
Abstract
Class IV chitinase, an allergenic protein of Vitis vinifera (grape), was purified by anion exchange chromatography and used for immunization of Balb/c mice. Monoclonal antibodies (MAbs) were raised by hybridoma technology using Sp2/0 myeloma cells. Finally after three limiting dilutions, six stable clones were generated. Antibody isotyping showed that IgG2a, IgG2b, and IgM were produced by one, two, and three of the clones, respectively. All of the MAbs had kappa light chain. The affinities were in the range of 3 × 108 to 1.2 × 109 M−1. The MAbs were specific for grape chitinase as confirmed by Western blotting. In conclusion, we successfully produced several MAbs against grape class IV chitinase, which could be used for assessment of this allergen in different grape cultivars.
Introduction
Grapes are among the allergenic fruits that are highly produced and consumed all over the world. Several pathogenesis-related (PR) proteins, including chitinases, are expressed in different stages of grape berry development, which some studies have found to be allergenic.(15,16)
Chitinases are glycosyl hydrolases that catalyze degradation of chitin, a homopolymer of N-acetyl-D-glucosamine.(17) Many organisms possess chitinolytic enzymes that make them capable of degrading chitin fibrils. Depending on the organism type, these enzymes show different functions. In bacteria and fungi, the chitinolytic enzymes may have a nutritional profit by exploiting chitin, the second abundant bio-polymer of the nature, as a source of carbohydrate.(18) These enzymes were also shown to play a role in fungal morphogenesis.(19) Plants express several PR proteins such as chitinases, mostly in the course of natural defense reactions against invasion of pathogenic microorganisms. The antifungal defense mechanism of plant chitinases may be explained by direct damage of the cell wall and indirect release of chitin oligomers, which can elicit plant defensive responses.(17)
Endochitinases are the main chitinolytic enzymes of the plants and were extensively studied.(20) They are classified according to their structure, enzymatic property, and subcellular localization. Previous studies show that class IV chitinase is one of the major allergens of grapes,(16) and it is highly expressed during grape verification and ripening time.(21) Class IV chitinases belong to PR-3 proteins and are categorized in the family 19 of glycosyl hydrolases. They have a molecular mass of around 28 kDa and are characterized by the presence of an N-terminal hevein-like region or chitin-binding domain.(22)
Characterization and assessment of allergenic proteins may be useful in the determination of the allergenicity potential of foods and pollens. Several techniques including functional and proteomic methods have been applied for these proposes. But the simplicity and availability of enzyme immunoassays keep them more popular. PR proteins such as chitinases are among the major allergens of fruit, and assessment of their quantity in fruit extracts could be useful. Considering that the starting step for immunodetection of these proteins is production of specific monoclonal antibodies (MAbs), anti-chitinase MAbs were produced and characterized in this experiment.
Material and Methods
Vitis vinifera (cultivar sultana) was collected from the Golmakan vineyard of Iranian Ministry of Agriculture. Chromatography columns and equipment were obtained from Pharmacia (Uppsala, Sweden). Balb/c mice and Sp2/0 Ag-14 (National Cell Bank of Iran, code C129) were obtained from the Pasture Institute of Tehran, Iran.
Cell culture flasks and plates were obtained from Orange Scientific (Braine-l'Alleud, Belgium). Cell culture media, antibiotics, and fetal calf serum (FCS) were purchased from Invitrogen/Gibco (Grand Island, NY). Biotinylated rabbit anti-mouse antibody and horseradish peroxidase (HRP)-conjugated streptavidin were obtained from KPL (Gaithersburg, MD). HT and HAT media and other chemicals were obtained from Sigma-Aldrich (St. Louis, MO). Mouse monoclonal isotyping kit was purchased from Serotec (Oxford, United Kingdom).
