
Abstract
We recently isolated and identified Zizimin2 as a functional factor that is highly expressed in murine splenic germinal center B cell after immunization with T-cell-dependent antigen. Zizimin2 was revealed that it is a new family member of Dock (dedicator of cytokinesis), Dock11, which was the guanine nucleotide exchange factor for Cdc42, the Rho family of low molecular weight GTPases. However, neither the molecular function of Zizimin2 in the acquired immunity nor the physiological role in decrease of immunological activity along with aging has been known. To facilitate the further analysis of physiological function of this molecule, we have established rat monoclonal antibodies (MAbs) against mouse Zizimin2 protein. Recombinant His-tagged partial mouse Zizimin2 protein was expressed in Escherichia coli, purified, and applied as an antigen for immunization. The desired hybridomas were selected by the recombinant Zizimin2 protein coated enzyme-linked immunosorbent assay screening, and we generated stable hybridoma cell lines that produced the antibody against murine Zizimin2 protein. We determined their isotypes and further examined capability or availability in immunoblotting, immunoprecipitation, or immunofluorescence microscopy. Here we demonstrated the several appropriate antibodies for immunoblotting and immunofluorescence microscopy. These MAbs should therefore be very useful tools for study of the characterization of the Zizimin2 protein and the understanding of the biological function of Zizimin2-mediated immunosenescence.
Introduction
Materials and Methods
Cells and culture conditions
If not otherwise specified in this study, human and mouse cell lines 293T, mouse embryonic fibroblasts (MEFs), NIH-3T3, and A20 cells were maintained in Dulbecco's modified Eagle's medium (DMEM, Wako Pure Chemical Industries, Osaka, Japan) supplemented with 10% fetal bovine serum (FBS, EQUITECH-BIO, Ingram, TX) and penicillin/streptomycin. WEHI231 cells were maintained in RPMI1640 (Sigma, St. Louis, MO) supplemented with 10% FBS, penicillin/streptomycin, 0.05 mM 2-mercaptoethanol, and 1% sodium pyruvate. The obtained hybridoma cells were maintained in GIT medium (Nihon Pharmaceutical, Osaka, Japan) without antibiotics.
Plasmids and expression
For immunizing antigen, C-terminal-half fragment (Zizimin2 CZH2, 1653–2073aa) was retrieved and inserted into pET15b vector. The following primers were used to amplify the C-terminal half fragment: 5′-CGG GAT CCG CAC CGA AAG AAA TTA TTC CC-3′ (sense) and 5′-CGG GAT CCT CAT ACT TCC GCG TAT CTT G-3′ (anti-sense). For CZH2 domain expression vector, C-terminal region (1569–2073aa) of murine Zizimin2 was amplified and inserted into the retrovirus expression vector. The following primers were used for the amplification: 5′-CTA GGC GCC GGA ATT GAC CAC CAT GAA GAG GAA AAC CTT TCT GAG-3′ (sense) and 5′-ATT GAT CCC GCT CGA GTC ATA CTT CCG CGT ATC TTG-3′ (anti-sense). Expression and purification of recombinant Zizimin2 protein were essentially performed as described previously.(10)
Immunization, cell fusion, and MAb purification
Immunization, cell fusion, and MAb purification were essentially performed as described previously.(10) Briefly, 6-week-old female Wister rats were immunized by footpad inoculation with 50 μg/body of Escherichia coli derived recombinant Zizimin2-CZH2 protein four times every 10 days. Hypoxantine-aminopterin-thymidine selected hybridoma clone candidates were analyzed (the antigen specificity) by enzyme-linked immunosorbent assay (ELISA) screening and then the positive clones were prepared using limiting dilution to obtain the stable clones, which produces anti-mouse Zizimin2 MAbs. As described above, the C-terminal half region of Zizimin2 was also used as a coated antigen (5 μg/mL) in this ELISA screening. The ELISA plates were also washed as described previously(10) and incubated with alkaline phosphatase conjugated goat anti-rat IgG antibody (1:5000, Jackson ImmunoResearch, West Grove, PA) for 1 h at 37°C. The plates were washed as before and incubated with the substrate solution (Bio-Rad, Hercules, CA); then the absorbance was measured at 405 nm by microplate reader to obtain positive clones.
