
Abstract
Sensorineural hearing loss (SNHL) is the most common sensory deficit globally. Acquired SNHL results from ototoxic damage to cochlear hair cells (HCs) and is typically irreversible due to their limited regenerative capacity. While no cure currently exists, targeting the underlying pathology offers potential. Preclinical studies have investigated transcription factors like ATOH1, which can induce non-sensory cells to transdifferentiate into HCs. Gene therapy using viral vectors to deliver ATOH1 is emerging as a promising regenerative approach. PubMed, Web of Science, and Embase were systematically searched. The review was conducted following the Systematic Review Center for Laboratory Animal Experimentation guidelines. Random-effects meta-analysis was conducted using R’s “meta” and “metafor” packages. To corroborate our findings, differential gene expression (DEG) analysis was performed on the GEO dataset GSE127683 using DESeq2. K-means clustering and gene set enrichment analysis (GSEA) were conducted using iDEP 2.0 and Enrichr, respectively. Four studies including 52 rodents were included. ATOH1 gene therapy significantly reduced Auditory Brainstem Response thresholds (MD = −21.37 dB SPL, CI: [−40.19; −2.54], p = 0.027), indicating improved hearing. DEG analysis showed upregulation of genes crucial for hair cell differentiation and functioning, including GFI1, PTPRQ, OTOF, USH2A, and POU4F3. GSEA highlighted key upregulated pathways related to inner ear development, auditory receptor cell differentiation and sensory perception of sound. ATOH1 gene therapy shows promise for treating acquired SNHL. However, further clinical trials are essential to confirm these preclinical findings and advance towards a potential cure.
INTRODUCTION
Hearing loss is the most common sensory deficit, affecting over 466 million people globally. 1 It presents in two forms: conductive hearing loss, caused by issues with sound transmission, and sensorineural hearing loss (SNHL), the more prevalent form, which results from neurodegenerative changes in sensory hair cells, their synapses with the cochlear nerve, and the cochlear nerve itself. 2 Acquired SNHL is primarily caused by aging, noise exposure, and ototoxic drugs, which damage key inner ear structures like hair and supporting cells, disrupting their function and leading to hearing loss. 3 A complex intercalation of genetic and environmental factors contributes greatly to the pathology, 4 which in combination with the inner ear hair cells limited or even absent capacity for regeneration 3 has rendered SNHL an incurable disease where a satisfactory therapy has yet to be discovered and implemented. 4
For those experiencing mild to moderate hearing loss, hearing aids are an appropriate choice. In cases of severe to profound loss, cochlear implants are typically used. These implants, however, can be expensive, often necessitate surgical intervention, and may not always yield satisfactory results for all patients. Drug therapies, including steroids and neurotrophic factors, remain unproven due to a lack of randomized, double-blind, case-controlled studies to confirm their benefit. Meanwhile, novel therapies focused on hair cell regeneration, particularly stem cell and gene therapy, offer a new horizon of potentially curative treatments. 4
Treatment strategies focused on genetic intervention aim to enhance hair cell survival or induce transdifferentiation from progenitor cells via three key mechanisms: replacement, silencing, or editing of defective genes. 4 These therapies are generally delivered into the inner ear using viral vectors, such as retroviruses (including lentivirus), and more commonly, adenoviruses (AdVs) and adeno-associated viruses (AAVs), both of which exhibit high transduction efficiency and tropism for various cochlear cell types. 1
Gene therapy targeting ATOH1 represents a promising avenue of research for a potential SNHL cure. The ATOH1 gene, located on chromosome 4q22.2, encodes a basic helix-loop-helix transcription factor crucial for hair cell differentiation during inner ear morphogenesis. 5 ATOH1 regulates downstream genes Pou4f3 and Gfi1, which are essential for the differentiation and maturation of progenitors into hair cells. It also targets Barhl1, a transcription factor strongly expressed in cochlear hair cells and vital for their long-term maintenance. 6 ATOH1 upregulation leads to the formation of additional or ectopic hair cells, while its absence culminates in the failure of vestibular and cochlear hair cell formation. 7 Moreover, differentially timed expression patterns of ATOH1 aligns with the period of hair cell differentiation. During early embryonic development it is expressed in differentiating hair cells but is downregulated in supporting cells and even absent outside the sensory epithelium during later stages. Collectively, this makes ATOH1 a prime candidate for manipulating and controlling hair cell differentiation. 5
The literature on ATOH1 gene therapy for SNHL presents conflicting results. Some studies report that delivering ATOH1 into the inner ear using adenoviral or adeno-associated vectors significantly improves hearing thresholds in deaf mice, 8 successfully transducing new hair cells and hair cell-like cells. These studies demonstrate robust hair cell regeneration and effective transdifferentiation into hair cells when ATOH1 is delivered via viral vectors in SNHL animal models. 9,10 However, opposing studies show that while AdV-mediated ATOH1 therapy promotes the differentiation of non-sensory cells into immature sensory cells, increasing hair cell numbers, it fails to improve function in aminoglycoside-deafened guinea pigs, with no significant post-treatment auditory brainstem response (ABR) threshold or auditory neuron survival improvement. 11
The inconsistency in the literature highlights the need for a comprehensive synthesis to determine the efficacy of ATOH1 gene therapy via viral vectors for acquired SNHL. This need is emphasized by the lack of human clinical trials, with the first expected to conclude by 2027. 12 This systematic review and meta-analysis aims to assess whether this therapy is effective and worth further investment. To our knowledge, it is the first meta-analysis to incorporate extensive bioinformatic analysis of the ATOH1 gene, bolstering the integrity of our findings and offering an integrated understanding of its potential and limitations. By translating preclinical findings to clinical application, this research could revolutionize SNHL management and reduce its global burden.
METHODS
Protocol and registration
Systematic Review Center for Laboratory Animal Experimentation (SYRCLE) guidelines were followed to minimize bias in preclinical study selection and assessment. 13 The protocol was prospectively registered in PROSPERO – International Prospective Register of Systematic Reviews (CRD42024560874), and a detailed description of the methods is available in the published protocol. The reporting of this systematic review and meta-analysis adheres to Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines to ensure clarity, completeness, and methodological transparency in presenting the findings (Supplementary Table S1).
Search strategy and study selection
A systematic literature search was conducted on PubMed, Web of Science, and Embase. Keywords included “acquired sensorineural hearing loss,” “deafness,” “ATOH1,” “gene therapy,” “preclinical studies,” “hair cell renewal,” and “auditory function improvement” from inception to April, 2024. An updated search covering the period from March 2024 to March 2025 identified no additional eligible studies. Full search strings are in the Supplementary Table S2. Two independent investigators (Z.A.-S. and N.A.-A.) screened articles using Rayyan.ai’s blinded screening. 14 Titles and abstracts were first reviewed, followed by full-text screening based on inclusion and exclusion criteria. Disagreements were resolved through a third investigator.
Eligibility criteria
Inclusion criteria included: (1) studies originally in English language, (2) preclinical adult animal models (to mimic human hearing loss scenarios), (3) acquired SNHL (ototoxic drugs, noise-induced or age-related hearing loss, etc.), (4) ATOH1 gene therapy, (5) viral vector delivery within 10 days of deafness induction), and (6) quantitative auditory function reporting. No date restrictions were applied.
