
Abstract
The β8 integrin, which forms αvβ8 heterodimers, is being widely investigated because of its specific structure and functions compared with other integrins. In this report, a 12 aa-long peptide of β8 integrin cytoplasmic domain was synthesized according to a published sequence and covalently coupled to keyhole limpet hemocyanin (KLH). Three stable strains of hybridomas (3G6, 5C7, 5H3) that can secrete high specific monoclonal antibodies against β8 integrin were successfully established by hybridoma technique. The isotypes of these MAbs were tested to be IgG2a. Their characterizations were shown by enzyme-linked immunosorbent assay (ELISA), Western blot analysis, and immunocytochemistry (ICC). The affinity constants (Kaff ) of the MAbs 3G6, 5C7, and 5H3 were measured by non-competitive ELISA respectively. Western blot analyses and immunocytochemistry demonstrated that all the MAbs were directed against β8 integrin with high specificity.
Introduction
Integrin αv subunit is the exclusive partner of β8 subunit,(4) which is widely expressed in various cells and tissues; however, distribution of the counterpart β8 subunit varies greatly depending on cell type and developmental stage. At present, expression of β8 was reported to be restricted to brain, ovary, uterus, kidney, placenta,(3) lung,(5) eyelid,(6) and spleen.(7) Expression in brain, localized in dendritic spines, synaptic terminals, and glial processes,(4) is far greater than in other tissues; αv subunit and β8 subunit hold a similar function in the interactions between cerebral blood vessels and brain parenchymal cells. Most αv–/– or β8–/– mice died during embryogenesis as a result of placental defects.(8–10) Targeted deletion of αv or β8 integrin in both central nervous system glia and neuroepithelial cells, but not endothelial cells, results in neonatal vascular defects and intracerebral hemorrhage; some mutants survive for several months, but develop severe neurological phenotypes, leading to ataxia and paresis.(11,12) In the other tissues, kidney β8 integrin is localized in normal glomerular mesangial cells, and integrin β8–/– mice exhibit myofibroblast(13); β8 integrin is expressed in airway epithelial cells but is absent in lung cancer, suggesting a role in epithelial homeostasis(5); β8 integrin plays a role in the initial merger of the epithelial tissues of the eyelid and stabilization of the epithelial bridge(6); β8 integrin is important for negative regulation in peripheral tissues, preventing immune dysfunction that results in autoimmunity.(7)
In this study, we used synthetic peptide coupled to KLH eliciting antibodies against a conserved epitope from the cytoplasmic domain of β8 integrin. A 12 aa-long peptide selected from the sequence of β8 integrin cytoplasmic domain was synthesized and covalently coupled to carrier proteins for the preparation of immunogen and coating antigen. The MAbs against β8 integrin were prepared by hybridoma technique. Furthermore, the expression of β8 in cells and tissues were analyzed by immunocytochemistry and immunohistochemistry.
Materials and Methods
A 12 aa-long peptide (EEIKMDISKLNA) selected from the cytoplasmic domain of β8 integrin was synthesized by Sbsbio (Beijing, China) and conjugated to KLH using Imject KLH carrier protein and EDC conjugation kit (Pierce, Rockford, IL) according to the manufacturer's instructions. One hundred μg β8-KLH emulsified in complete Freund's adjuvant (CFA, Sigma, St. Louis, MO) were injected subcutaneously in two 8-week-old Balb/c mice and boosted twice with the same quantity of antigen in incomplete Freund's adjuvant (IFA, Sigma) at 2-week intervals. A small volume of blood was collected from the tail vein for testing the antibody titer. A final boost of 100 μg antigen was injected intraperitoneally (i.p.) without adjuvant 3 days before harvesting of splenocytes for cell fusion. Three days after the final booster injection, splenocytes from immunized animals were harvested and fused to the HAT-sensitive murine myeloma cell line SP2/0 with polyethylene glycol 4000 (Sigma) at a splenocyte-myeloma cell ratio of 10:1. Then cells (hybridomas) were distributed in 96-well culture plates, which were supplemented with HAT medium (Sigma) for selection, containing peritoneal macrophages as feeder cells from young Balb/c mice.
Culture supernatants were harvested for screening by indirect ELISA after incubating for 2 weeks: β8–KLH conjugate (100 μL/well in 0.05 mol/L carbonate buffer [pH 9.6]) were bound to 96-well microtiter plates overnight at 4°C. The plates were washed three times with PBST and then blocked with 1% casein in PBS (120 μL/well) for 1 h at 37°C. After washing the plates three times with PBST, 70 μL/well of hybridoma supernatants were pulsed into wells and incubated at 37°C for 2 h. After washing with PBST, 100 μL of HRP-GaMIgG (ZSBio, Beijing, China) diluted in PBS was added to each well, and incubated for 1 h at 37°C. After another washing step, the substrate solution (100 μL/well) was added. The reaction was stopped with sulfuric acid (80 μL/well) after 15 min and the absorbance was read at 450 nm using an ELISA reader (Bio-Rad, Hercules, CA).