Isotype determination
Both class and subclass of isotypes of established anti-Zizimin2 MAbs were investigated from the corresponding supernatants by a rat monoclonal antibody isotyping test kit (Serotec, Oxford, United Kingdom) following the manufacturer's instructions.
DNA transfection and virus infection
293T cells were transfected with retroviral expression plasmids together with helper retrovirus plasmids, as described previously.(11) Culture supernatants were harvested 24 h after transfection, pooled, and stored at −80°C. Exponentially growing cells (5 × 106 cells/mL) in 100-mm-diameter culture dishes were infected with 5 mL of fresh virus-containing supernatant in complete medium with 8 μg/mL polybrene (Sigma). Infection efficiency was confirmed either by flow cytometric assay for green fluorescent protein (GFP) expression or by selection for drug resistance. The cells were collected 72 h after infection, lyzed, and subjected to immunoblotting analysis.
Immunoblotting analysis
Cells were lyzed by the addition of RIPA buffer (50 mM Tris [pH 7.5], 1% NP-40, 0.5% Na-deoxycholate, 0.05% sodium dodecyl sulfate [SDS], 1 mM ethylenediaminetetraacetic acid, 150 mM NaCl) and protease inhibitors (Roche, Mannheim, Germany) and sonicated on ice for 15 s. Insoluble materials were pelleted by centrifugation at 15,000 g for 20 min at 4°C, and protein in the supernatant was quantified by BCA assay (Pierce, Rockford, IL). Thirty μg of protein dissolved in SDS sample loading buffer were boiled for 5 min, subjected to 6% SDS-PAGE, and transferred to polyvinylidene fluoride membrane. Membrane was blocked in Tris-buffered saline Tween-20 (TBST; 25 mM Tris [pH 7.4], 0.15 M NaCl, 0.1% Tween-20) containing 5% non-fat milk for 24 h at 4°C. Then the membrane was probed for 2 h with hybridoma culture supernatants (1:2), anti-α-tubulin antibody (1:2000) (Sigma), or anti-actin antibody (1:2000) (Sigma) in TBST containing 5% non-fat milk powder, washed three times with TBST, incubated with peroxidase-conjugated goat anti-rat IgG (1:2000) (Jackson ImmunoResearch) or peroxidase-conjugated goat anti-mouse IgG (1:2000) (Jackson ImmunoResearch) for 2 h at room temperature (RT). After several washes with TBST, reaction was visualized with ECL (GE Healthcare ECL plus Western blotting Detection System).
Immunofluorescence microscopy
NIH-3T3 cells cultured on coverslips were washed three times in phosphate-buffered saline (PBS), fixed for 15 min in 4% paraformaldehyde at 37°C, washed in PBS, and permealized with 0.1% TritonX-100 in PBS for 5 min at RT. All subsequent steps were performed at RT. Fixed coverslips were washed three times in PBS, blocked for 1 h with 0.2% bovine serum albumin in PBS, washed three times in PBS, and incubated for 1 h with hybridoma culture supernatant (1:1). Following three washes in PBS, coverslips were incubated for 1 h with Cy3-labeled goat anti-rat IgG (1:500) (Jackson ImmunoResearch) to visualize primary antibodies. Coverslips were washed three times in PBS and mounted onto microscope slides using Vectashield containing 4′,6-diamino-2-phenylindole (DAPI, Vector Labs, Burlingame, CA) for nuclei detection. Images were acquired using a fluorescence microscope.