Exclusion criteria were: (1) non-English publications, (2) non-primary studies (reviews, editorials), (3) incorrect study design (including in vitro/in silico studies), (4) incorrect intervention (non-ATOH1 or combinatorial gene therapy strategies), (5) no control group, (6) genetic or congenital hearing loss models (e.g., Prestin knockout, Usher Syndrome), (7) non-viral or non-ATOH1 gene therapy, (8) no ABR thresholds, and (9) qualitative auditory function reporting only.
Quality assessment
Two investigators (M.A.-T. and F.Q.) independently assessed study quality using SYRCLE’s risk of bias (RoB) tool, evaluating 10 domains across six bias types: selection, performance, detection, attrition, reporting, and other biases. Each entry was judged as “yes” (low risk), “no” (high risk), or “unclear” (insufficient detail). 13 Disagreements were resolved by consensus, with a third investigator consulted if necessary.
Study characteristics and data extraction
Our primary outcome is the ABR threshold, a functional hearing test that reflects neural activity along the auditory pathway in response to auditory stimuli. 15 Data extraction was conducted by (F.Q.) and reviewed by (M.A.-T.). Extracted data included bibliographic details (authors, year), experimental study design (study type, deafness induction method, hearing tests, treatment timing, and post-treatment assessment period), animal model characteristics (species/strain, age, number), and intervention details (viral vector type, serotype, gene delivered, dose, injection route). Additional methodological parameters were also extracted, including the promoter used, post-delivery wound sealing method, and whether functional and structural regeneration was reported. ABR threshold values (dB SPL) were extracted from graphical data using plot digitizer software, 16 as no numerical/tabular data was available. Missing variance measures (standard deviation [SD], scanning electron-micrography [SEM]) were imputed by either averaging available variances for the same frequency or, if no frequency-specific variances were available, averaging all variance measurements. 17 Where any data or methodological detail was not reported in the original publications, corresponding authors were contacted.
Meta analysis
A meta-analysis was conducted to compare ABR threshold differences between ATOH1-treated and control animals.
To ensure the independence of data points, multiple ABR measurements at different testing frequencies reported within the same study were aggregated into a single effect size. Aggregation was performed using the metafor package’s aggregate.escalc() function, which applies inverse-variance weighting to compute a single combined effect size per study. This method accounts for within-study correlations and ensures that each study contributes only one independent effect size to the meta-analysis, minimizing bias due to data dependency. A random-effects meta-analysis was then performed using R Studio’s “meta” and “metafor” packages, 18,19 with study weighting based on the inverse variance method. The pooled measure was the weighted mean difference (WMD), reported with 95% confidence intervals (95% CI). Heterogeneity was assessed using Higgins and Thompson’s I 2 and τ2 statistics.
Exploratory subgroup analyses were conducted to examine variability in treatment effects, focusing on differences between animal models (mice vs guinea pigs), deafness induction methods (ototoxic vs. noise-induced), and timing of intervention. As no standardized definition for early versus delayed intervention exists in this context, we defined “early” as gene delivery within ≤4 days of deafening and “delayed” as >4 days, based on the observed distribution across included studies. Leave one out sensitivity analyses were performed to evaluate the robustness of the results and to identify potential sources of heterogeneity. Statistical significance was set at p < 0.05.
Bioinformatic analysis
Data source and experimental design
RNA sequencing data were retrieved from the GSE127683 dataset, where mice were divided into two groups: (1) a control group (n = 34), injected with Adenovirus serotype 5 (Ad5) and the red fluorescent protein tdTomato, and (2) an experimental group (n = 22), injected with Ad5 containing the ATOH1 gene flanked by a T2A sequence and regulated by the Ef1a promoter. RNA was then sequenced for both groups, providing the data that served as the basis for our subsequent bioinformatics analysis.
Bioinformatics and functional analysis
A bioinformatics analysis examined gene expression changes associated with ATOH1 gene therapy, corroborating our meta-analysis. Differential expression was performed using the “DESeq2” package (fold change [FC] = 2, false discovery rate [FDR] = 0.05). 20 Gene expression data were normalized with the variance-stabilizing transformation method, with missing values imputed using the gene median method, which replaces absent values with the median expression value of the respective gene across all samples as to preserve the central tendencies and reduce influence of outliers. Genes with count per million lower than 0.5 were excluded.
K-means clustering was applied to the top 1000 genes. Exploratory data analysis aided by visual interpretation of the gene clusters identified six clearly defined clusters with distinct subgroupings, capturing the data’s underlying complexity and displaying biologically interpretable features with optimal within-cluster consistency and between-cluster separation (Supplementary Fig. S1). Both differential gene expression and K-means clustering were performed using the IDEP 2.0 web tool. 21
Gene set enrichment analysis (GSEA) for differentially expressed genes (DEGs) was carried out on Enrichr, 22 with upregulated and downregulated genes analyzed separately, retrieving statistically significant relevant enrichment terms (FDR <0.05). Enriched terms were visualized using Python 3.11’s “matplotlib” package, 23 with metrics like combined score and q-values highlighting key terms.
RESULTS
Study selection
Our search retrieved 960 articles from PubMed, Web of Science and Embase. After removing 205 duplicates, 755 articles were screened by title and abstract. Of these, 742 irrelevant studies were excluded for not meeting the inclusion/exclusion criteria. Thirteen studies underwent full-text screening, of which nine were excluded (three wrong intervention, four lacked the primary outcome of interest, and two had incorrect study design), leaving four studies for inclusion in the meta-analysis. 8,11,24,25 These exclusions included some closely related studies that were nonetheless ineligible due to differences in the experimental model (e.g., genetic rather than acquired hearing loss), use of combinatorial transcription factor strategies instead of ATOH1 monotherapy, or the absence of functional outcome data such as ABR. The study selection process is illustrated in the PRISMA flow chart (Fig. 1).