Preparation of grape crude extract and purification of class IV chitinase
Grape total proteins were extracted by the Bjorksten method, with some modification.(23) Briefly, grape berries were ground using a fruit juicer and homogenized with the same amount of 0.1 M potassium phosphate buffer (pH 7.0) containing 20 mM EDTA, 5% w/v polyvinylpolypyrrolidone (PVPP). The mixture was shaken for 6 h and centrifuged at 9000 g for 20 min, and the clear supernatant dialyzed against 10 mM potassium phosphate buffer (pH 8.0) and freeze-dried. The major proteins of the crude extract were separated by ion-exchange chromatography. Briefly, diethylaminoethyl (DEAE) sepharose 6B was packed in a column and pre-equilibrated with 20 mM Tris-HCl (pH 8.8). The dialyzed extract was loaded onto the column, and the proteins were eluted by starting buffer containing increasing amounts of NaCl. Effluents were monitored at 280 nm by an LKB Uvicord (Pharmacia) absorptiometer. The protein fractions were collected manually and freeze-dried. The purity of the fractions was determined by silver staining. The purified proteins were analyzed by MALDI-TOF/TOF mass spectrometry (ABI 4800, Applied Biosystems, Foster City, CA) using GPS Explorer Workstation, v. 3.6 (Applied Biosystems) and MASCOT, NCBInr software as previously described.(56) Finally, fractions containing class IV chitinase were used for sensitization of the mice.
Immunization and preparation of monoclonal antibody
Two young female BALB/c mice were immunized with purified chitinase. Briefly, 40 μg of purified protein were injected as the first step after emulsification with complete Freund's adjuvant. The immunization followed by three intra-peritoneal injections of 20 μg of protein with incomplete Freund's adjuvant in 2-week intervals. Anti-chitinase antibody titer was determined by indirect enzyme-linked immunoadsorption assay (ELISA). The mouse with the higher titer of antibody (OD450nm = 0.92 at 1:128,000 dilution) was selected and injected with 20 μg of antigen intravenously, 3 days prior to cell fusion. The splenocytes of the immunized mouse were fused with Sp2/0 Ag-14 myeloma cells, according to the previously described procedure.(24) The fused cells were incubated with HAT media. After 2 weeks, the supernatants were screened by in house–designed indirect ELISA.
The highly reactive wells were selected for limiting dilution, and after three cloning procedures, the positive ones were nominated and selected for further studies. The isotype of the selected MAbs was determined according to the manufacturer's instruction.
Antibody purification
Protein-A column chromatography was used for purification of monoclonal antibodies. Briefly, columns with 2.5 mL of protein-A sepharose gel were packed and equilibrated with PBS (pH 7.4). Ten mL of fresh supernatant from each IgG producing line were loaded on an appropriate column. The unbound proteins were removed by several washings of the column with PBS. Finally, IgG was eluted by adding 0.2 M glycine-HCl buffer (pH 3.0). The collected fractions were neutralized with 1 M Tris-HCl (pH 8), dialyzed against PBS, and used for determination of affinity by the Beatty method.(25)
Indirect ELISA
To measure the titer of specific antibody in mice sera and determine the secretory state of the hybridoma cells, an indirect ELISA was developed. Briefly, microtiter plates were coated with 2 μg of purified protein in 0.1 M bicarbonate buffer (pH 9.5) at 4°C for overnight. Each well was then blocked for 2 h at 37°C with 300 μL of 2% bovine serum albumin (BSA) in phosphate buffer saline (PBS), and then washed five times with PBS containing 0.05% Tween-20 (PBS-T). Then 50 μL of diluted mice sera or supernatants of hybridoma cells were added to the wells and the plates were incubated for 3 h. After washing, 50 μL of 1:3000 diluted biotinylated rabbit anti-mouse antibody were added to each well and the plates were incubated for 1 h. After another washing step, 1:30,000 diluted enzyme-conjugated streptavidin was added to the wells and the plates were incubated for 45 min. The plates were washed again and chromogenic substrate (3,3',5,5'-tetramethylbenzidine + H2O2) was added to each well and incubated for 15 min in the dark. Color development was stopped by addition of 50 μL of 3 M HCl. The absorbance was measured at 450 nm with an ELISA reader. All of the dilutions (including mouse serum, cell culture supernatants, anti-mouse antibodies, and enzyme-conjugated streptavidin) were prepared in 1% BSA. All of the incubations except for the coating step were carried out at 37°C on an orbital ELISA shaker.
