
Abstract
Myogenic determination 1 (MyoD) is a myogenic regulatory factor (MRF) possessing a basic domain and a helix-loop-helix domain. MRFs play a critical role in myoblast fate and terminal differentiation. MyoD is a transcriptional factor that induces transcription by binding with gene regulatory factors expressed in skeletal muscle. As a master gene, MyoD also determines skeletal muscle differentiation. In this study, we established a monoclonal antibody specific for MyoD using the rat medial iliac lymph node method. Immunoblot analysis revealed that our monoclonal antibody against MyoD could identify full-length MyoD. Moreover, immunocytochemical staining revealed a change in the expression of MyoD at the skeletal muscle differentiation stage. This monoclonal antibody against MyoD allows for further studies to elucidate the mechanism by which MyoD influences skeletal muscle differentiation.
Introduction
In addition to MyoD, MRFs include Myf5, myogenin, and MRF4.(2) MRFs activate transcription by binding with the E-box sequence in the gene expression regulatory region in skeletal muscle cells,(4,5) and are thereby involved in myoblast fate and differentiation.(6–10) MyoD was isolated as an MRF that transcribes 10T1/2 fibroblasts into muscle.(11) Forced MyoD expression induces the differentiation of various cells into myoblasts.(12) Therefore, MyoD functions as a master gene that determines skeletal muscle differentiation. Moreover, muscle differentiation in MyoD- and Myf5-deficient mice is inhibited at an early stage of development.(13) Myf5 deficiency does not affect MyoD expression, but deficiencies in Myf5 and Pax3 result in the repression of MyoD expression.(14) The regulation of MyoD transcription is complex and remains to be fully elucidated.(15) MyoD has 48% homology with Myf5; therefore, a specific antibody to elucidate the specific function of MyoD is in high demand. To examine the transcriptional regulation of MyoD, we established a monoclonal antibody specific for MyoD that can distinguish MyoD from Myf5.
Materials and Methods
Cell culture
Mouse skeletal myoblast C2C12 cells and rat myoblast L6E9 cells were cultured in growth medium (GM), Dulbecco's modified Eagle's medium supplemented with 20% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 μg/mL) (Nacalai tesque, Kyoto, Japan) under humidified atmosphere with 5% CO2 at 37°C. C2C12 and L6E9 cells were differentiated in differentiation medium (DM), Dulbecco's modified Eagle's medium supplemented with 2% horse serum (HS), with the addition of 25 ng/mL of IGF-1 to L6E9,(16) penicillin (100 U/mL), and streptomycin (100 μg/mL) (Nacalai tesque) under humidified atmosphere with 5% CO2 at 37°C.
Production and purification of recombinant proteins
An expression vector of glutathione S-transferase (GST) fusion proteins to full-length MyoD were transformed into Escherichia coli BL21 (DE3) (Novagen, Madison, WI).(17) The E. coli cells were grown in MMI medium containing 50 μg/mL calbenicillin (Nacalai tesque) at 37°C. Expression was induced by the addition of 0.5 mM isopropyl-1-thio-b-D-galactopyranoside (IPTG) (Nacalai tesque), followed by incubation for 12 h at 37°C. Lysed bacteria and purification of the fusion proteins were performed as previously described.(18)
Immunization of rat and production of MAb
Anti-MyoD rat MAbs were generated based on the rat lymph node method established by Sado and colleagues.(19–21) A 10-week-old female WKY/lzm rat (Japan SLC, Shizuoka, Japan) was injected via the hind footpads with 200 μL of an emulsion containing 100 μg of recombinant mouse MyoD protein and Freund's complete adjuvant. After 2 weeks, the cells from the lymph nodes of a rat immunized with an antigen were fused with mouse myeloma Sp2/0-Ag14 cells at a ratio of 5:1 in a 50% polyethylene glycol (PEG1500, Merck, Darmstadt, Germany) solution. The resulting hybridoma cells were plated onto 96-well plates and cultured in HAT selection medium (hybridoma SFM medium [Invitrogen, Carlsbad, CA]; 10% fetal bovine serum [FBS]; 10% BM-condimed H1 [Roche, Basel, Switzerland]; 100 μM hypoxanthin; 0.4 μM aminopterin; 1.6 μM thymidine). After 7 days post-fusion, the hybridoma supernatants were screened by means of an enzyme-linked immunoadsorbent assay (ELISA) against GST-fused MyoD. Positive clones were subcloned and rescreened by ELISA and immunoblotting. For the preparation of hybridoma supernatants containing highly concentrated antibodies, the resulting positive clone 5F11 was then cultured at a high density using a miniPERM bioreactor (Vivascience, Hannover, Germany).
