
Abstract
Runx2 is a new transcription factor that takes part in odontoblast differentiation. This study is aimed at investigating the immunolocalization and expression of Runx2 in the process of dental pulp injury and repair using immunohistochemical technique. In normal dental pulp, positive staining can hardly be detected. In experimental groups, strong positive staining was detected at the site of the impaired pulp after 1 day, while only weak Runx2 staining was detected 3 days after operation. Five days later, a large number of stellate cells in the root apex expressed Runx2, and after 7 days, followed by the reparative dentinogenesis, Runx2 expression vanished slowly, then totally disappeared. Taken together, the expression of Runx2 has temporal and spatial specificity during different phases in rat tertiary dentinogenesis.
Introduction
The transcription factor Runx2 (runt-related transcription factor 2) is a key regulator of mesenchymal stem cell differentiation and plays an important role in dental germ differentiation,(7) dental tissue formation,(8) and the process of tooth eruption.(9) During the bud and cap stages, Runx2 is intense in all mesenchymal condensations of teeth.(10) However, Runx2 cannot be detected at the late bell stage.(11) Studies have confirmed that mutation of one Runx2 allele may cause cleidocranial dysplasia syndrome (CCD) in mice and humans.(12) CCD is an autosomal dominant disorder characterized by delayed skeletal development, large fontanelles, short stature, dental abnormity, and other changes in skeletal development and growth.(13–15) However, the function of Runx2 during the phase of tertiary dentin remains largely unknown.
In this study, experimental models of rat molar pulpitis were established. We have firstly investigated the immunolocalization and expression of Runx2 in the process of dental pulp injury and repair using immunohistochemical analysis.
Materials and Methods
Surgical procedure
A total of 18 Sprague-Dawley rats (200 g) were used in this study. All experimental procedures were conducted in accordance with the animal experimental guidelines of the Fourth Military Medical University (Xi'an, China). All steps were carried out at room temperature (18–22°C).
The animals underwent general anesthesia with pentobarbital sodium. The first and second maxillary molars were cleaned and disinfected with 75% ethanol. Pulpotomy was performed with a #1/2 round burr rotated by a low-speed electric engine. The size of the cavities was 2 mm in depth and 1.2 ∼ 1.3 mm in diameter. Bleeding was washed away with sterile saline. After using cotton pellets, cavities were sealed with glass ionomer cement (Fuji IX, Tokyo, Japan).
Section preparation
Each group was randomly divided into six additional groups. The animals were killed at 1, 3, 5, and 7 days and 2 or 3 weeks after treatment. The number of teeth analyzed in each group was 12. Each animal was fixed by intracardiac perfusion of paraformaldehyde. The first and second maxillary molars were dissected with the surrounding jawbones and post-fixed with 4% paraformaldehyde at 4°C for 24 h. The samples were then demineralized with 10% EDTA at 4°C for 3 weeks. The decalcified tissue blocks were then dehydrated in ascending series of ethanol, cleared in xylene, and embedded in paraffin. Samples were serially step-sectioned (5 μm) parallel to the long axis of the tooth.
Immunohistochemistry
After deparaffinization, paraffin sections were incubated with 0.3% H2O2 for 10 min to remove endogenous tissue peroxidase. Then sections were washed three times using 0.1 mol/L PBS, each time for 5 min. The sections were treated with normal rabbit serum for 30 min at 37°C. The sections were then incubated with the multiclonal antibody of rabbit-anti-mouse (1:100) in a moist chamber overnight at 4°C. The sections were treated with biotinylated secondary antibodies for 30 min at 37°C. Then sections were washed three times using 0.1 mol/L PBS and treated with streptavidin-perosidase for 30 min at 37°C. Finally, the sections were exposed to DAB until the color was developed, smeared with hematoxylin, cleared in xylene, and covered by general clarity gum.
Statistical analysis
Statistical differences were evaluated by paired t test. Significance was defined as p < 0.05.
