
Abstract
Tudor-SN is a multifunctional regulator of gene expression that has been shown to function as a transcriptional co-activator, regulator of miRNA processing, mRNA splicing, and stability. Tudor-SN has also been identified as a component in RNA-induced silencing complex. Here we have produced and characterized seven monoclonal antibody (MAb) clones against human Tudor-SN. Antibodies were generated against the fourth staphylococcal nuclease-like domain (SN4) and the Tudor domain of human Tudor-SN. The MAbs recognize the Tudor-SN protein in Western blot analysis and immunoprecipitation, and detect the specific antigen in immunohistochemistry assays. One of the antibody clones also recognizes the Drosophila melanogaster and Danio rerio Tudor-SN. Immunocytochemistry of HeLa cells revealed Tudor-SN localization in nucleolus, suggesting a possible new function for the protein in the compartment. An extensive expression analysis in human tissue arrays shows moderate to high expression of Tudor-SN in a wide range of organs and tissues, especially in epithelial cell types.
Introduction
Tudor-SN is expressed in several tissues and organs,(1) and especially highly in a range of exocrine and endocrine cells.(8) Tudor-SN protein has been detected both in cytoplasm and nucleus.(1) Recently, Tudor-SN has been shown to be overexpressed in a range of cancers(9–11) and to have a role in inheritary autosomal-dominant polycystic kidney disease (ADPKD).(12) Investigation of the Tudor-SN protein has been hampered by the lack of good antibodies against the protein and only polyclonal antibodies have been available. Here we have generated mouse MAbs against human Tudor-SN protein that can be used in Western blot, immunoprecipitation, and the detection of the protein in cells and tissues. Interestingly, in immunocytochemistry, one of the generated antibodies gave a signal in nucleolus, suggesting that Tudor-SN could have a function in ribosomal RNA expression, processing, or assembly.
Materials and Methods
Generation of the antibodies
Antibodies were generated as previously described.(13) Immunization of mice was carried out with purified GST-fusion proteins. GST-SN1-4(3) (corresponding to aa 26–664 of the human Tudor-SN) and GST-TSN (corresponding to aa 664–910; see Tudor-SN domain organization, Fig. 1) were used for initial immunizations. In order to create SN4 specific antibodies, the SN4 domain (aa 507–674) was PCR cloned and inserted into pGEX4T-1 vector, expressed in bacteria, and purified using Glutathione Sepharose 4B (GE Healthcare, Uppsala, Sweden). The SN4 protein was purified from the GST by digestion with thrombin and dialyzed against phosphate-buffered saline (PBS) before it was used to boost the production of SN4 domain specific antibodies. Similarly, the TSN domain was purified from its GST fusion partner and used to boost the GST-TSN immunized mice. Splenocytes from the immunized/boosted mice were fused with the myeloma cell line Sp2. The generated hybridomas were cultured in HAT selection medium, subsequently in HT medium, and finally in RPMI supplemented with 10% fetal bovine serum (FBS), glutamine, and antibiotics. Specific hybridomas were screened by using homologous antigens in indirect ELISA and cells from positive wells were cloned and recloned by the method of limiting dilution. The functionality of the produced MAbs was further evaluated in different immunoassays. For the evaluation procedures, the concentrations of the different MAbs in hybridoma cultures were quantified by immunoblotting with polyclonal anti-mouse IgG and the concentrations were adjusted to equal by addition of medium.

Domain organization of Tudor-SN and the functionality of generated MAbs in Western blot and immunoprecipitation. (
Cell lysates, Western blot analysis, and immunoprecipitation
HeLa cells were grown in DMEM supplemented with 10% FBS, penicillin-streptomycin, and non-essential amino acids. Confluent cells were washed with PBS, scraped and pelleted by centrifugation, and lysed on ice using 10–20 volumes of 1% Triton X-100 lysis buffer (50 mM Tris [pH 7.5], 150 mM NaCl, 1% Triton X-100, 10% Glycerol, 1 mM EDTA) supplemented with protease inhibitor cocktail set III (Calbiochem/EMD Biosciences, San Diego, CA). S2 cells were cultured in Schneider's medium supplemented with 10% FBS and penicillin-streptomycin. Zebrafish (Danio rerio) extracts were prepared from 3-day-old embryos using RIPA lysis buffer (1% Triton X-100 lysis buffer with additional 0.1% SDS and 0.5% sodium deoxycholate) with sonication.