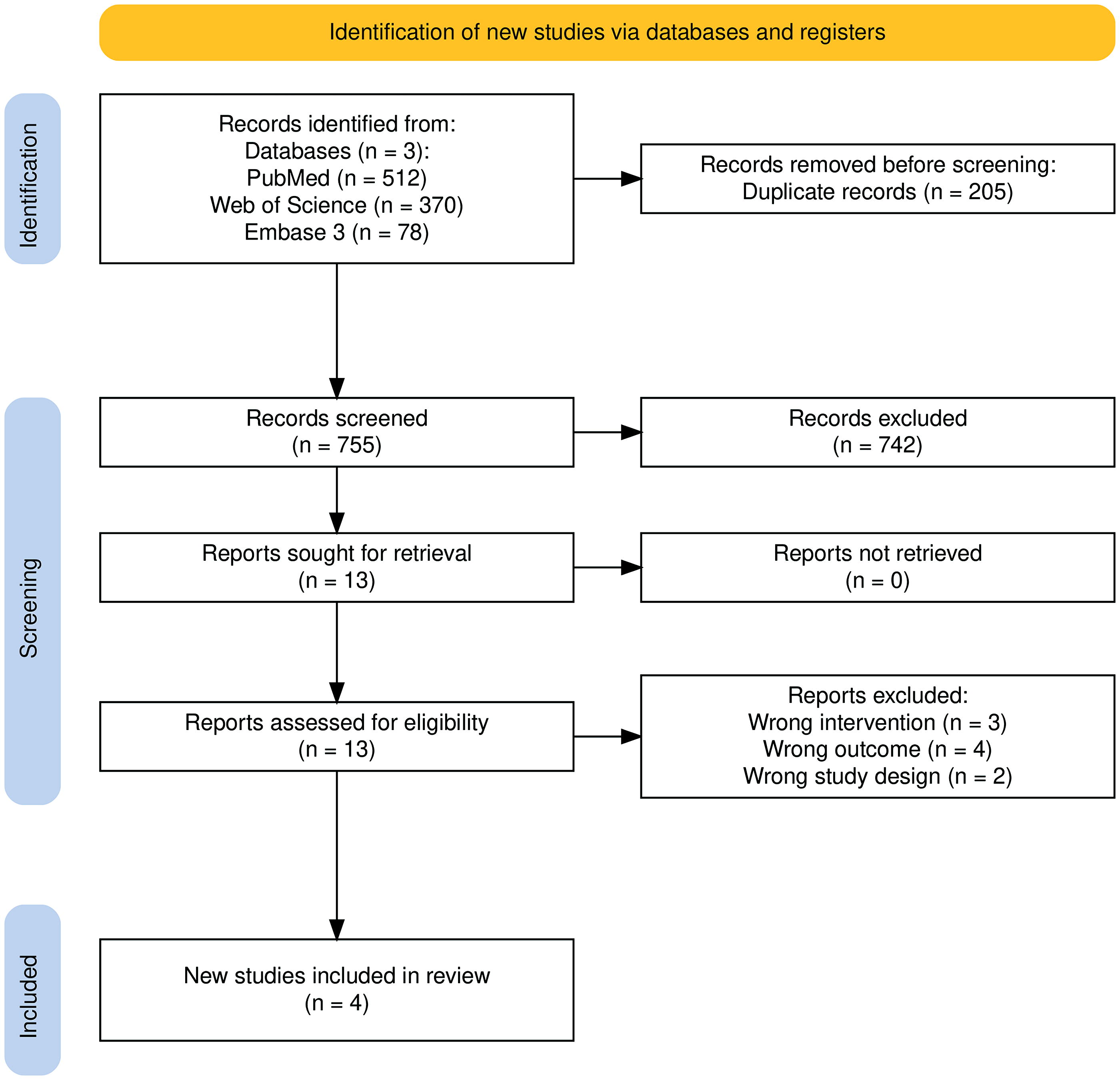
PRISMA Flowchart Diagram.
Characteristics of included studies
Four preclinical trials 8,11,24,25 investigating ATOH1 viral gene therapy were included, all conducted on adult rodent models (n = 52) of acquired SNHL using mice (n = 1) and guinea pigs (n = 3). Adenoviral vectors were used in all studies to deliver ATOH1, aiming to regenerate auditory hair cells and restore hearing function. Vector serotypes included Ad5 or Ad28 where specified. Promoters varied, with some studies employing cytomegalovirus-based constructs or glial fibrillary acidic protein-driven expression. Reported vector doses ranged from 1 to 5 μL, with concentrations between 1 × 1010 and 1 × 1012 PU/mL. The time between deafening and gene delivery ranged from 4 to 10 days, and post-treatment ABR assessment was performed between 3 weeks and 2 months. Three studies 8,11,24 used ototoxic drugs to induce deafness, while one 25 used noise-induced model.
Functional outcomes were assessed by ABR threshold changes, while histological hair cell regeneration was evaluated through immunohistochemical staining in all studies, targeting markers such as Myosin VIIa, Prestin, and stereocilia architecture. Only one study 24 reported separate outcomes for inner vs. outer hair cells, noting that outer hair cell regeneration was not sustained over time. Meanwhile Atkinson et al. (2014) reported no functional improvement in ABR thresholds despite a noted structural regeneration. None of the studies reported transcriptomic data suitable for differential gene expression analysis. Table 1 summarizes the study characteristics.
Summary of Key Methodological Characteristics and Outcomes from Studies Included in the Systematic Review and Meta-Analysis on ATOH1 Gene Therapy for Hearing Restoration
The sexes of the animal models were not reported; it is assumed that there was no sex predilection in these studies and that both sexes were equally represented.
Dose information obtained through direct communication with study authors.
Variables include animal model, deafness induction, vector details, route of administration, timing, outcomes, and histological evidence. None of the studies reported transcriptomic data.
ABR, auditory brainstem response; Ad, Adenoviral vector; CMV, Cytomegalovirus; F, Female; GFAP, glial fibrillary acidic protein; IHC, Immunohistochemistry; IHC, inner hair cells; IP, Intraperitoneal; IV, Intravenous; M, Male; OHC, outer hair cells; OPU, optical particle units; RWM, round window membrane; SC, Subcutaneous; SEM, scanning electron microscopy.
Risk of bias assessment
Risk of bias was assessed using the SYRCLE RoB tool. All studies reported baseline characteristics, while none provided details on random sequence generation. Although random housing information was not reported, the outcomes were minimally affected by housing, so this was judged to be low risk. Selective reporting was unclear, as none of the studies had published protocols. For other biases, 75% of studies showed low risk. Allocation concealment, blinding, and random outcome assessment were mostly unclear. Results are visualized in Figure 2.
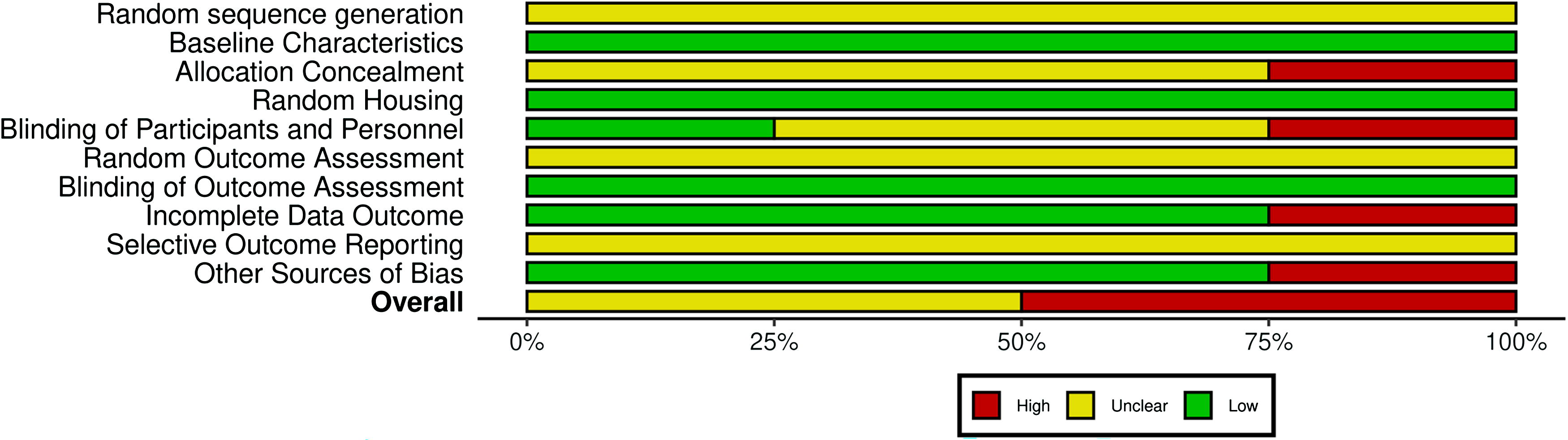
Quality assessment of the studies included in the meta-analysis using the SYRCLE risk of bias tool. The left panel presents the 10 domains that the SYRCLE assesses studies against, each falling under one of the 6 tested biases. Red: high risk of bias, Green: low risk of bias. Yellow: unclear risk of bias. SYRCLE, Systematic Review Center for Laboratory Animal Experimentation.