Results
Hematoxylin eosin
For the control group, pulp tissue was composed of odontoblasts, free cell zone, cell-rich zone, and pulp core. Odontoblasts were columnar cells arranged in an epithelioid sheet along the junction between dentin and pulp. The microscopic images of the one-day group showed serous inflammation and many neutrophils around the exposure site. As for the 3-day group, the necrosis region appeared around the exposure site with many neutrophilic granulocytes and some mononuclear macrophages. Small vessels of the pulp were obviously extended; the quantity was larger and vacuolar degeneration was beginning to emerge. The inflammation of the pulp in the 5-day group was more serious, and coronal pulp was necrotic. The necrosis region of the 7-day group extended to radicular pulp with many phlogocytes. The proportion of the medium-sized lymphocyte and the fiber between the cells was greater. Several calcified deposits could be observed in the root canal wall. As for the 2-week group, there were many osteoid odontoblast-like hard tissues in the pulp chamber. The odontoblasts of radicular pulp were arranged normally, and the phlogocytes could not be obviously observed. In the 3-week group, reparatice dentin could be observed in the pulp chamber, while no phlogocytes could be seen in the pulp tissue. The diameter of the root canal became narrower because of the reparatice dentin.
Immunohistochemical staining
The expression of Runx2 was negative in the pulp tissue of the control group (Fig. 1). As for the 1-day group (Fig. 2), the Runx2 expression could mainly be detected around the exposure site, and strong positive staining of Runx2 was mainly distributed in the nucleus. Positive staining of Runx2 was also observed in cells around the small vessels. Runx2 expression could be weakly detected under the necrotic layer in the 3-day group (Fig. 3). The weak positive staining of Runx2 was distributed in the cells around the small vessels. In the 5-day group, the weak expression of Runx2 could be detected under the necrotic layer and also in the wall of the root canal (Fig. 4). Strong positive staining of Runx2 was distributed in astrocytes of the root apex. As for the 7-day group (Fig. 5), strong expression of Runx2 was detected in the predentin layer, while no Runx2 expression was detected in calcified