The lysates were cleared by centrifugation and ∼30 μg protein/lane of the whole cell lysates were resolved on an 8% SDS-PAGE and blotted. Antibodies from hybridoma cell culture supernatants were diluted by one-fourth in TBS-T (Tris-buffered saline supplemented with 0.1% Tween), supplemented with 2 mg/mL bovine serum albumin (BSA), and used in immunodetection. Biotin-conjugated anti-mouse secondary antibody (Dako, Glostrup, Denmark) and streptavidin-conjugated horseradish peroxidase (GE Healthcare, Buckinghamshire, United Kingdom) were used together with ECL Western blotting detection reagents (GE Healthcare) to detect the signal.
Immunoprecipitation was performed with antibodies from 0.5 mL of hybridoma cell culture supernatants that were prebound to Protein-G Sepharose (GE Healthcare, Uppsala, Sweden). Whole cell extract from HeLa cells was incubated with the Protein-G Sepharose bound antibodies for 2 h on rotation at +4°C. The immune complexes were washed four times with the 1% Triton X-100 lysis buffer and boiled in SDS-PAGE loading buffer before SDS-PAGE and immunoblotting.
Immunocytochemistry and immunohistochemistry
HeLa and S2 cells grown on glass coverslips were fixed in 4% paraformaldehyde in PBS for 10 min at room temperature. After PBS washes, the cells were permeabilized with 0.2% Triton X-100 in PBS for 10 min. Coverslips were incubated for 1 h at +4°C with hybridoma cell culture medium from the individual clones that had been diluted 1:1 in PBS with 2 mg/mL BSA. After PBS washes, the coverslips were incubated for 1 h with the secondary antibody Alexa 488-conjugated goat anti-mouse IgG (Molecular Probes, Eugene, OR), and after additional PBS washes mounted using Immunomount (Dako, Glostrup, Denmark). Immunohistochemistry of human tissue arrays using the anti-SN4 clone α-SN4-1 was performed by the Swedish Human Proteome Resource (HPR) program(14,15) (
Results and Discussion
Tudor-SN is an interesting protein that has been assigned several functions in regulation of transcription and RNA biogenesis, as well as in certain diseases. Characterization of the Tudor-SN protein has been hampered by the lack of good antibodies. We wanted to create mouse MAbs against Tudor-SN that could be used as tools for studying the functional and physiological roles of this multifunctional regulator of gene expression. In order to create these antibodies, mice were first immunized with GST-SN1-4 and GST-TSN fusion proteins. Individual GST-SN1-4 immunized mice were boosted with purified SN3 and SN4 domains. The SN3 domain immunization did not produce any antibodies that recognize full-length Tudor-SN, whereas SN4 domain turned out to be highly immunogenic, resulting in dozens of hybridoma clones expressing antibodies against Tudor-SN. Five α-SN4 clones were chosen for further characterization. Similarly, GST-TSN immunized mice were boosted with pure TSN protein and two clones produced Tudor-SN recognizing MAbs. The generated MAbs are summarized in Table 1.
Data not shown. Immunocytochemistry with the α-TSN-1 clone resulted in a weaker signal compared to that of other antibodies.
The estimated suitability is indicated for each clone in Western blot for detection of the human Tudor-SN (Fig. 1B) and in immunoprecipitation (IP) (Fig. 1C). Signal in immunocytochemistry (ICC) indicates the detected localization of the endogenous Tudor-SN in HeLa cells in either cytoplasm or nucleus.