Positive mother hybridomas for β8 were cloned three times in 96-well plates by limiting dilution (calculated as 0.75 cell/well to ensure single cell in most of the wells) in RPMI 1640 (Gibco, Paisley, United Kingdom) containing 10% FBS (Minhai, Lanzhou, China). Supernatants of growth-positive wells were screened for antibodies against β8 by indirect ELISA.
The pristane primed Balb/c mice were injected intraperitoneally with 1.0 × 106 hybridoma cells per mouse. The ascites was collected after successful ascites development (10–14 days) and purified by ammonium sulfate precipitation. Isotypes were determined using a BD Bioscience isotyping kit (Heidelberg, Germany).
The affinity constants of MAbs against β8 integrin were determined by noncompetitive ELISA, as described by Beatty and colleagues.(14) Briefly, the ELISA plates were coated with antigen diluted to concentrations of 2.0, 1.0, and 0.5 μg/mL. After being blocked, the plate was added with MAbs against β8 integrin in a series of diluted concentrations and incubated for 2 h at 37°C. The following steps were performed as mentioned above and the optical absorbance at 450 nm was measured. The sigmoid curve was plotted to represent the relationship of A450 value versus logarithm of MAbs. The affinity constant of antigen-antibody interaction can be estimated according to the following equation:
where (Ag)t and (Ag′)t are the total antigen concentrations in the wells, (Ab)t and (Ab′)t are the total antibody concentrations in the wells at OD-50 for plates coated with two different concentrations of antigen, respectively.
Fresh mouse kidneys were fixed in 4% paraformaldehyde overnight at 4°C and then embedded in paraffin. Cut sections of 5 μm were affixed onto the 3-amino-propyl-tri-ethoxy-silane (APTES) coated slides and baked. The slides were then deparaffinized and rehydrated. For immunocytochemistry, 1 × 105 astrocytes were plated on glass coverslips in 6-well plates and cultured overnight, then fixed in 4% paraformaldehyde and baked. The following steps for immunohistochemistry and immunocytochemistry are nearly the same. Sections and cells were heated up in antigen retrieval buffer(15) and permeabilized using 0.1% Triton X-100. Endogenous peroxidase was blocked with 0.3% H2O2; then the cells were blocked in 10% goat serum in PBS. Primary monoclonal antibody (1:100) was added at 4°C overnight. The primary antibody was omitted in control cells. They were then incubated with biotin-labeled goat anti-mouse IgG (ZSBio) at a 1:1000 dilution. Horseradish peroxidase (HRP)-conjugated streptavidin (ZSBio) was added at a 1:2000 dilution and 3,3′-diaminobenzidine (DAB) was added to stain. Cells were restained with hematoxylin, then sealed with neutral balsam. Images were acquired by a digital imaging system (Olympus, Tokyo, Japan).
Lysates of fresh mouse brains were prepared in lysis buffer (50 mM Tris-HCl [pH 7.5], 150 mM NaCl, 0.1% deoxycholate, 0.1% SDS, 1% Triton X-100, 300 mg/mL PMSF, 1 mg/mL pepstatin A, 2 mg/mL aprotinin, and 4 mg/mL leupeptin). Samples were electrophoresed on 12% SDS-PAGE gels with prestained markers (Fementas, Burlington, Canada), then transferred to a polyvinylidene difluoride membrane (Millipore, Billerica, MA). Membranes were blocked with 3% BSA in PBST (4°C, overnight) and MAb to β8 was added at a 1:1000 dilution (4°C, overnight). After washing with PBST, HRP-conjugated goat anti-mouse antibodies were added at a 1:2000 dilution (37°C, 1 h), followed by further washes. Blots were developed with enhanced electrochemiluminescence (ECL, Bio-Rad). The size of each band was determined based on the molecular weight of the protein standard.
Results and Discussion
A 12 aa-long peptide selected from the sequence of the cytoplasmic domain of β8 integrin, which was highly conserved among different species of mammals, was synthesized and coupled to KLH as immunogen and coating antigen.