Gel electrophoresis and Western blot analysis
Electrophoresis of grape proteins or purified fractions in the presence of sodium dodecyl sulfate (SDS) was carried out using a Bio-Rad Mini PROTEAN tetra cell vertical slab gel electrophoresis system (Bio-Rad, Hercules, CA) on 15% polyacrylamide gel as described by Laemmli.(26) The separated proteins were transferred on a polyvinylidene difluoride (PVDF) membrane at 300 mA for 15 min.(27) The membrane was cut into strips, blocked with 2% BSA for 12 h, washed with PBS-T, and incubated 4 h at 37°C with 1:5 diluted supernatant of monoclonal antibodies. The strips were washed again and incubated with 1:1000 diluted biotinylated rabbit anti-mouse antibody for 2 h at 37°C. To detect bound biotinylated antibodies, we added 1:30,000 diluted HRP-conjugated streptavidin and incubated the plates for 45 min. After a final vigorous washing, the reactive bands were visualized by Supersignal West Pico Chemiluminescent Substrate (Pierce, Rockford, IL), according to the manufacturer's instruction. Chemiluminescent signals were captured by SynGene chemi-documentation system (GBox Chemi HR, Cambridge, UK) using GeneSnap, v. 6.08 software (Synaptics, Cambridge, UK).
Results
The SDS-PAGE analysis of grape crude extract and fresh grape juice showed that a 28 kDa protein was highly expressed in grape berries (Fig. 1). We were able to purify milligrams of this protein by anion exchange chromatography and use them for determination of protein identity as well as immunization of the mice and development of different immunoassays.
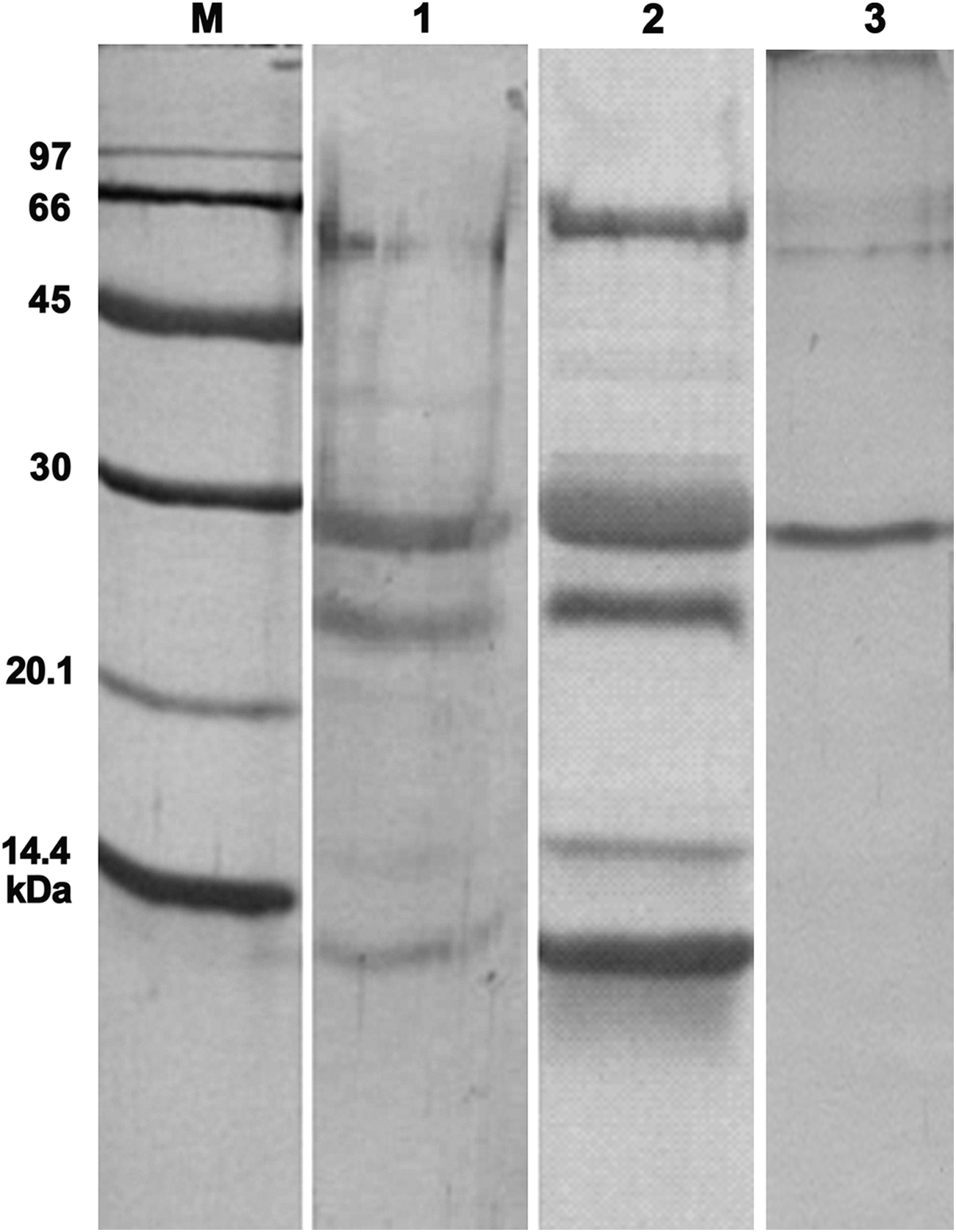
SDS-PAGE analysis of grape extract and the purified chitinase. M, Amersham low molecular weight marker; lane 1, fresh Sultana grape juice; lane 2, grape crude extract; lane 3, purified class IV chitinase. Staining method, Brilliant Coomassie blue.