Although mostly unclear, overall quality is moderate-to-poor, with Kraft et al and Izumikawa et al. showing higher risk of bias, while Atkinson et al. and Yang et al. had lower risk. Detailed SYRCLE domain scores and quality indicators are provided in Supplementary Table S3.
Meta analysis
Meta-analysis of data from four studies showed that ATOH1 gene therapy significantly reduced ABR threshold values in animal models of acquired SNHL, improving hearing. The random-effects model revealed a pooled WMD of −21.37 dB SPL (decibels Sound Pressure Level) (95% CI: [−40.19, −2.54], p = 0.027), indicating the effectiveness of ATOH1 gene therapy in improving hearing thresholds compared to controls.
Although common to meta-analysis of preclinical studies, the between-study heterogeneity was high, with τ2 estimated at 340.47 and I 2 at 98%, indicating considerable variability in the effect of ATOH1 gene therapy across studies, which warrants careful interpretation of the results. Despite the heterogeneity, the pooled results demonstrated a consistent trend favoring the intervention. The combined forest plot illustrating individual study results and the overall effect is presented in Figure 3.
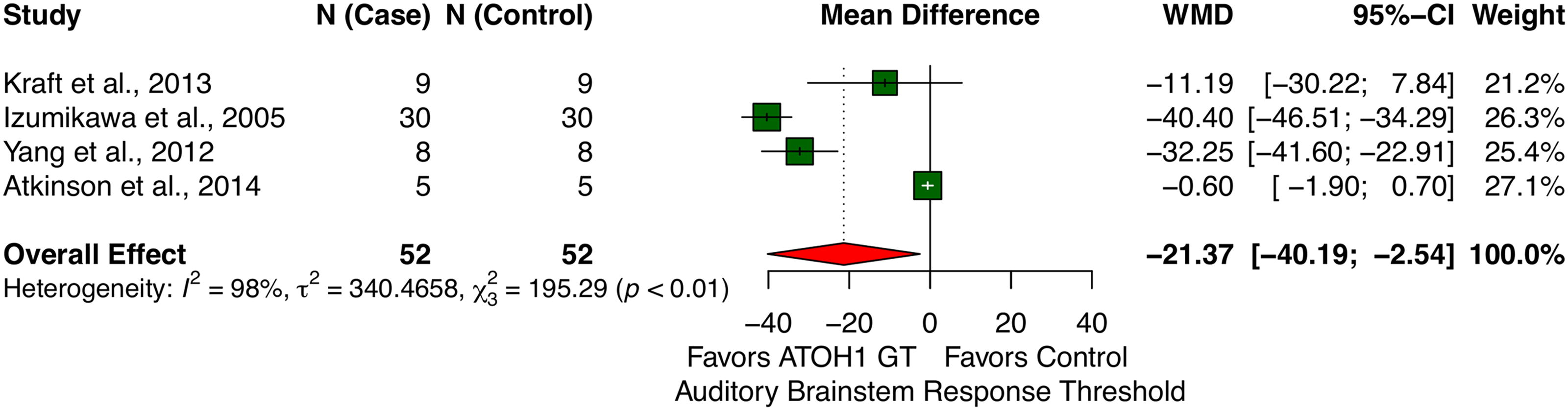
Forest plot summarizing pooled effect sizes (weighted mean differences of ABR values between ATOH1-treated and control groups) from four studies. Negative values indicate reduced ABR thresholds, reflecting improved hearing. ABR, auditory brainstem response.
Subgroup analyses
Subgroup analyses evaluated the efficacy of ATOH1 gene therapy across different animal models (mice vs. guinea pigs) (Fig. 4A). In guinea pigs, ATOH1 gene therapy significantly reduced ABR thresholds, with a pooled WMD of −24.18 dB SPL (95% CI: [−48.29, −0.06]). In mice, the reduction was larger but not statistically significant, with a WMD of −11.19 dB SPL (95% CI: [−30.22, 7.84]). A test for subgroup differences yielded a non-significant result (p = 0.41), indicating no strong evidence that the therapy’s efficacy varies by animal model. These findings suggest that while ATOH1 gene therapy may improve ABR thresholds, the degree of efficacy does not significantly differ between mice and guinea pigs in this analysis. Further research is warranted to explore interspecies variations and their implications for translation to human therapies.
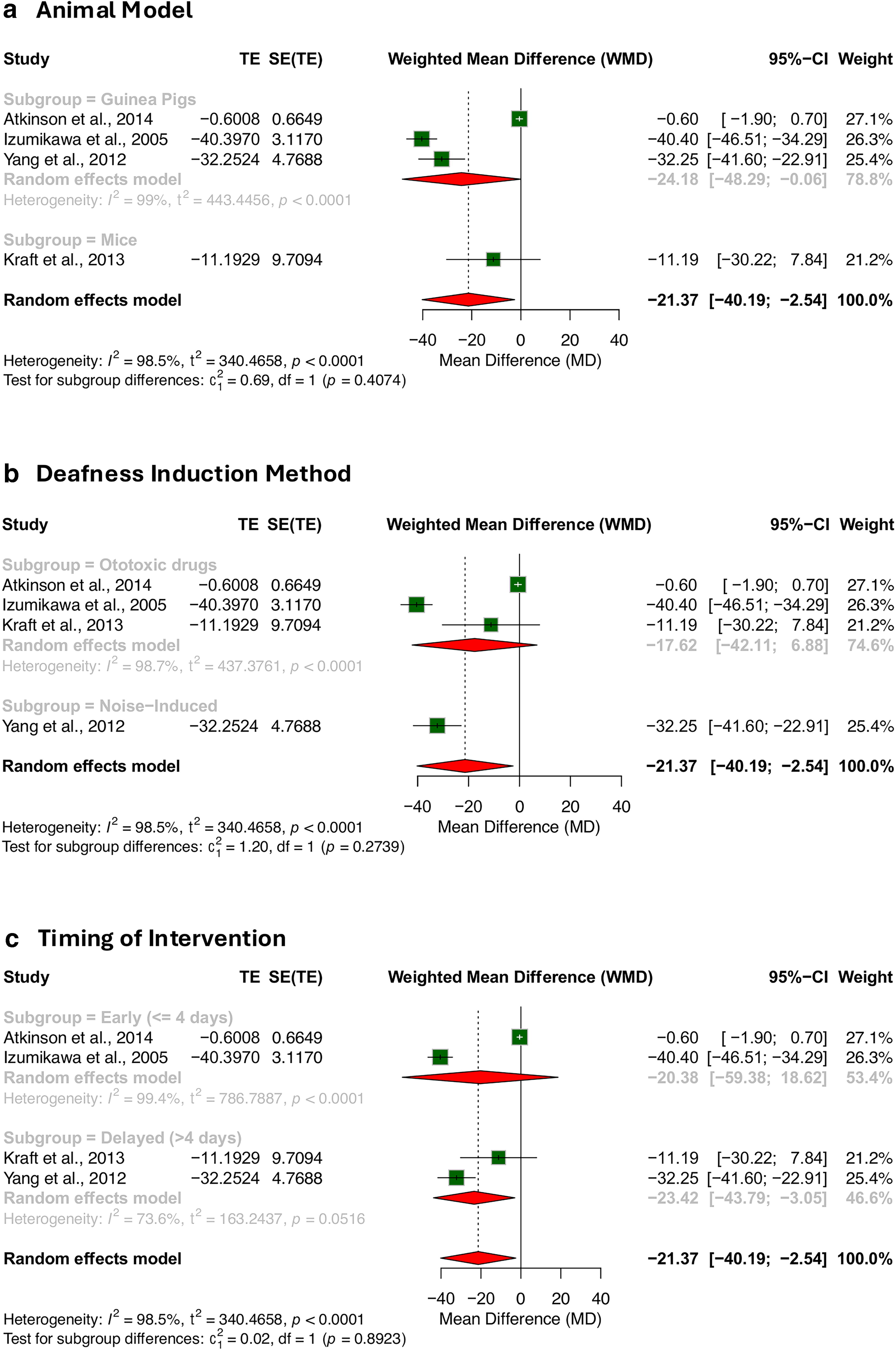
Subgroup analyses of ATOH1 gene therapy efficacy by
Further subgroup analyses explored whether the method of deafness induction influenced treatment outcomes (Fig. 4B). Among studies using ototoxic drugs, the pooled WMD was –17.62 dB SPL (95% CI: [–42.11, 6.88]), a non-significant reduction. By contrast, the noise-induced model (represented by a single study) showed a significant WMD of –32.25 dB SPL (95% CI: [–41.60, –22.91]). Despite the visual difference in effect sizes, the test for subgroup differences did not reach significance (p = 0.27), which suggests that ATOH1 gene therapy may be effective across different models of acquired hearing loss, regardless of the method of deafness induction.