The SN domains 1, 2 and 3 have a basic surface charge and are thereby proposed to interact with nucleic acids,(16) whereas the SN4 domain has an acidic surface charge and is thereby less likely to be involved in nucleic acid interaction. Thus, the SN4 domain specific antibodies are less likely to disturb with nucleic acid interaction and could thereby be used in e.g. enrichment of Tudor-SN interacting RNA molecules by immunoprecipitation.
All of the obtained clones were highly specific in recognizing the endogenous Tudor-SN from HeLa cell extract using Western blot (Fig. 1B). Treatment of cells with siRNA against Tudor-SN significantly decreased the signal in Western blot analysis, demonstrating the specificity of the antibodies (data not shown). The anti-SN4 clone α-SN4-3 resulted in the highest level of immunoprecipitation (Fig. 1C), suggesting that the clone is especially suitable in recognizing the native full-length Tudor-SN protein.
The Tudor-SN protein sequence is highly conserved between mammals. For example, human, mouse, rat, and cow Tudor-SN protein sequences are over 95% identical to each other, and the antibodies are thus likely to recognize the Tudor-SN protein of all of the mammalian species. The clones α-SN4-1, α-SN4-4, and α-SN4-5 also recognized the mouse Tudor-SN (data not shown). One of the clones, α-SN4-4, recognized Tudor-SN of the fruit fly (D. melanogaster) (Fig. 2A, B). The identity of the detected protein was confirmed to be Tudor-SN, since knockdown of the protein by dsRNA in the S2 fly cell line decreased the signal in both Western blot analysis and immunocytochemistry (Fig. 2A, B). The α-SN4-4 clone also recognized a protein in zebrafish (D. rerio) extract that corresponds to the size of Tudor-SN (Fig. 2C).

α-SN4-4 MAb recognizes the Drosophila melanogaster (fruit fly) and Danio rerio (zebrafish) Tudor-SN. (
Antibodies were also used in immunocytochemistry to analyze the intracellular localization of Tudor-SN in HeLa cells. All of the clones gave a signal in cytoplasm (Fig. 3) and, with the exception of clone α-SN4-1, also in the nucleus. Intriguingly, one of the antibody clones, anti-SN4 clone α-SN4-3, gave a distinct dotty signal in the nucleolus. The