Purity assessment of the eluted fractions showed that the highly expressed 28 kDa protein had been reasonably purified (Fig. 1). Mass spectrometry was used for characterization of protein identity. Peptide mass fingerprinting (PMF) and tandem mass spectrometry (MS/MS) results showed that the purified protein contained precursors matching highly with a 264 amino acid protein of Vitis vinifera. The observed mass of at least two peptides matched with an ion confidence interval of 100% and an ion score of 189 with class IV chitinase (Table 1). The results of PMF are shown in Figure 2.
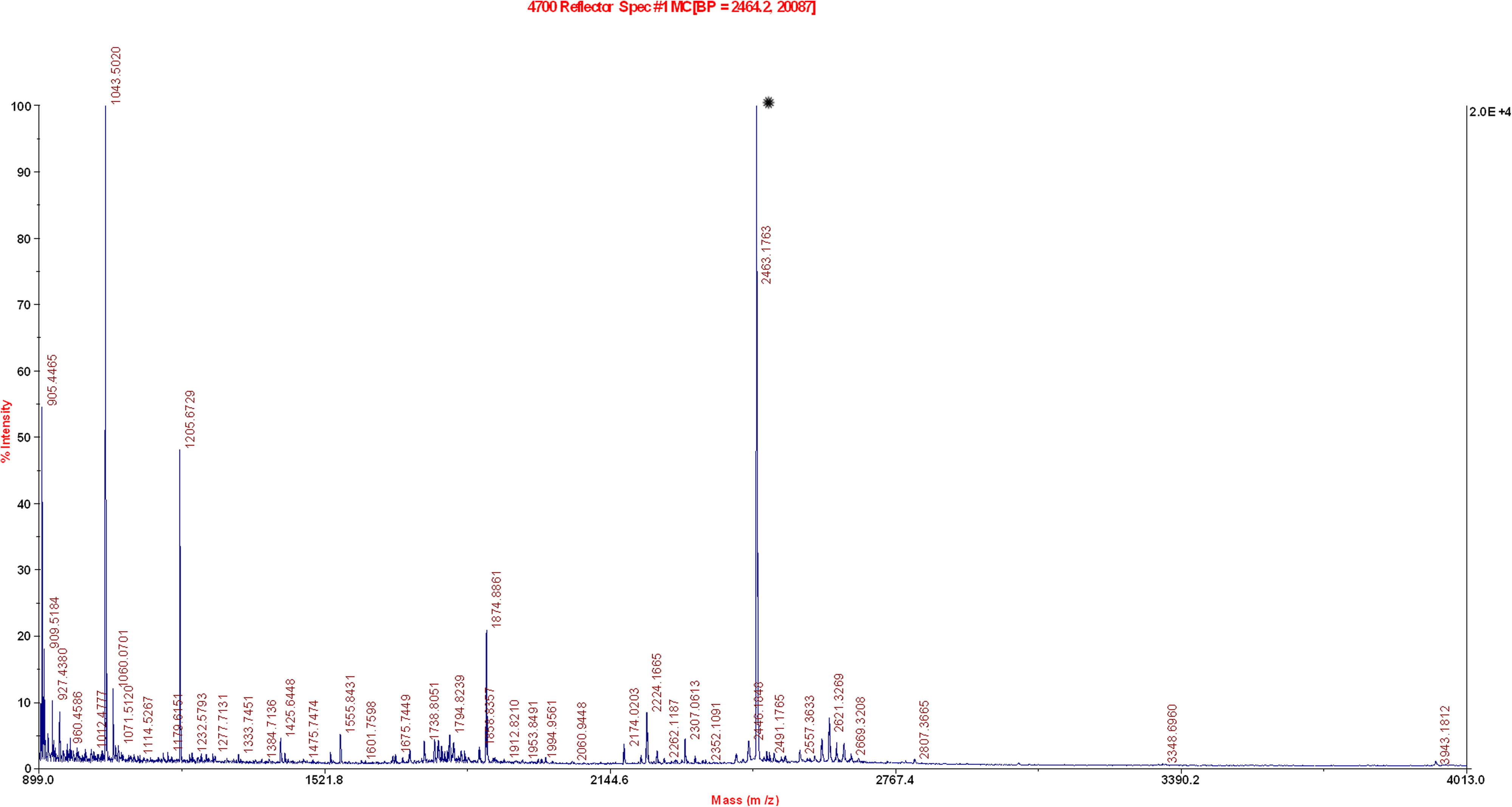
Peptide mass fingerprint result of purified protein. *The high intensity peptide corresponds to class IV chitinase.
Indirect ELISA showed that the titer of anti-chitinase antibody in mice sera was over 1:128,000. After fusion, screening of the plates showed that most of the wells contained at least one or two hybridoma cells. This finding proved that the mice were successfully immunized and the fusion protocol had an acceptable efficiency. Among 33 highly reactive hybridoma cells, only 10 were selected after the primary limiting dilution. Finally, according to the level of the optical density observed in ELISA, six clones were chosen after the second limiting dilution. Several clones lost their secretory state. After a third limiting dilution, results showed that the clones were stable and produced monoclonal antibodies with the same characteristics of the second limiting dilution clones. The results of limiting dilution and preparation of MAbs are shown in Table 2.
SP2/0 supernatant and 1:2000 diluted immunized mouse sera were used as negative and positive controls, respectively.
The rapid solid phase isotyping test revealed that IgG2a, IgG2b, and IgM were produced by one, two, and three clones. All of them contained kappa light chains. The characteristics of purified MAbs are summarized in Table 3.