Studies were categorized as “early” (gene delivery within ≤4 days post-deafening) or “delayed” (>4 days), based on the distribution across the included studies (Table 1). In the early group, the pooled WMD was –20.38 dB SPL (95% CI: [–59.38, 18.62]) and did not reach statistical significance. In contrast, the delayed treatment group demonstrated a significant reduction in ABR thresholds, with a pooled WMD of –23.42 dB SPL (95% CI: [–43.79,–3.05]). However, the test for subgroup differences again showed no statistical significance (p = 0.89) (Fig. 4C). These results raise the hypothesis that delayed intervention may permit more effective transgene uptake or regeneration, but definitive conclusions are limited by sample size.
All in all, given the small number of included studies and subgroup imbalance, these findings should be interpreted cautiously and considered exploratory and hypothesis-generating rather than definitive.
Leave-One-out sensitivity analysis
Omitting Kraft et al. (2013) resulted in a WMD of −24.18 (95% CI: [−48.29, −0.06]; p = 0.05), indicating that the overall effect remained borderline significant. Excluding Izumikawa et al. (2005) yielded a WMD of −14.51 (95% CI: [−34.09, 5.06]; p = 0.15), and omitting Yang et al. (2012) produced a WMD of −17.62 (95% CI: [−42.11, 6.88]; p = 0.16), both leading to non-significant results. In contrast, removing Atkinson et al. (2014) resulted in a WMD of −30.14 (95% CI: [−45.29, −14.99]; p < 0.01), maintaining statistical significance.
These results suggest that no single study is solely responsible for driving the observed effect. However, the shift toward non-significance when excluding Izumikawa et al. (2005) or Yang et al. (2012) indicate that their inclusion plays a role in maintaining the statistical significance of the pooled outcome. The relative consistency of effect direction across iterations reinforces the robustness of the findings, despite the high heterogeneity observed in the full model (Tau 2 = 340.47; I 2 = 98%) (Fig. 5).

Leave-one-out sensitivity analysis. Each row shows the effect size recalculated after omitting one study at a time. While the direction of the effect remains consistent across all iterations, statistical significance is lost when either Izumikawa et al. (2005) or Yang et al. (2012) is omitted, suggesting these studies contribute meaningfully to the pooled effect. The robustness of the findings is supported by the stability of effect direction and the persistence of significance in other omissions, despite substantial heterogeneity in the full model.
Bioinformatics analysis
Clustering analysis of gene expression profiles
K-means clustering on the top 1000 genes ranked by standard deviation identified six distinct clusters based on gene expression patterns (Fig. 6). Fifteen genes involved in hearing-related processes were highlighted to determine their respective clusters.
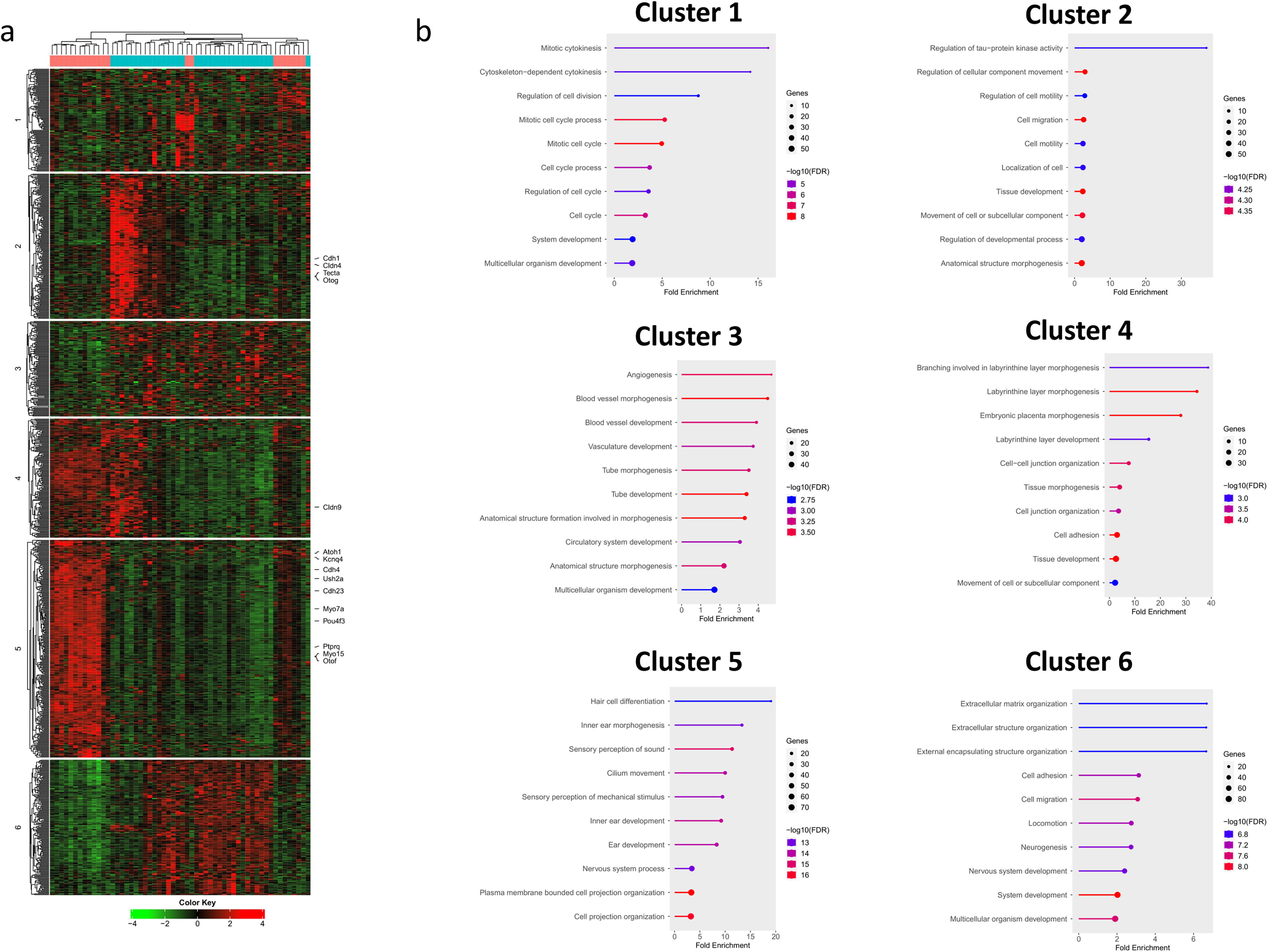
Cluster 1 was associated with cell-cycle regulation, with enriched terms such as mitotic cytokinesis, regulation of cell division, and mitotic cell-cycle, though expression patterns varied widely across control and experimental groups without any consistent trend. Cluster 2, enriched for terms related to cell–cell adhesion, motility, and migration, and regulation of cellular component movement, included four genes involved in hearing loss—Cdh1, Cldn4, Tecta, and Otog— (refer to the heatmap) and showed heavy upregulation in some control samples while being downregulated or unchanged in others.