ELISA
MyoD antigen (5 μg/mL) in TBS-T (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, and 0.05% Tween-20) was adsorbed on the surface of 96-well flexible microplates (Thermo Scientific, Rockford, IL) by overnight incubation at 4°C. To avoid unspecific binding, the plates were blocked with 1% bovine serum albumin (BSA) (Nacalai tesque) in TBS-T. Hybridoma supernatants were incubated for 1 h at room temperature (RT) and then washed with TBS-T three times. The plates were incubated for 30 min at RT with alkaline phosphatase-conjugated anti-rat IgG antibody (Sigma, St. Louis, MO) diluted 1:10,000. After washing with TBS-T three times, immunoreactivity was then visualized by a pNPP phosphatase substrate system (KPL, Gaithersburg, MD).(22)
Immunoblotting
C2C12 and L6E9 cells at growth phase and differentiated state were washed twice with phosphate-buffered saline (PBS) and then lysed in 2x SDS-PAGE sample buffer. The samples were separated by 10% SDS-PAGE, and electrophoretically transferred to a polyvinylidene difluoride membrane (Millipore, Bedford, MA). The membranes were blocked for 1 h at room temperature with a blocking solution containing 5% skim-milk (Nacalai tesque) in TBS-T (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, and 0.05% Tween-20), and then incubated for 1 h at 4°C with anti-MyoD rat MAb 5F11 diluted in Immunoreaction Enhancer Solution 1 (Toyobo, Osaka, Japan). After washing with TBS-T, the membranes were incubated for 30 min at RT with anti-rat IgG conjugated HRP (GE Healthcare, Buckinghamshire, United Kingdom). After washing with TBS-T, the membranes were detected by LAS3000 (Fuji Film, Tokyo, Japan).
Immunocytochemistry
C2C12 cells grown and differentiated on coverslips were washed twice in PBS at 37°C and fixed with 1% paraformaldehyde for 5 min at room temperature. After rapid washing in PBS, cells were permeabilized at 37°C for 5 min in 0.5% Triton X-100 in PBS, then incubated in blocking buffer as described.(23) MyoD was detected with 5F11, followed by an Alexa 488-conjugated goat anti-rat IgG (Invitrogen) for 1 h at RT. Coverslips were mounted with Prolong (Molecular Probes, Carlsbad, CA).(24) MyoD-expressing cells were visualized using a fluorescent microscopy (Olympus, Tokyo, Japan).
Results and Discussion
MyoD possesses a full-length 317 amino acid sequence. To establish a specific antibody against MyoD, we utilized a GST-fused full-length MyoD protein as an antigen. This sequence was confirmed to have homology with another MyoD, yet has no homology with other proteins. Using this protein, we established a monoclonal antibody against MyoD using the rat lymph node method. Enzyme-linked immunosorbent assay screening was performed with the domain, and 1 positive clone, 5F11, was obtained from the hybridoma. The isotype of this clone was IgG2a/k.
The deduced molecular weight of MyoD is 36 kDa. Utilizing the obtained positive clone 5F11, we performed an immunoblot analysis to evaluate reactivity against MyoD. Clone 5F11 bound to a protein that had a molecular weight of 36 kDa and was highly expressed in the cell extract of mouse skeletal muscle precursor cells C2C12. A weak band appeared at 36 kDa in rat skeletal muscle precursor cells L6E9, which have low MyoD and high Myf5 expression.(25) Therefore, 5F11 specifically identified MyoD. An additional band was also observed at 60 kDa, which may have been a result of a chemical modification of MyoD. Thus, 5F11 can specifically identify MyoD by immunoblot (Fig. 1).