Results
Establishment of rat anti-Zizimin2 MAbs
We determined the C-terminal region (1654–2073) of mouse Zizimin2 as an immunizing antigen. We screened hybridomas by ELISA with E. coli derived recombinant Zizimin2–CZH2 domain as described above. In the rounds of hybridoma screening, we obtained four clones that possessed specific reactivity with murine Zizimin2-CZH2 region (214I9, 121E167, 126F233, and 5A10) out of 299 clones. As listed in Table 1, we investigated isotypes of each established MAbs. All four clones produced class-switched antibodies; therefore they could potentially react against their epitopes with high affinity. For further analysis, to characterize these MAbs, we quantitatively purified 214I9, 121E167, and 5A10 clones from hybridoma culture supernatants; however only 126F233 was purified from ascites from culture supernatant.
Isotype of each MAb was determined with corresponding hybridoma supernatant. IB, immunoblot analysis; IP, immunoprecipitation; IF, immunofluorescence microscopy.
Characterization of anti-Zizimin2 MAbs
In order to investigate the availability of these MAbs, we first examined the cognate antigen reactivity of the MAb against the recombinant mouse Zizimin2-CZH2 domain by immunoblot analysis. Virus containing the CZH2 domain expression vector was prepared and infected to NIH-3T3 cells in which we could monitor the independent GFP expression as an infection efficiency (Fig. 1B). Then we examined the reactivity of a representative MAb, 214I9, against the recombinant CZH2 domain by immunoblot analysis (Fig. 1). We could demonstrate that 214I9 detected the recombinant CZH2 domain (70 kDa), which was consistent with the result of the ELISA screening. In addition to the cognate antigen, 214I9 also detected the endogenous Zizimin2 protein but not the GFP protein in NIH-3T3 cells (Fig. 1A). Since 214I9 MAb could detect the physiological full-length Zizimin2, we investigated the antigen specificity of all remaining established MAbs in both immune or non-immune cell lines and murine normal splenic cells as a positive control. As we expected, all of the MAbs were able to recognize intact Zizimin2 in mouse B cell lymphoma lines WEHI231 and A20 cells as the same as positive control splenic cells. However, Zizimin2 in mouse non-immune cell lines, MEFs, and NIH-3T3 cells was detected rather weaker than in the immune cells we tested here (Fig. 2). It has been reported that the expression of Zizimin2 gene is limited mainly to lymphoid organs or immune cells; therefore, we next investigated the physiological Zizimin2 protein expression specificity in C57BL/6 mice by immunoblot analysis. The result clarified that Zizimin2 protein is expressed in the lymphoid organs such as thymus, spleen, and lymph node (Fig. 3). In addition, we found a weak expression in intestine; it is known that intestinal epithelial cells induce the immune response against the invading pathogens from food-derived substances and indigenous intestinal bacteria (see discussion). Next, the availability of the MAbs for immunoprecipitation was examined to enrich Zizimin2 protein itself or to co-precipitate with associated molecules as Cdc42. Consequently, we unfortunately could not confirm any immunoprecipitation complex because we used all of the MAbs, including 214I9 (data not shown). To confirm and extend the availability of these antibodies, we finally characterized whether they were useful for immunofluorescence staining. The two MAbs, 126F233 and 214I9, were able to recognize the cognate Zizimin2 protein in NIH-3T3 cells through immunofluorescence microscopy (Fig. 4 and Table 1). In Figure 4, the localization of the Zizimin2 protein seemed to be borderless; in any case, one of the MAb (214I9) detected the signal of endogenous Zizimin2 in quiescent NIH-3T3 cells. These profiles of the MAbs against murine Zizimin2 protein are summarized in Table 1.