Cluster 3 was involved in angiogenesis, blood vessel development, and morphogenesis, with more genes upregulated in control samples compared to experimental ones, though no definitive pattern emerged.
Clusters 4 and 5 were the main clusters related to sound perception. Cluster 4 was enriched for labyrinthine layer morphogenesis, tissue morphogenesis, and cell–cell junction organization, which are directly tied to inner ear development, with Cldn9 identified as a key gene related to hearing processes. Cluster 5 was associated with hair cell differentiation, inner ear morphogenesis, sensory perception of sound, inner ear development, and sensory perception of mechanical stimulus, identifying 10 genes related to hearing loss: ATOH1, Otof, Kcnq4, Pou4f3, Ptprq, Myo15, Myo7a, Cdh23, Ush2a, and Cdh4. In these clusters, the experimental ATOH1 group exhibited higher expression of these hearing-related genes compared to the control group.
Finally, cluster 6 was enriched for terms related to extracellular matrix organization, cell adhesion, and migration, with higher expression of genes in control samples compared to the experimental group. This suggests that this cluster is more involved structural support processes rather than directly contributing to hearing.
As a result, the clustering analysis revealed distinct gene expression patterns between the control and ATOH1-treated groups. While clusters 1, 3, and 6 primarily represent general cellular processes like cell cycle regulation, angiogenesis, and extracellular matrix organization, clusters 4 and 5 stand out as key regulators of auditory function. The significant upregulation of genes such as ATOH1, Pou4f3, Myo7a, and Otof in clusters 4 and 5 highlights the biological processes essential for hair cell differentiation, inner ear morphogenesis, and sensory perception of sound. These findings strongly support the hypothesis that ATOH1 gene therapy promotes auditory function restoration through the activation of pathways critical to inner ear and hearing.
Differential gene expression analysis
Differential gene expression analysis using DESeq2 (FC >2, FDR <0.05) identified 1891 upregulated and 1805 downregulated genes following ATOH1 treatment (Fig. 7a, b). The volcano plot (Fig. 7c) reveals a clear separation between upregulated and downregulated genes, with ATOH1, Gfi1, Barhl1, and Pou4f3 showing significant upregulation in the experimental group, supporting their role in hair cell differentiation and sensory perception of sound. Downregulated genes from cluster 2, such as Cdh1, Cldn4, Tecta, and Otog, are primarily involved in cell adhesion and junction organization, and their downregulation may reflect structural changes in response to ATOH1 therapy.

Functional enrichment analysis of upregulated and downregulated genes
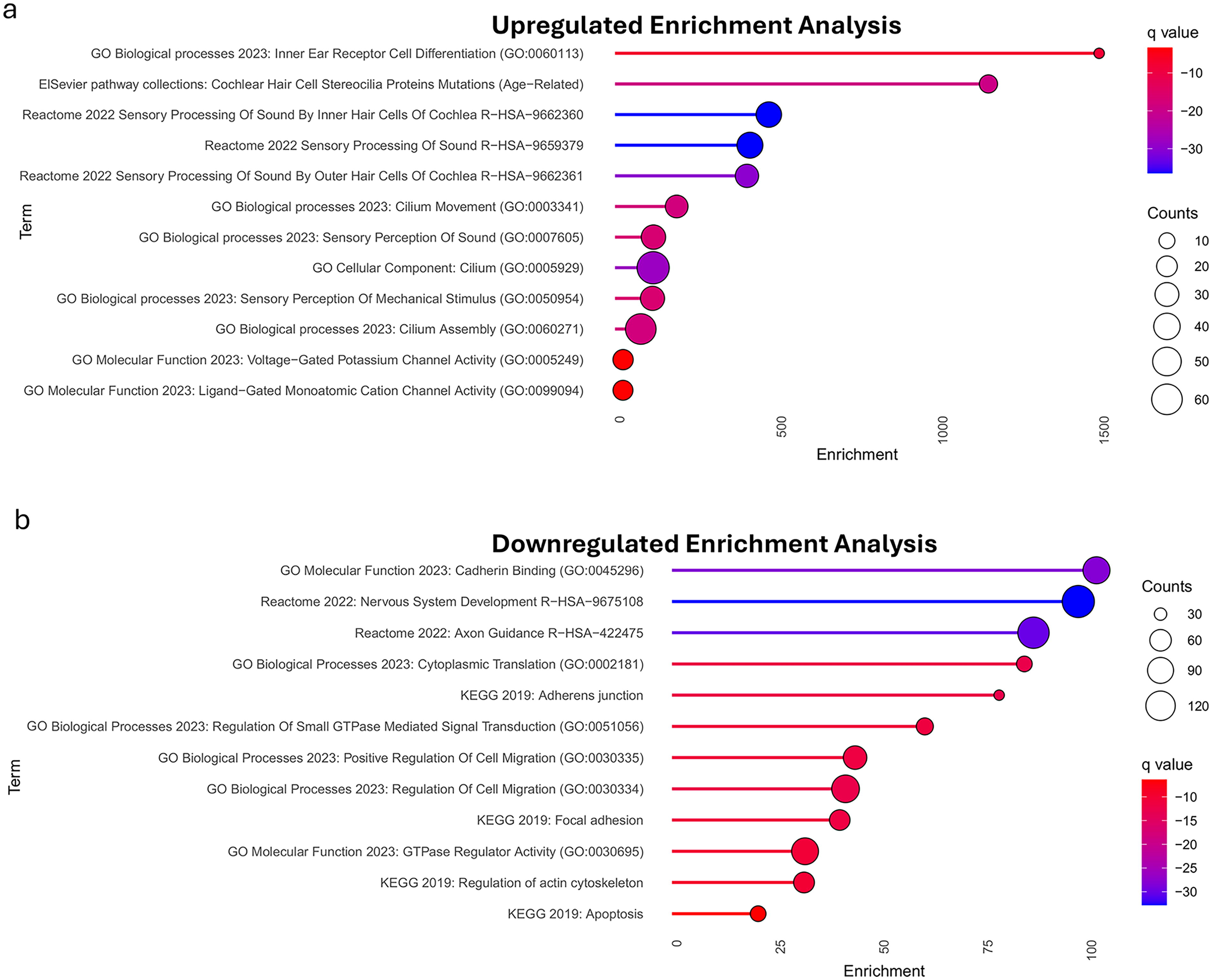
GSEA showed upregulated genes enriched for hearing-related terms, such as inner-ear hair cell differentiation and sensory sound processing (Fig. 8a). Downregulated genes were linked to cadherin binding, adherens junction, regulation of cell migration, nervous system development, axon guidance (Fig. 8b). These results align with the functional clusters of the highlighted genes.