Two Balb/c mice were immunized with β8 integrin–KLH. The specific antibody titer during immunization was monitored by analyzing blood samples using an indirect ELISA. After the third injection of antigen, it was observed that antisera collected from two immunized mice displayed high affinity binding with coating antigen; the titers of the antibody against the antigen in mice sera reached up to 1:1.024 × 105 (Fig. 1A). Three days after the final booster injection, the splenocyte fusion experiment was carried out. After 2 weeks incubation, the supernatants from hybridoma cells were screened by an indirect ELISA. There was no cross reactivity to β actin–KLH. Supernatants of β8 integrin positive wells were subcloned three times by limiting dilution. Three stable monoclonal hybridoma cell lines of 3G6, 5C7, and 5H3, which could constitutively produce specific MAbs to β8 integrin, were successfully established. Monoclonal antibodies were then produced in mouse ascites. The titers of the MAb clones 3G6, 5C7, and 5H3 were 1:4.096 × 105, 1:4.096 × 105, and 1:1.024 × 105, respectively. After being purified by saturated ammonium sulfate precipitation, the concentrations of the MAbs 3G6, 5C7, and 5H3 were 5.482, 3.566, and 4.702 mg/mL, respectively.
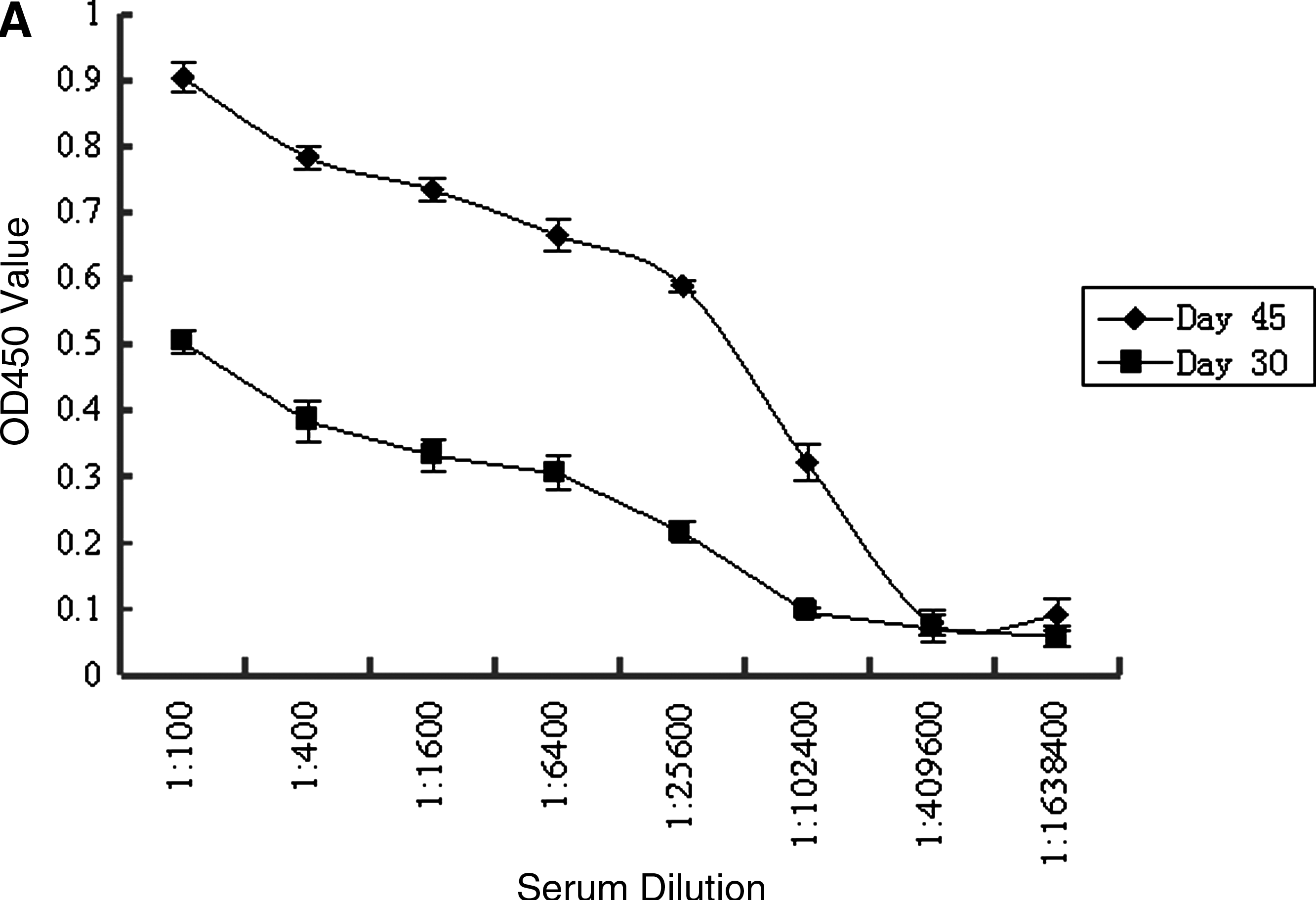
(
All β8 integrin MAbs were determined to be IgG2a by the Isostrip kit. The affinity constants (Kaff ) of MAbs against β8 integrin were measured by non-competitive ELISA as described earlier. The result and curve are shown in Table 1 and Fig. 1B. Kaff of MAbs 3G6, 5C7, and 5H3 to β8 integrin were 3.504 × 108 M−1, 3.327 × 108 M−1, and 2.964 × 108 M−1, respectively.