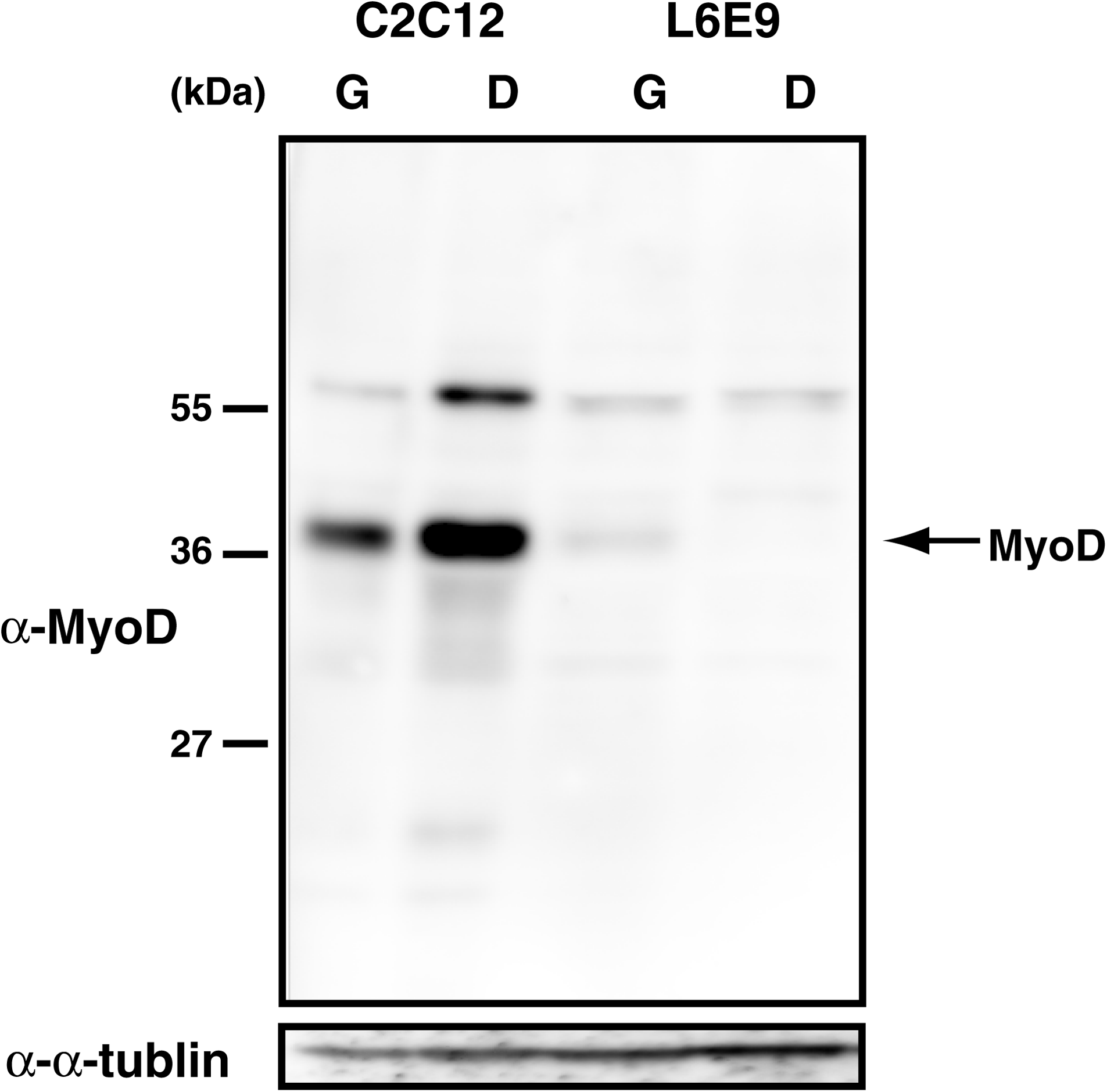
MyoD signal detected by monoclonal antibody 5F11. The specificity of the antibody was examined. Whole cell proteins of mouse C2C12 cells and rat L6E9 cells were separated on 10% sodium dodecyl sulfate polyacrylamide gels by electrophoresis, transferred to a polyvinylidene difluoride membrane, and immunoblotted with the MyoD antibody clone 5F11. G, cells in growth phase; D, differentiated cells.