DISCUSSION
Summary of findings
Our meta-analysis findings demonstrate that ATOH1 gene therapy has significantly reduced ABR threshold values in animal models with acquired sensorineural hearing loss, indicating an improvement in hearing. No statistically significant subgroup difference among animal models used reflects the translational potential of ATOH1 gene therapy posing a robust effect irrespective of the model tested. Subgroup analyses further examined whether treatment outcomes varied by deafness induction method or timing of gene delivery. Although only the noise-induced and delayed intervention subgroups showed statistically significant within-group improvements, no significant differences were found between any of the subgroups. This suggests that ATOH1 gene therapy may be broadly effective across various models and delivery timelines, though findings remain exploratory given the limited number and imbalance of included studies. Furthermore, LOOA demonstrated that while some studies had a greater influence on significance than others, no single study disproportionately drove the overall results, supporting the stability of the pooled effect.
To corroborate and validate our meta-analysis findings, bioinformatics analysis was conducted. Cluster analysis for gene expression revealed six clusters, with two key clusters being related auditory functions and hearing restoration following ATOH1 gene therapy administration. Furthermore, differential gene expression analysis revealed that the experimental ATOH1 samples exhibited greater expression of most genes pertinent to the aforementioned clusters in comparison to the control group. For the mice samples given ATOH1 gene therapy, ATOH1 itself along with its target genes Gfi1, Barhl1, and Pou4f3 were all significantly upregulated. Through GSEA analysis, it became evident that the set of upregulated genes were involved in paths related to inner ear receptor differentiation, cochlear hair cell stereocilia mutations, sensory processing of sound by inner hair cells of cochlea, sensory processing of sound, and sensory processing of sound by outer hair cells of cochlea. Therefore, this bioinformatics analysis revealed the intricate nuances of how ATOH1 gene therapy significantly improves hearing threshold, as indicated in our meta-analysis.
Comparison with existing literature
Our findings are consistent with several key studies on ATOH1’s role in hair cell differentiation and regeneration. Bermingham et al. demonstrated that ATOH1 is crucial for hair cell growth and differentiation, affirming the gene’s fundamental role in auditory function. 26 Additionally, a study by Luo et al. investigated unique therapeutic approaches for hair cell regeneration by exploring the roles of enhancer elements in controlling the expression of ATOH1 gene, which promoted hair cell development and subsequently improved hearing. 27 In an in-vitro study utilizing an ATOH1-expressing plasmid, Zheng and Gao demonstrated that transfection led to the generation of additional hair cells, further underscoring the role of ATOH1 in hair cell maturation and differentiation. 28 Iyer and Groves’ study further elucidated ATOH1’s mechanism of action by demonstrating that ATOH1 not only drives hair cell differentiation but also reprograms adjacent non-sensory supporting cells to adopt a sensory cell fate, that is, promoting their transdifferentiation into hair cells. 29
Other studies have confirmed that the canonical roles of Wnt and Notch pathways in cochlear hair cell development and regeneration, with ATOH1 identified as a key downstream target responsible for hair cell differentiation and proliferation. 30 This underscores ATOH1’s regenerative potential. Furthermore, Izumikawa et al. provided immunohistochemical evidence for ATOH1’s role, showing its nuclear localization and upregulated expression in non-sensory supporting cells following Ad.Atoh1 inoculation. 8 This confirmed the efficacy of ATOH1 transduction and the subsequent transdifferentiation process. Additionally, SEM images from the same study demonstrated the regeneration of hair cells post-inoculation. 8 Zhao et al. also observed, through SEM and immunohistochemistry, increased hair cell numbers and intact stereocilia following ATOH1 overexpression, further supporting its role in hair cell regeneration. 31
Our study is centered around a novel approach by using viral gene delivery vectors to upregulate ATOH1. In this context, Tan et al. demonstrated the safety and efficacy of “Adeno-associated virus — inner ear (AAV-ie)” vectors for delivering ATOH1 to the cochlea. Furthermore, SEM in the same study revealed that the gene therapy applied posed no toxic effects on the original functional hair cell population, preserving their number and assembly, which implies that the viral vector does not bring about any negative effects on the auditory function. 10
Recent research also suggests that combining ATOH1 with other genes can enhance its therapeutic effect. A 2020 study found that co-delivery of ATOH1 and GFI1 produced 6.2 times more hair cell-like cells than ATOH1 alone, indicating a synergistic effect. 32 Costa et al. showed that co-transfection of ATOH1, GFI1, and POU4F3 increased the efficiency of hair cell transformation, while ATOH1 alone directed cells toward a neuronal rather than a mechanosensory fate. 33 Likewise, co-expression of ATOH1 and Isl1 significantly boosted hair cell reprogramming compared to ATOH1 alone. 34 Most recently, Zhang et al. (2024) employed an AAV-delivered gene cocktail (GPAS: Gfi1, Pou4f3, Atoh1, and Six1), which led to robust hair cell regeneration based on immunohistochemistry; however, ABR measurements showed no functional hearing restoration. 35 This highlights a recurring challenge in the field, where structural regeneration does not always translate into functional recovery.
While our meta-analysis demonstrated an overall improvement in ABR thresholds following ATOH1 gene therapy, some individual studies reported variable responses across specific frequencies (Table 1). However, no studies to date have systematically examined frequency-specific effects in a controlled or comparative manner. Interestingly, findings by Russel et al. showed that time restricted ATOH1 expression is essential for the survival and differentiation of hair cells (HCs), as its prolonged overexpression impedes, instead of promotes, their maturation. 36 This raises the possibility that variability in expression timing or levels may contribute to inconsistent frequency-specific outcomes observed in some studies.