nodules. Positive staining of Runx2 was less distributed in astrocytes of the root apex in the 2-week group, while no staining of Runx2 was detected in reparative dentin. As for the 3-week group (Fig. 6), a considerable amount of newly formed cementum led to canal blocking and Runx2 expression could barely be detected.
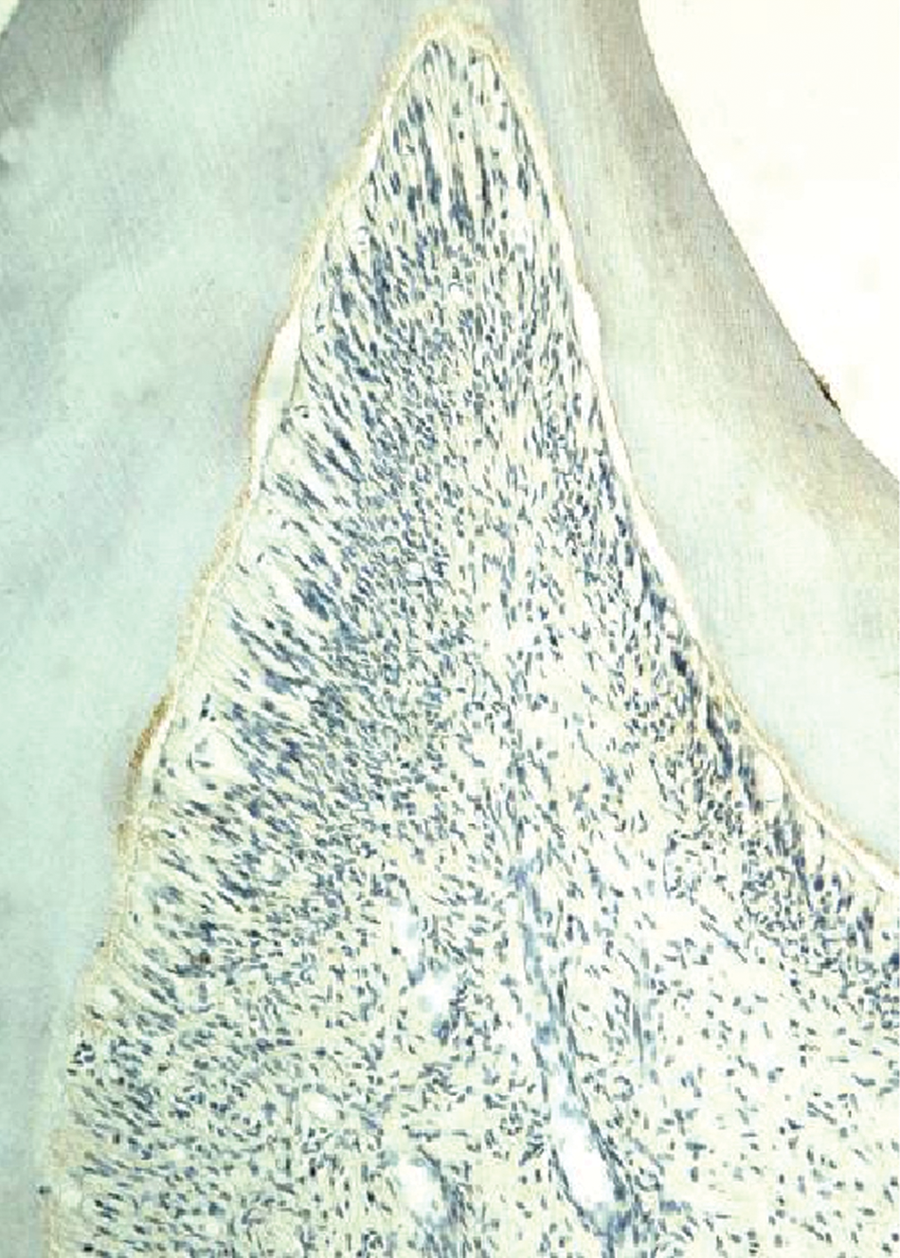
Expression of Runx2 in the pulp tissue of the control group.

Expression of Runx2 in the pulp tissue of the 1-day group.
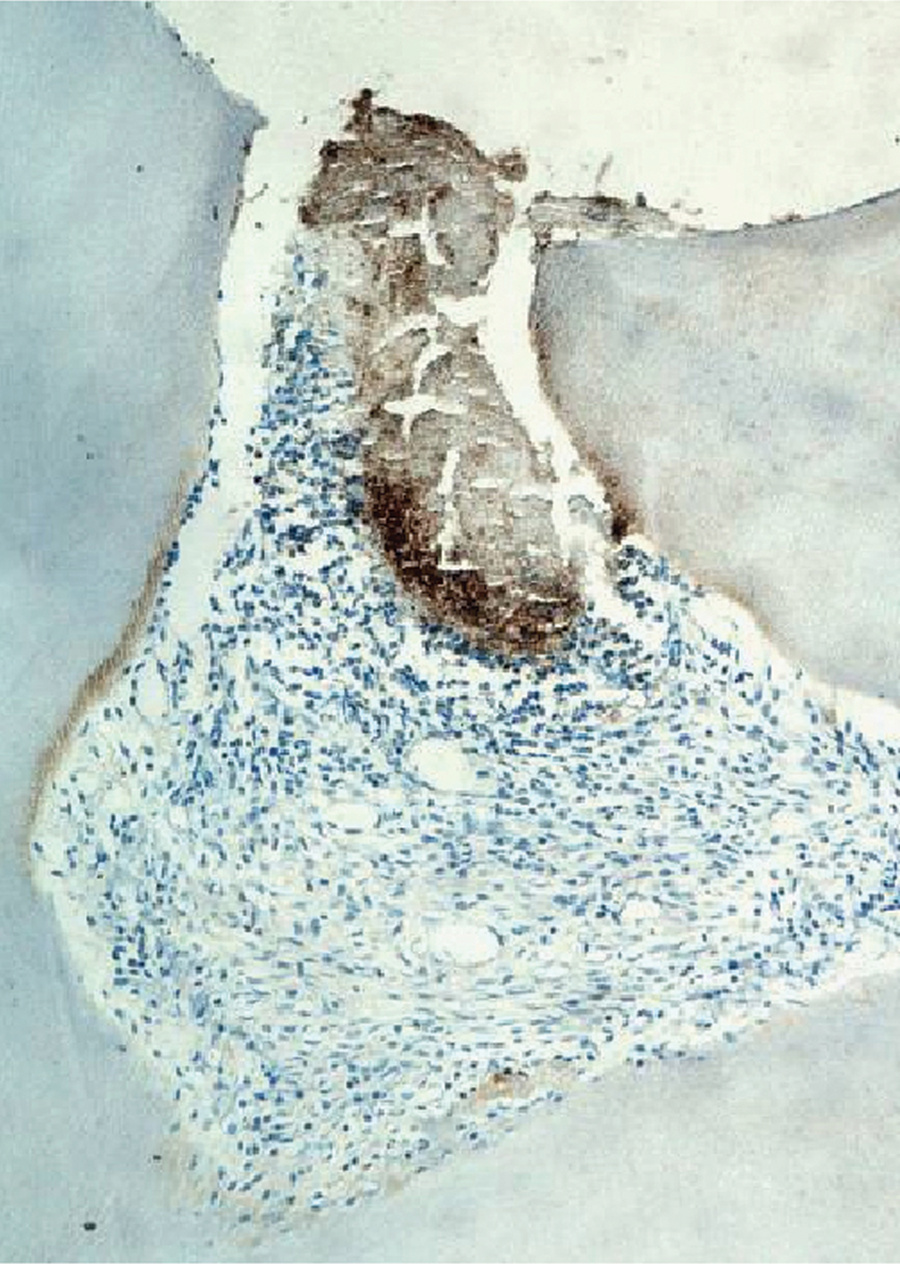
Expression of Runx2 in the pulp tissue of the 3-day group.

Expression of Runx2 in the pulp tissue of the 5-day group.
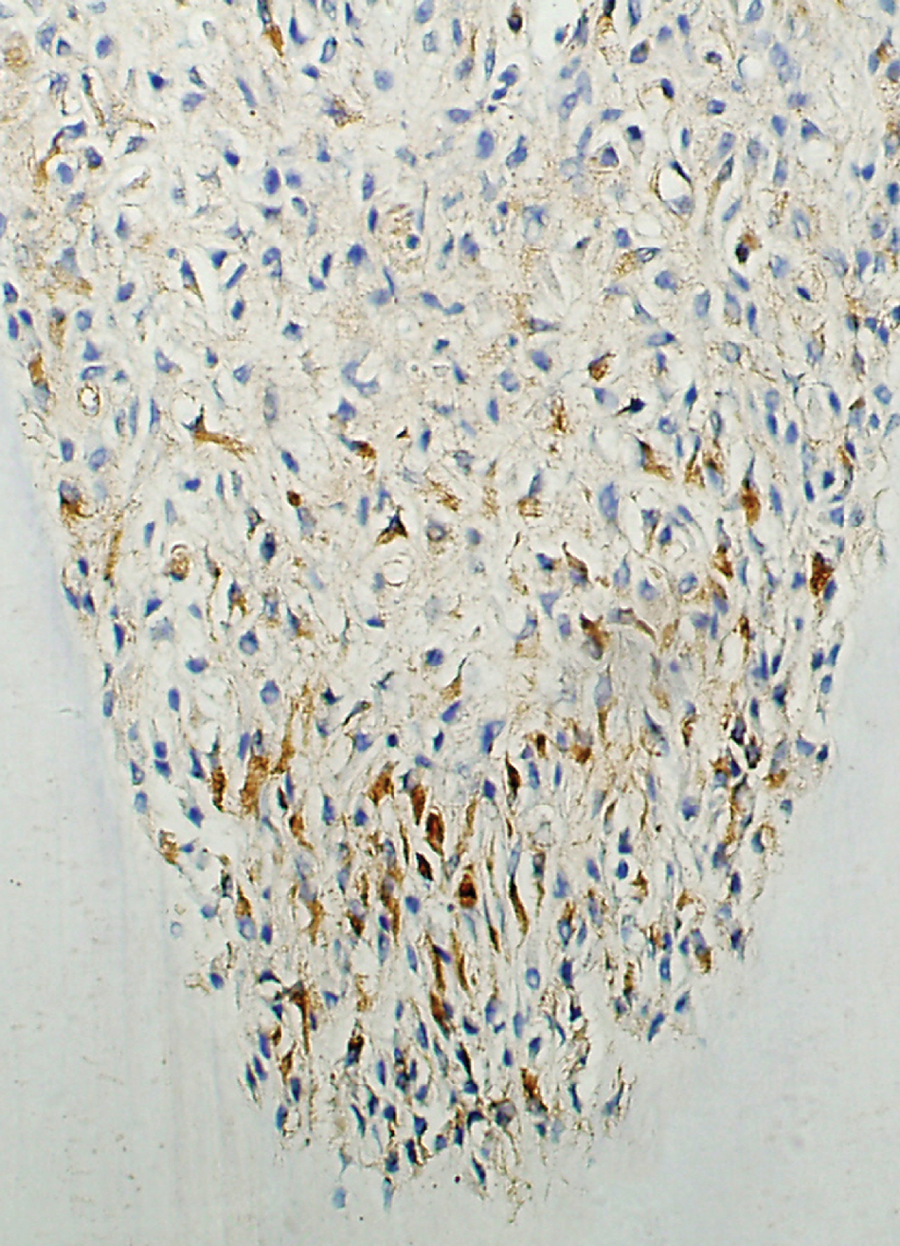
Expression of Runx2 in the pulp tissue of the 7-day group.
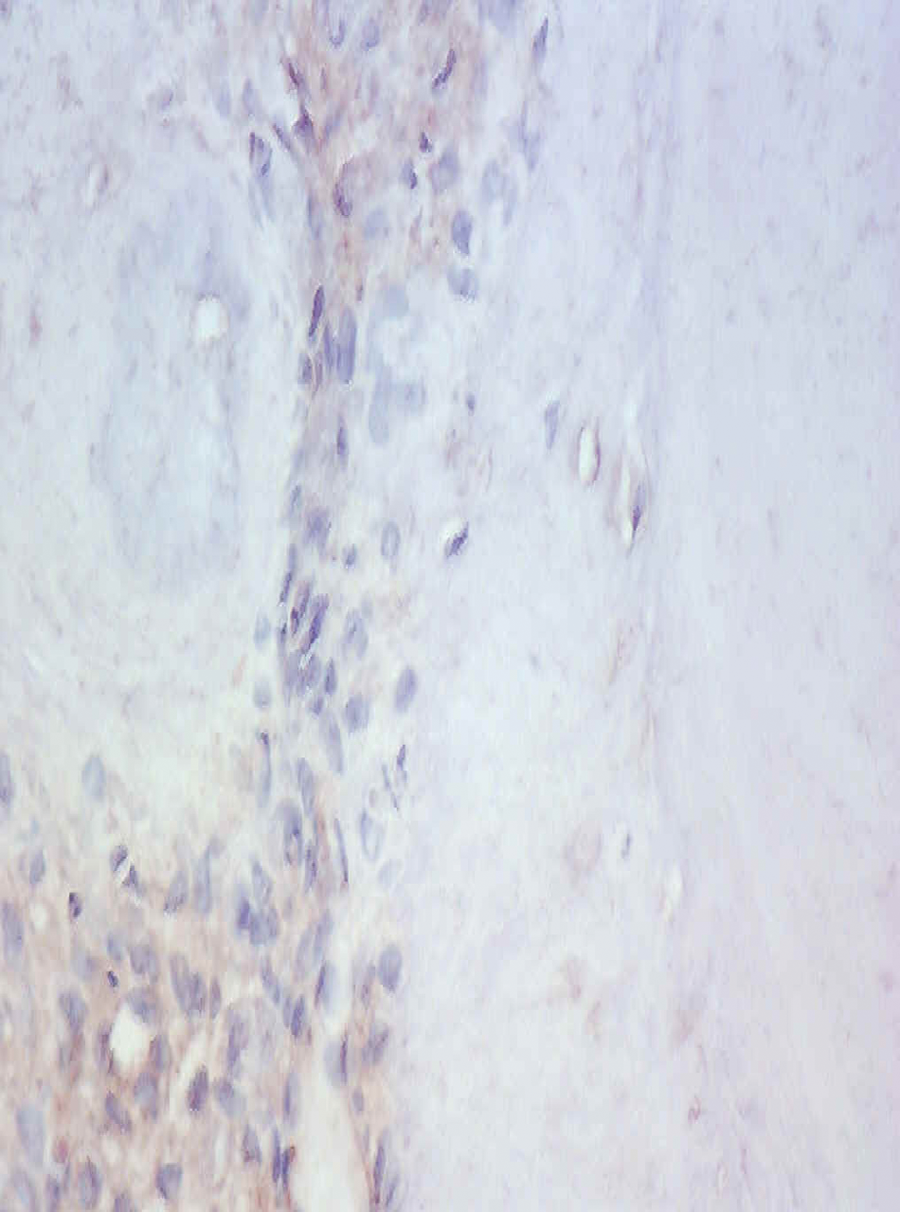
Expression of Runx2 in the pulp tissue of the 3-week group.
Discussion
Runx2 is identified as an osteoblast-specific transcription factor and a regulator of osteoblast differentiation.(16) The expression of Runx2 is important for the differentiation of bone marrow stromal cells, while dentin sialophosphoprotein contributes to the odontogenic differentiation.