MyoD protein localizes in the nucleus. Immunocytochemical staining using C2C12 cells was performed to evaluate if 5F11 could reveal the nuclear localization of MyoD protein. 5F11 staining showed that MyoD localized to the nucleus. Moreover, after the induction of differentiation in C2C12 cells, MyoD expression increased within 6 h, peaked at 24 h, and decreased within 48 h. Immunostaining of MyoD with 5F11 showed that MyoD expression was increased by positive feedback upon differentiation. Therefore, 5F11 can specifically identify MyoD in immunocytochemical staining (Fig. 2).
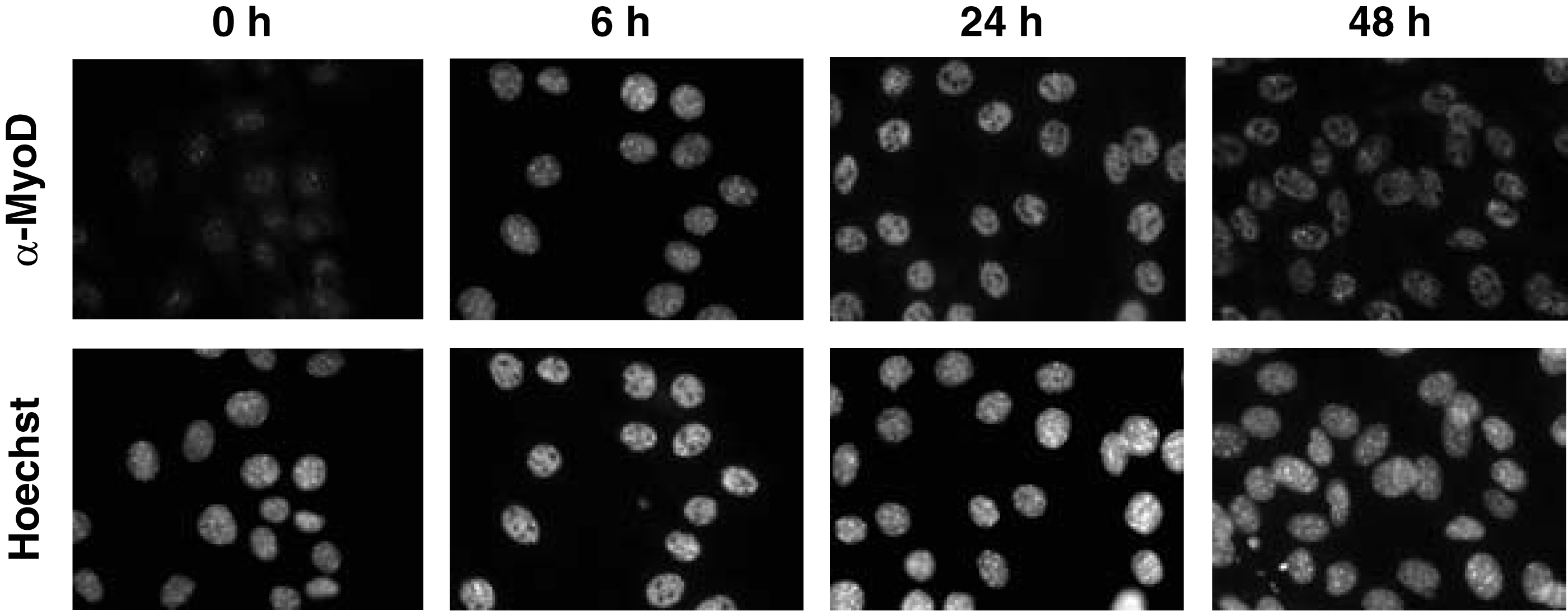
5F11 localized in the euchromatin of mouse C2C12 cells. C2C12 cells were cultured in growth medium for 1 day, then shifted to differentiation medium for 0, 6, 24, and 48 h. The cells were immunostained with the 5F11 clone. The antibody was detected with Alexa 488-conjugated anti-rat IgG. The sample was counterstained with Hoechst.
The use of monoclonal antibody 5F11, which is specific for MyoD, will contribute to further investigation of the regulation of MyoD expression.