Limitations
While our findings provide valuable insights into the potential of ATOH1 gene therapy for treating sensorineural hearing loss, several limitations should be acknowledged. First, the high heterogeneity (I 2 = 98%) observed across studies highlights significant variability in study design, animal models (guinea pigs and mice), and intervention protocols, which limits the comparability and generalizability of the results. Although the leave-one-out sensitivity analysis confirmed the robustness of the findings, omitting certain studies led to shifts in significance, indicating their contribution to heterogeneity. Thus, while no single study solely drove the overall effect, the high heterogeneity warrants cautious interpretation. Additionally, despite a comprehensive database search, only four preclinical studies met the inclusion criteria and was included in the meta-analysis. The small sample size limits the generalizability of our results and complicating the assessment of publication bias as recommendations suggest at least 10 studies to be included to warrant assessment of publication bias. 37 Given this limitation, we chose to omit publication bias analysis. Another limitation is the confounding between animal model and viral serotype: The only study using a non-Ad5 vector (Ad28) was also the only study conducted in mice. This overlap precluded meaningful subgroup analysis by serotype, as any observed differences would be indistinguishable from species effects. Future studies should be sufficiently powered and better stratified to disentangle the contribution of viral serotype from other methodological factors influencing treatment outcomes.
Moreover, some studies lacked measures of variability, which we addressed by imputing SD values, though this may still affect the accuracy of our findings. Additionally, the high heterogeneity and risk of bias, common in preclinical studies, complicate the interpretation of pooled results, likely due to variations in intervention protocols, outcome measures, and study designs. Furthermore, since our study focused on guinea pigs and mice as models for analysis, it could not fully represent the effectiveness of ATOH1 in humans. Therefore, human clinical trials are needed to confirm the results and improve generalizability. Additionally, variations in the transduction routes in the included studies is a limitation, as the efficiency and effectiveness of these different routes vary, ultimately affecting the pooled effect. These limitations underscore the need for future trials to thoroughly investigate ATOH1 gene therapy’s effectiveness to assess its potential as a treatment for hearing loss.
A limitation of the transcriptomic analysis is that it is based on a single dataset (GSE127683) derived from a specific experimental model. While the results align with known ATOH1-associated pathways, validation across additional datasets and experimental conditions is needed to strengthen the generalizability of these findings. Finally, the choice of six clusters in our bioinformatics analysis was guided by exploratory data patterns, which, although reasonable, was somewhat intuitive and may benefit from more rigorous validation in future studies.
Future prospects and clinical implications
Given the limited functional studies on auditory outcomes following ATOH1 gene therapy, future research should focus on the mechanosensory transduction ability of newly generated hair cells. Understanding the extent to which these cells can contribute to hearing restoration could inform gene therapy’s therapeutic potential. Furthermore, current reprogramming strategies often involve prolonged transcription factor expression, which deviates from natural development. Research is needed to determine how this prolonged expression impacts the long-term functionality and maturation of reprogrammed cells. Future studies should prioritize enhancing the survival and developmental maturity of reprogrammed hair cells, as they currently only survive for a few weeks. Ensuring the long-term functionality of these cells is crucial for developing effective treatments.
The clinical significance of our study lies in its potential to advance gene therapy as a treatment for sensorineural hearing loss, which currently lacks regenerative options. Future research should involve more trials with diverse animal models and, eventually, human clinical trials to determine the effectiveness of ATOH1 therapy. Additionally, a deeper understanding of the mechanisms underlying ATOH1-mediated HC regeneration could improve gene therapy outcomes, particularly when combined with other therapies. Investigating the mechanisms behind frequency-specific responses in both animal models and humans will guide future research and clinical applications. Moreover, future work should also explore combinatorial gene therapy approaches involving ATOH1 and other synergistic factors (e.g., Gfi1, Barhl1) to enhance both regeneration and functional recovery. Beyond that, future studies should be adequately powered to explore the functional implications of key methodological variables such as the mode of deafness induction (e.g., ototoxic vs. noise-induced), timing of ATOH1 gene delivery, and vector-related parameters including viral serotype, promoter selection, and delivery method. These factors may influence treatment efficacy but remain underexplored due to insufficient reporting and limited comparative data in current preclinical literature.
While studies like Atkinson et al. 11 have demonstrated hair cell regeneration, they often fail to show corresponding improvements in ABR thresholds, highlighting a persistent structural—functional mismatch. To address this, future research should employ transcriptomic tools such as DEG analysis to compare models with and without functional recovery. This would enable identification of key transcriptional signatures, regulatory networks, or missing molecular components that distinguish “structural-only” from “functionally competent” regenerated hair cells. Ultimately, this could help refine gene therapy strategies to achieve not only morphological regeneration but also durable, functional auditory rescue.
Finally, clinical trials must rigorously assess the safety of ATOH1 gene therapy in humans to ensure there are no short- or long-term risks. Identifying the most efficient and safe in vivo transduction routes is also essential for optimizing reprogramming therapies. Such advances will be crucial for the successful implementation of ATOH1 gene therapy in treating sensorineural hearing loss.
CONCLUSION
The findings of this systematic review and meta-analysis provide compelling evidence supporting the potential of ATOH1 gene therapy in restoring auditory function in preclinical models of acquired SNHL. Our meta-analysis revealed a significant reduction in ABR thresholds following ATOH1 gene therapy, suggesting improved hearing recovery. Additionally, our bioinformatics analysis demonstrated robust upregulation of key genes involved in hair cell differentiation and inner ear morphogenesis, further reinforcing the therapeutic potential of ATOH1. However, the high heterogeneity across studies, limited sample size, and risk of bias highlight the need for cautious interpretation of these findings. The absence of human clinical trials remains a critical gap, underscoring the necessity for well-designed studies to assess safety, efficacy, and long-term functional outcomes. Future research should focus on optimizing gene delivery techniques, evaluating the longevity of newly generated hair cells, and exploring combinatorial approaches that enhance ATOH1-mediated regeneration. While ATOH1 gene therapy presents a promising avenue for SNHL treatment, further investigations are required to bridge the gap between preclinical success and clinical applicability. Ultimately, the translation of these findings into human trials could revolutionize hearing restoration strategies and significantly improve outcomes for individuals with acquired SNHL.
Footnotes
DATA AVAILABILITY STATEMENT
The datasets generated and analyzed during the current study are available in publicly accessible repositories. Specifically, the differential gene expression data used for bioinformatic analysis were retrieved from the Gene Expression Omnibus (GEO) database under the accession number GSE127683. Additional data generated during the meta-analysis are available from the corresponding author upon request.
ETHICS STATEMENT
This study is a systematic review and meta-analysis using publicly available data from previously published studies. No new experiments involving human participants or animals were conducted. Ethical approval was not required for this secondary analysis.
AUTHORS’ CONTRIBUTIONS
A.S.: Conceptualization, formal analysis, investigation, project administration, writing—original draft preparation, writing—review and editing. O.Y.: Data curation, formal analysis, investigation, visualization, writing—original draft preparation. N.A.-A.: Data curation, formal analysis, investigation, writing—original draft preparation. F.Q.: Investigation, methodology, writing—original draft preparation. Z.A-S.: Investigation, methodology, writing—original draft preparation. S.S.: Investigation, methodology, writing—original draft preparation. M.A.-T.: Investigation, methodology, writing—original draft preparation. L.K.: Supervision, conceptualization, methodology, project administration, writing—review and editing.
AUTHOR DISCLOSURE STATEMENT
The authors declared no competing interests.
FUNDING INFORMATION
No funds, grants, or other support was received.
SUPPLEMENTARY MATERIAL
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.