
Abstract
CCR5 (CD195) is a receptor for the chemokines RANTES, MIP-1α, and MIP-1β and is used by HIV-1 as a co-receptor for entry into macrophages and CD4+ T cells. CCR5 exists in multiple conformations in the membrane and is present at low levels on human macrophages, making it difficult to detect. Nine commercially available anti-CCR5 monoclonal antibodies were evaluated for their specificity and their recognition of CCR5 expressed by macrophages. Unexpectedly, we found that three of the nine clones tested displayed substantial background binding to CCR5 negative cells, suggesting that these antibodies may give unreliable results. We recommend the use of clones CTC8, 45531 (PE conjugated), and 45523 and advise that data obtained using CTC5, 45531 (unconjugated), and 45549 anti-CCR5 antibody clones should be interpreted with care.
Introduction
CCR5 has seven hydrophobic transmembrane domains, three extracellular loops, and an extracellular N-terminal domain (Fig. 1). There are four tyrosine residues in the N terminus, at least some of which are sulfated post-translationally. This is important for both ligand binding and for HIV-1 entry as they supplement the negative charge in the region; it has been shown that sulfotyrosines at positions 10 and 14 are the most crucial for these functions. CCR5 has four cytoplasmic loops, with the fourth being created by the association of three palmitoylated cysteine residues, located towards the C-terminus, with the membrane (this may also target the protein to lipid rafts). A conserved sequence in the second intracellular loop is thought to interact with a G protein, and the N-terminal domain is important for interactions with chemokines and with HIV-1 gp120.(1) The C-terminal domain is critical for interactions with kinases that regulate receptor function and with arrestin proteins that mediate receptor internalization.(1,8)
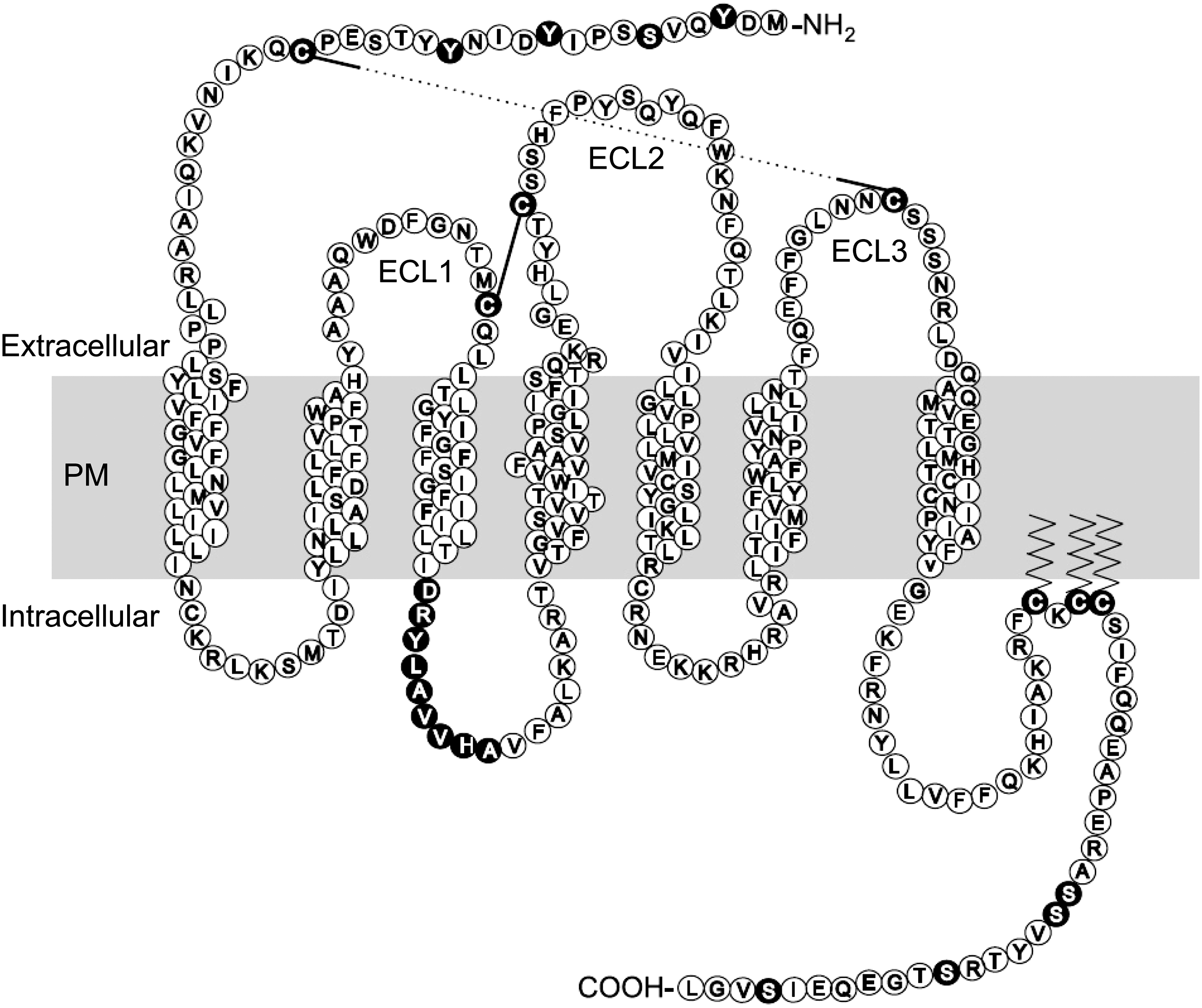