differences in intracellular localization of Tudor-SN obtained with the different antibody clones could be explained by the multifunctional nature of the Tudor-SN protein. The protein may adopt different conformations in different protein complexes and some epitopes may be shed or exposed by protein interactions. The signals obtained by immunocytochemistry were specific since decreasing the cellular Tudor-SN levels by siRNA treatment significantly decreased the signal in immunocytochemistry (data not shown).
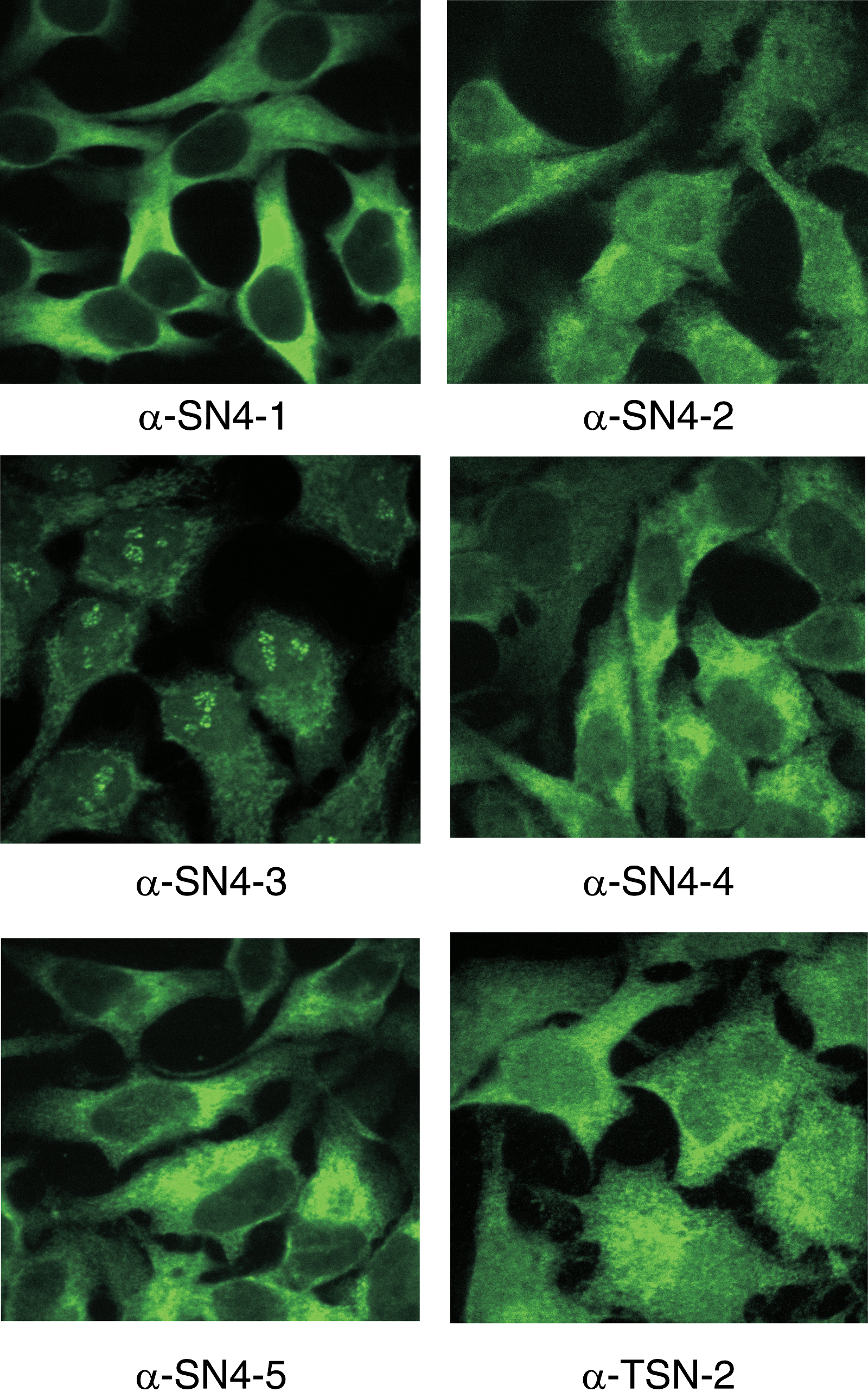
Immunocytochemistry of HeLa cells using the Tudor-SN antibodies. HeLa cells were paraformaldehyde fixed and immunocytochemistry was carried out using the indicated antibodies.
An extensive expression analysis of Tudor-SN was performed by human tissue arrays. Most of the tissues showed moderate to strong cytoplasmic signal in the immunohistochemistry using the MAb clone α-SN4-1. However, in both skeletal and heart muscle myocytes the signal was weak or undetectable (Fig. 4). In lymphoid organs moderate to strong signal was detected in spleen, lymph node, and tonsils. Signal was also strong in bone marrow. Especially high signal was detected in epithelial cells of many endocrine and exocrine tissues. In brain samples, Tudor-SN was detected in both glial cells and neurons of cerebral cortex and hippocampus and also in Purkinje cells of cerebellum. The antibodies created and characterized here will serve as important tools in approaches to understand the physiological role and functions of the Tudor-SN protein. The antibodies may also serve as a tool in cancer diagnostics.

Immunohistochemistry of selected human tissues. The anti-SN4 α-SN4-1 clone was used by the Swedish Human Proteome Resource (HPR) program (
Footnotes
Acknowledgments
We would like to thank Drs. Mataleena Parikka and Susanna Valanne for materials, and Paula Kosonen and Merja Lehtinen for technical assistance. This work was supported by the Medical Research Council of Academy of Finland, the Sigrid Juselius Foundation, the Medical Research Fund of Tampere University Hospital, the Finnish Cancer Foundation, the Tampere Tuberculosis Foundation, and the Finnish Cultural Foundation.