The specificity of the monoclonal antibodies was tested by Western blot analyses. All three clones are reactive particularly with the lysates of fresh mouse brains (Fig. 1C). The single band was shown to be consistent with the molecular mass (∼95 kDa) of β8 integrin, which was determined based on the pre-stained molecular weight of the protein standard. Furthermore, the reactive intensity of clones 3G6 and 5C7 with cell lysate is stronger than that of 5H3.
Immunohischemistry and immunocytochemistry were performed to further confirm the specificity of the anti-β8 integrin antibodies. We found β8 integrin was expressed primarily in the membrane and cytoplasm of renal proximal tubular epithelial cells, whereas it was negative in glomeruli. A positive reaction gradually declined from the cortex to the medulla (Fig. 2). Meanwhile β8 integrin was also expressed in the membrane, cytoplasm, and neurites of astrocytes while no positive immunoreactivity in negative controls was observed (Fig. 2). These results confirmed the specificity of the anti-β8 integrin antibodies in detecting the β8 integrin in tissues and cells.

The specificity of monoclonal antibodies analyzed by immunohistochemistry in the kidneys and immunocytochemical in the astrocytes. Paraffin-embedded sections were stained with anti-β8 integrin monoclonal antibodies (3G6) at a 1:100 dilution (
The cytoplasmic domains of β8 integrin subunit is structurally unique to the other β subunits, indicating β8 cytoplasmic domains perform distinct functions.(3) Generally, integrins play an important role in attachment of the cell to the ECM. However, β8 integrin does not appear to be able to promote cell adhesion because of its divergent cytoplasmic domain, though vitronectin of ECM is a ligand for αvβ8.(16) Furthermore, β8 cytoplasmic domain is sufficient to induce both cell growth inhibition and cause cell shape changes.(5)
Cytoplasmic domains of β integrins except β4 and β8 mediate outside-in or inside-out signal transduction via FERM domain of talin.(17) β8 integrin cytoplasmic tail lacks talin binding sequences, such as NPXY or NPXY-like motifs,(18) suggesting that β8 integrin activation and signal transduction may be mediated via distinct mechanisms from other integrins. In CNS, band 4.1B interacts with β8 integrin via the band 4.1B C-terminal domain instead of its FERM domain to influence αvβ8-mediated function.(19) In addition, β8 integrin has a defined role for maintaining brain vessel homeostasis through regulation of latent forms of TGF-β activation that influence the expression of anti-angiogenic molecules.(20) A similar result was that integrin αvβ8 mediates cell growth inhibition via membrane type 1(MT1)-matrix metalloprotease (MMP)–dependent activation of TGF-β1.(21) In the kidney, β8 integrin stimulation by multiple pathways results in interaction between β8 and guanine nucleotide dissociation inhibitor-1, Rac1, and Cdc42 activation but not RhoA activation. Rho, Rac, and Cdc42 belong to the Rho subfamily; RhoA activation induces stress fiber and focal adhesion formation, whereas activated Rac1 and Cdc42 prevent these processes. Thus β8 integrin activation leads to suppression of pathological mesangial cell features.(13) Moreover, β8 integrin can be upregulated by Fas through an apoptosis-independent process to mediate pathophysiological functions of renal tubular epithelial cell.(22)
In this study, we used synthetic peptide coupled to KLH eliciting antibodies against a conserved epitope from cytoplasmic domain of β8 integrin, which was different from expression protein eliciting antibodies in eptiope.(5) A 12 aa-long peptide of β8 integrin cytoplasmic domain was synthesized and coupled to carrier proteins. Monoclonal antibody against β8 integrin was prepared by hybridoma technique. Since the 12 aa-long peptide was conjugated to KLH, choosing the β-actin coupled to KLH as the negative control in the screening assay could effectively eliminate false-positive MAbs to KLH. The results proved that the method was efficacious, and the Western blot analyses showed that the three MAbs obtained had no reactivity with KLH. Moreover, three hybridoma cell lines secreting MAbs against the β8 integrin were generated and identified with high affinity, stability, and specificity.
Footnotes
Acknowledgment
This work was supported by the staff of the Department of Immunology, School of Preclinical and Forensic Medicine, West China Medical Centre, Sichuan University, China.