Two-dimensional representation of CCR5 sequence and topology in the plasma membrane. Amino acids critical for CCR5 function are denoted by filled circles, and cysteine bonds are shown by black lines. Palmitoylation is indicated by zig-zag lines. PM, plasma membrane (approximate position); ECL, extracellular loop. Representation reproduced (additional labels added) with kind permission of Prof. M. Oppermann.
Chemokine binding to CCR5 leads to G protein activation, and subsequent synthesis of intracellular signaling molecules such as cyclic AMP (through adenylyl cyclase) and release of intracellular calcium (via phospholipase C activation). The latter leads to activation of protein kinase C (PKC), which can phosphorylate CCR5 and terminate the signaling through subsequent receptor internalization and degradation. CCR5 ligands are also able to activate MAP kinase signaling, and RANTES binding has been shown to activate phosphatidylinositol-3 kinase (PI-3) kinase and RhoA in T cells.(1,8) Along with PKC, serine/threonine protein kinases called G protein coupled receptor kinases phosphorylate CCR5 to mediate receptor desensitization following signaling.(1,8)
CCR5 is recycled continuously from the plasma membrane, through early endosomes, and back to the plasma membrane. Arrestins act as adaptor proteins that link CCR5 to clathrin and AP-2 adaptors, which effect endocytosis. Caveolar internalization pathways may also be in operation for this receptor,(2) although this has been disputed.(9)
CCR5 is thought to exist in multiple conformations in the membrane, and this influences recognition of the protein by antibodies.(10,11) It is important to be able to accurately measure levels of CCR5 on the cell surface when investigating HIV-1 infection of macrophages, as receptor concentration is key to susceptibility to infection.(12) We therefore investigated the specificity of a panel of commonly used anti-CCR5 monoclonal antibodies (MAbs), as well as determined their ability to recognize CCR5 on the surface of primary human macrophages. Our findings suggest that several of the anti-CCR5 MAbs that are commercially available lack specificity; thus results using these reagents should be interpreted cautiously.