Western blot analysis with crude extract showed that the monoclonal antibodies were specifically reactive with grape 28 kDa class IV chitinase (Fig. 3).

Western blot analysis of grape crude extract by supernatants of selected MAbs after final limiting dilution. M, Amersham low molecular weight marker; lanes 1 – 6, clones D1, D4, K1, L1, X1, Z1, respectively; N, supernatant of SP2/0 cell line as control.
Discussion
Grapes are one of the main agricultural products of Iran, and this fruit is reported as a frequent cause of fruit allergy in the country.(33) The potential of allergenicity of grapes and its by-products has been demonstrated in different studies.(15,16,28–30) To date, many severe anaphylactic reactions have also been reported in grape allergic patients.(15,29–32) Hundreds of grape cultivars are daily consumed as fresh fruits or raisins and are also used for production of wines all over the world. Yet differences in allergenicity among grape cultivars have not been studied.
It has been shown that fruit allergy is mostly caused by pathogenesis-related proteins such as lipid transfer proteins, thaumatin-like proteins, and chitinases with the approximate molecular weights of 9, 24, and 28 kDa, respectively.(34–36) Several studies have shown that these allergens compose the most abundant proteins in ripening grape berries. All of these proteins have a cysteine-rich amino acid sequence with several intra-chain disulfide bonds and possess a strict structure, which helps their successful extraction and concentration. The stability of these proteins is a concern for the wine industry too. They may encounter in turbidity of wines.(57) Therefore, assessment of the concentration of these proteins in different fruits and their by-products is of medical and industrial importance.
