in turn, triggers a series of severe symptoms including breathing difficulties, recurrent pulmonary infections, and digestive dysfunctions. The CRISPR technology, with its ability to precisely locate and cleave specific DNA sequences, can be used to edit the mutated CFTR gene. Some studies on cell models and animal experiments have yielded encouraging results. Researchers have designed guide RNAs complementary to the mutated gene sequences, which direct Cas nucleases to accurately reach the mutation sites. By repairing the mutated bases, the normal function of the CFTR gene is restored. 77 –79 This lays a solid foundation for the future realization of effective treatments for cystic fibrosis in human clinical trials.
Severe combined immunodeficiency (SCID) is a group of extremely severe hereditary immune disorders. Patients with SCID have multiple defects in their immune systems, rendering them almost defenseless against various pathogens, which seriously threatens their lives and health. The IL2RG gene plays a crucial role in the normal development and function maintenance of the immune system. Mutations in this gene can lead to abnormalities in the development, differentiation, and functional performance of immune cells such as T cells and B cells, thus triggering SCID. The CRISPR technology, with its unique gene-editing capabilities, can precisely identify mutation sites on the IL2RG gene. The designed guide RNA (gRNA) directs the Cas nuclease to reach the mutation site accurately, enabling in-situ editing of the mutated gene directly and achieving precise repair of the pathogenic mutations. 80
CHALLENGES AND PERSPECTIVES ON THE USE OF CRISPR IN GENE THERAPY
Over the past decade, the development of CRISPR-based genome-editing tools has expanded significantly, accompanied by a deepening understanding of their diverse applications. Generally, CRISPRs are comprised of a large Cas protein and an sgRNA; these components (e.g., DNA, RNA, or RNP complexes) should be delivered into targeted cells or tissues for successful therapeutic genome editing. Presently, CRISPR/Cas9 is the most frequently used genome-editing tool for the treatment of genetic disorders. Cas9 is initially guided to the coding sequence of a deleterious protein; it then creates DSBs and induces a frameshift to silence protein expression. However, DSBs are usually associated with undesired outcomes, 26 –28 and most diseases are complex and cannot be cured via the targeted knockout of a deleterious variant. Therefore, genome editing technologies (e.g., base editing, prime editing, and epigenetic modification) that can mediate genome editing without requiring DSBs have been explored. For many genetic diseases, multiple genome-editing strategies may be possible, suitable, safe, and efficient and should be considered as the optimal treatment approach based on the characteristics of the disease being treated.
CRISPR-based genome-editing technologies offer unparalleled opportunities to treat specific cancers and blood disorders via an ex vivo approach. 2,57 In comparison, in vivo approaches have been less extensively employed and are largely limited by inadequate accessibility to the target tissue. For gene therapy, an AAV is the leading tool for delivering CRISPR. However, the packaging capacity of an AAV (∼4.7 kb) is limited. Small Cas orthologs have been explored for genome editing to overcome this problem. Moreover, dual AAV vectors and bioengineered delivery approaches (e.g., using polymers, nanoparticles, and modified viral platforms) have been investigated to improve the efficacy and safety of delivery vehicles. 81,82 Although these possibilities exist, off-target effects increase with increasing Cas9 activity. Unintended edits are one of the most important issues in CRISPR gene therapy. Therefore, researchers have developed several strategies to improve the specificity of CRISPR, including bioinformatic analyses of targets, the engineering of high-fidelity Cas variants, and the transient expression of editing reagents. 6 Cas9 mRNA-lipid nanoparticles have also been used in gene therapy, and they have proven to be effective in preclinical studies. 83,84 Additionally, emerging strategies using genetic regulation (i.e., chemical and physical strategies to spatiotemporally control CRISPR activity) could reduce undesirable edits and broaden the clinical applications of CRISPR. 74 Continued advancement of delivery methods to target tissues and improvements in the fidelity and specificity of CRISPR will fuel the development of clinical trials in the future.
Finally, safety and ethical concerns related to the use of CRISPR systems in gene therapy must be considered before they can be used clinically. With somatic editing, any potential risk would be contained within the individual after partaking in the therapy; therefore, somatic editing using CRISPR therapy has been permitted and applied after careful consideration. If with embryonic editing, the genetic information of the individual is modified, spread to all cells, and will be subsequently passed down through generations. This may lead to irreversible mutations and have serious consequences for future generations; long-term safety is impossible to predict. However, CRISPR-based gene editing of human embryos with the intention to confer HIV resistance was reported in 2018. A criminal case, the first involving genome-edited babies, ensued in 2019 and resulted in a three-year prison sentence for the lead scientist. For now, research on gene editing in human embryos is banned.
CONCLUSION
Careful consideration and study of above challenges and issues will promote the development of novel CRISPR-based technologies for disease treatment. Vertex and CRISPR Therapeutics announced authorization of CASGEVYTM, the first CRISPR-based gene-edited therapy has received regulatory approval for the innovative treatment. This not only opens a new chapter in personalized medicine, but also heralds its potential to transform the lives of millions of patients around the world. We anticipate that CRISPR-based tools, together with a greater understanding of disease mechanisms and other nascent technologies, will yield significant advances in gene therapy for cancer and sickle cell disease.
AUTHORS’ CONTRIBUTIONS
S.S.Z. and J.L.: Wrote the article. J.R.Z. and R.R.Z.: Created the figures and tables, while X.Z., J.R.Z., and R.R.Z.: Provided critical edits.
Footnotes
AUTHOR DISCLOSURE STATEMENT
The authors declare no competing interests.
FUNDING INFORMATION
No funding was received for this article.