Antigen specificity of anti-Zizimin2 214I9 MAb against recombinant Zizimin2-CZH2 proteins by immunoblot analysis. Established anti-mouse Zizimin2 MAb 214I9 recognized endogenous Zizimin2 protein and truncated recombinant Zizimin2-CZH2 protein in the virus containing both GFP and Zizimin2-CZH2 domain expression vector infected into NIH-3T3 cells (

Antigen reactivity of anti-Zizimin2 MAbs against endogenous murine Zizimin2 protein by immunoblot analysis. All four MAbs are described to the left of each panel. The arrowheads represent endogenous Zizimin2 protein. Whole cell lysates of splenic cells and MEFs from C57BL/6 mice, a couple of mouse B lymphocyte cell lines A20, and WEHI231 and NIH-3T3 cells described at the top of the panels were subjected to immunoblotting.

Tissue distribution of Zizimin2 protein expression in C57BL/6 mice. The lysates of the various tissues described at the top of the panel were prepared from 6-week-old C57BL/6 mice. Each protein concentration was quantified and the equal amount of protein (30 μg) was subjected to immunoblot analysis. Molecular weight standards are shown on the left, and the arrowheads indicate the predicted endogenous Zizimin2 proteins and α-tubulin as a loading control on the right.
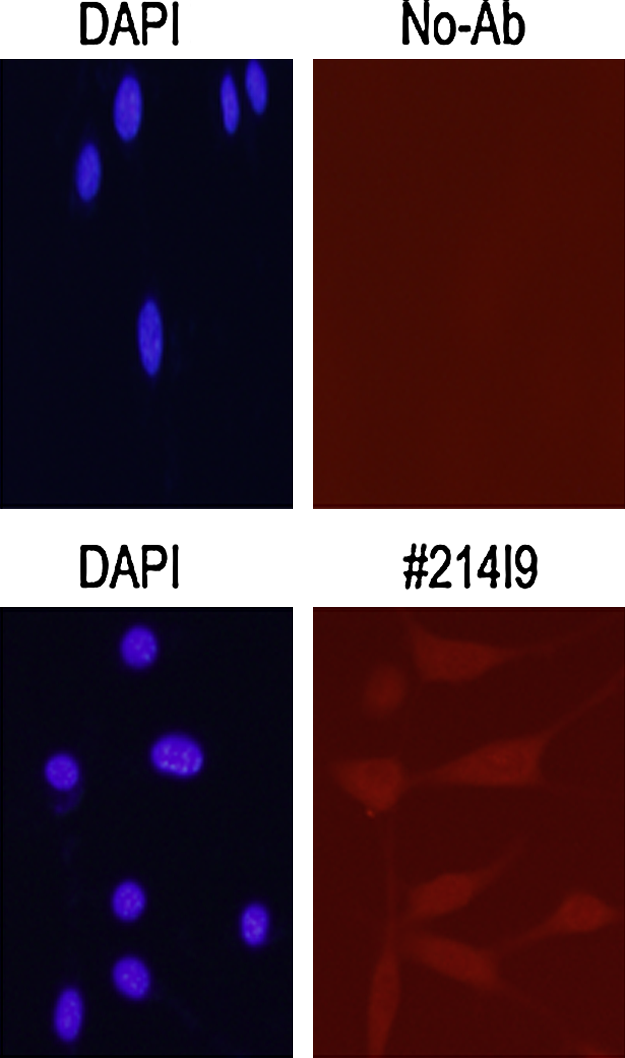
Immunofluorescence microscopy with anti-Zizimin2 214I9 MAb. The anti-mouse Zizimin2 214I9 MAb was subjected to immunofluorescence microscopy to stain the endogenous Zizimin2 protein (magenta) in NIH-3T3 cells. DAPI staining (blue) represents the nuclei of the cells.