(17) Runx2 was intense in all mesenchymal condensations of bones and teeth. In early tooth morphogenesis, Runx3 was co-expressed with Runx2 in a thin layer of mesenchymal cells underlying the dental epithelium.(18) Studies suggest that Runx2 regulates key epithelial-mesenchymal interactions that control advancing morphogenesis and histodifferentiation of the epithelial enamel organ in odontogenesis.(10)
In this study, the data showed that the expression of Runx2 was strongly detected below the exposure site during the early phase of pulp injury, suggesting that Runx2 was involved in forming the reparative dentin. The pulp cells under the exposure site gradually necrosized by the third day. The undifferentiated cells at the root apex had an active function with a large number of satellite cells on the fifth day, suggesting that Runx2 might take an important role in the proliferation and differentiation of undifferentiated mesenchymal cells. On the seventh day, Runx2 expression vanished slowly, with the formation of the pulp root wall or the odontoblast-like tissue. So the influence of Runx2 was weaker during the final phase of reparative dentin formation.
Runx2 was also highly expressed in early stages of tooth development and was involved in crown morphogenesis and cytodifferentiation of odontoblasts. The expression of Runx2 was decreased in differentiated odontoblast, even barely detected. However, the expression was still low, especially in dental papilla cells at the pulp apex and also the odontoblasts along the root canal wall. So we deduced that Runx2 might play a stage-specific role in forming tertiary dentinogenesis.(19)
The differentiation of postnatal human dental pulp stem cells (DPSCs) was initially marked by up-regulation by Runx2, suggesting that the formation of reparative dentin was related to DPSCs and other factors.(20) This study suggested that the expression of Runx2 was strong in stellate cells of the root apex. However, the molecular mechanism of Runx2 expression during differentiation of DPSCs into ondontoblasts was still uncertain. We are performing more studies to clarify the function of Runx2 in pulp preservation for clinic treatment.
Footnotes
Acknowledgment
This work was supported by grants from the Foundation Shaanxi Science and Technique Development Program 2006-K14-G10920 and Chinese National Science Foundation Program 30870636.