As Figure 1 shows, proteins with 9, 24, and 28 kDa molecular weights exist in the prepared grape total extract, as well as in fresh grape juice. The percentage of 28 kDa protein was higher than the others. These proteins were purified and analyzed by mass spectrometry. The MALDI TOF/TOF results showed that the highly expressed 28 kDa protein was a class IV chitinase, consistent with the finding of Robinson and colleagues.(21)
In addition to grapes, the allergenicity of chitinase has been confirmed in other allergenic fruits including banana (Mus a 2),(37) avocado (Pers a 1),(38) Chinese date (Ziz m 1),(39) chestnuts (Cas s 5),(40) and also aeroallergens from house dust mite (Der f 15, Der f 18)(41,42) and latex (Hev b 11).(43,44)
Standardization of SPT reagents, which are commonly used for allergy diagnosis, is a matter of interest in clinical immunology. A few studies showed that some commercial fruit extracts had poor quality and low allergic protein content. This could be due to a different ripening stage or unsuitable storage condition of the allergenic fruits before the total extract preparation.(45,46) Moreover, a variety of extracts are rich in proteolytic enzymes. These enzymes are capable of degrading crude extract allergens, resulting in unsuitable reagents.(47) Application of monoclonal antibodies against immunogenic epitopes of these allergens could be used to check if such an alteration has occurred on the major allergens or not. In several studies, MAb-based immunoassays were applied for standardization of diagnostic and therapeutic extracts, enlightening characteristics of a reliable reagent.(48–51) Moreover, determination of allergenicity potential of different cultivars needs development of appropriate immunoassays.(52) In this study, several monoclonal antibodies were produced against grape class IV chitinase, which specifically bind to their target and could be used for the allergen immunodetection.
Developing an easy method for extraction of grape class IV chitinase allowed us to extract enough protein for immunization of mice and development of indirect ELISA. The analyzing of crude extract and fresh juice of grape by Image J software showed that class IV chitinase is one of the abundant proteins of ripened grapes. This software also showed that this allergic protein represents about 40.3% and 51.5% of fresh grape juice and crude extract, respectively. The molecular structure of chitinase is stabilized by several disulfide bonds, which protect this molecule against possible proteolytic degradation during storage or protein extraction procedures. Moreover, because of the low PI, it was easily purified from other grape proteins by anion exchange chromatography.
Following fusion, the primary results indicated that various reactive clones had been generated. Wells with higher levels of optical density in ELISA were chosen and underwent three limiting dilutions. Finally, gaining similar optical densities in the second and third limiting dilutions confirmed that the clones were stable and further limiting dilution was not necessary (Table 3).
The rapid isotyping test showed that half of the clones produced IgG and the rest produced IgM. Since, the obtained IgM anti-chitinase antibodies showed lower signals in ELISA, the affinity of IgM antibodies were not determined. Meanwhile, the affinity of IgG monoclonal antibodies was in the range of 108 – 109 M−1, and they could be applied for quantitative and qualitative tests. Clone D1 had the highest affinity, which was 1.2 × 109 M−1; the affinities of K1 and Z1 were 7 × 108 and 3 × 108 M−1, respectively, indicating that these MAbs could be used for ELISA development.
Western blot analysis of the MAbs with crude extract demonstrated that they did not have significant cross-reaction with other grape proteins, and they had acceptable specificity to grape class IV chitinase. Dot blot inhibition assay revealed that the purified chitinase was capable of reducing the reactivity of a mixture of MAbs with the grape crude extract up to 80% (as calculated by Image J software), indicating that purification procedure did not affect protein integrity and that both purified and unpurified chitinase had a similar binding characteristic in immunoblotting assay. All of the produced antibodies were reactive in ELISA and Western blotting, showing that they could recognize both native and denatured forms of the enzyme. It is likely that the produced IgGs could be applied in immunohistochemical assays to demonstrate the enzyme location in grape berry and other tissues of this plant too, but this application has not been checked yet.
In conclusion, a few monoclonal anti-chitinase antibodies have been previously produced against bacterial or insect chitinases,(41,53–55) but this is the first report of production of monoclonal antibodies against fruit chitinases.
Footnotes
Acknowledgment
This study was supported by Grant 88449 from the Research Administration Department of Mashhad University of Medical Sciences, Mashhad, Iran. This article was derived from the Ph.D. thesis of the second author (Thesis No. A-277).
Author Disclosure Statement
The authors have no financial conflicts to disclose.