Discussion
In this study, rat anti-mouse Zizimin2 MAbs were established against bacterially expressed murine Zizimin2-CZH2 to study the still unknown physiological function of Zizimin2 protein. On the basis of sequence similarity in the CZH2 domain among Zizimin family proteins, we chose to obtain hybridoma clones that possessed the cross-reactivity with human Zizimin2 or even the other Zizimin family proteins, such as Zizimin1. Consequently, as summarized in Table 1, all four MAbs identified here can specifically react with mouse Zizimin2 protein in ELISA and immunoblot analysis, and both 214I9 and 121E167 were available in immunofluorescence microscopy (Fig. 4 and data not shown). Alternatively, we have recently obtained murine cells whose Zizimin2 was completely deleted, and in which any appropriate signal could not be detected by immunoblot analysis (IS and MM, unpublished data); therefore, we consider all antibodies carrying the antigen specificity against Zizimin2 collaterally. On the other hand, these MAbs potentially counteract the binding of Cdc42 or other molecules to functional CZH2 domain. Therefore, we should expect those MAbs to be very useful tools, not only to study Zizimin2 expression, but also to reveal further biological function in immunosenescence.
We demonstrated with the application of the Zizimin2 antibodies for immunoblot analysis that endogenous Zizimin2 protein expression is also limited in lymphoma cell lines as expected. On the other hand, we could detect trace amounts of signals in MEF cells or even in NIH-3T3 cells. These results led us to conclude that Zizimin2 expression is not so strictly regulated in multi-potentiated undifferentiated cells or a kind of immortalized cell line, respectively. Furthermore, to examine the physiological expression of Zizimin2 protein, we carried out another immunoblot with various tissues from 6-week-old mice using 214I9 MAb. In slight contrast to cell lines, a robust expression of Zizimin2 was confirmed in spleen, thymus, or lymph node and faint expression was also detected in intestine (Fig. 3). Taken together with latent intestinal immunity, the tissue distribution of Zizimin2 expression is reminiscent that Zizimin2 is fairly involved in the immunological system. The 214I9 MAb, which is available to detect endogenous Zizimin2 in NIH-3T3 cells morphologically, will give us the capability to pay more attention to reveal molecular and cellular interaction or functional pathway in which Zizimin2 is involved.
It has been reported that the activity of immunological response or function decreases with advancing age.(11–13) Information is limited about immunosenescence; therefore the above reports prompted us to examine any correlation between Zizimin2 expression and aging by using various tissues from both 6-week-old and 2-year-old mice. Consistently, expressions of both mRNA and Zizimin2 protein are similarly regulated in lymphoid organs between young and old mice; however, the relative expression values in thymus, spleen, and lymph nodes from young mice was reasonably higher than in old mice (YJ and MM, unpublished data).
Of note, one of Zizimin family, Zizimin1, is able to interact with Cdc42 through CZH2 domain, and this interaction triggers the induction of the formation of filopodia in NIH-3T3 cells expressed exogenous murine Zizimin1.(2) It is also known that Zizimin1 dimerizes via its CZH2 domain, and kinetic measurements demonstrated increased binding affinity of Cdc42 to Zizimin1 in a Cdc42-dependent manner.(14) In addition, we should note that overexpression of Zizimin1 promoted dendrite growth, whereas knockdown or expression of a mutant Zizimin1 lacking Cdc42 GEF activity suppressed dendrite growth in primary cultured rat hippocampal neurons.(15) Taken together, with the above information and the conserved CZH2 domains between Zizimin1 and Zizimin2 (∼71%), the pressing issue to address is whether Zizimin2 also can promote filopodia formation or dendrite growth by activation of Cdc42. Furthermore, these represented MAbs against murine Zizimin2 lend support to the profound roles of Zizimin2 and reveal a hitherto unrecognized physiological relationship between Zizimin2 and cellular functions, especially in immunosenescence.
Footnotes
Acknowledgments
We thank Drs. M. Sugimoto and H. Nakamura for their skillful technical suggestions and advice on hybridoma cloning. We also appreciate Drs. Y. Naoe and L. Tsuda's helpful comments and discussion. This research was supported in part by a research grant for Longevity Sciences (19C-3) from the Ministry of Health, Labor and Welfare of Japan.