Materials and Methods
Cell culture
CD14+ cells were isolated from peripheral blood mononuclear cells (PBMCs) using anti-human CD14 magnetic beads (Miltenyi Biotec, Bergisch Gladbach, Germany) according to the manufacturer's instructions, except that 1 μL of beads per 1.5×106 PBMCs was used. Cells were plated onto tissue culture-treated 6-well plates (Corning, NY) at a density of 1.5×106 cells/well, and differentiated in RPMI 1640 (PAA) with 10% FCS, 2 mM L-glutamine (PAA), 100 U/mL penicillin, 100 μg/mL streptomycin (PAA), and either 100 ng/mL recombinant human M-CSF (∼1.7×104 units of biological activity per mL) or 50 ng/mL GM-CSF (∼415 U/mL), both from R&D Systems (Minneapolis, MN). Cells were incubated at 37°C, with 5% CO2. Monocytes were differentiated for 7 to 9 days prior to use.
293T cells were cultured in DMEM containing 10% FCS, 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin. GHOST parental cells and GHOST Hi-5 cells were grown in DMEM containing FCS, L-glutamine, penicillin, and streptomycin as above, supplemented with 500 μg/mL G418 and 100 μg/mL hygromycin (both Sigma, St. Louis, MO). In the case of GHOST Hi-5 cells, 1 μg/mL puromycin (Sigma) was also added to retain CCR5 expression. 293T and GHOST cells were incubated at 37°C, with 10% CO2, and were passaged 24 h prior to flow cytometry.
Flow cytometry
Macrophages were lifted by pipetting with PBS containing 5 mM EDTA (Sigma) and 12 mM lidocaine (Sigma) after 1 h on ice. GHOST and 293T cells were detached with 5 mM EDTA. Cells were centrifuged at 514 g for 5 min at 4°C, resuspended in flow cytometry buffer (FCB; PBS, 0.01% NaN3, 1% FCS, 10 μg/mL human IgG) and left at 4°C for 10 min in order for blocking to occur. 1×105 cells per antibody were stained in a total volume of 50 μL of antibody diluted in FCB, for 30 min at 4°C (with mixing after 15 min) in the dark. For fluorophore-conjugated primary antibodies, the cells were washed twice in FCB and fixed with 4% formaldehyde in PBS. In the case of unconjugated primary antibodies, the cells were washed twice after staining, and then incubated for a further 20 min with a PE-conjugated polyclonal secondary antibody at 4°C in the dark. Cells were washed a further two times in FCB, and fixed as above (see Table 1 for details of antibodies used). Flow cytometry was carried out using a FACSCalibur flow cytometer (Becton-Dickinson, Franklin Lakes, NJ), and data were analyzed using FlowJo software (v. 7.1.3, Ashland, OR).
Treatment with RANTES
In one experiment, M-CSF-derived macrophages were pre-treated with RANTES prior to flow cytometry. Macrophages were lifted as above, and following centrifugation at 514 g for 5 min at 4°C, cells were resuspended in 100 nM RANTES (PeproTech, Rocky Hill, NJ) in RPMI at a concentration of 1×107 cells/mL and incubated at 4°C or 37°C for 30 min. Cells were washed three times with PBS and prepared for flow cytometry analysis as described above.
Results
Some anti-CCR5 MAb clones are not useful for detecting CCR5 on macrophages
Two anti-CCR5 MAb clones supplied by Becton Dickinson, 2D7 and 3A9, are widely used for detection of CCR5 expression. To determine whether these antibodies could be valuable for use with primary macrophages, we stained two types of human macrophage, one differentiated from blood monocytes with M-CSF (M-MΦ) and another derived with GM-CSF (GM-MΦ), with unconjugated 2D7 and 3A9, followed by a fluorescent secondary antibody and detection of binding by flow cytometry. For GM-MΦ, very small amounts of staining were seen above isotype control levels (Fig. 2A). However for M-MΦ, virtually no staining was seen at all (Fig. 2A), suggesting that these clones do not efficiently recognize CCR5 on the surface of primary human macrophages. This confirms the observations of others in our group (Dr. Karl Karlsson, unpublished data). These antibodies also did not bind to CCR5 negative 293T cells as expected (Fig. 2A).
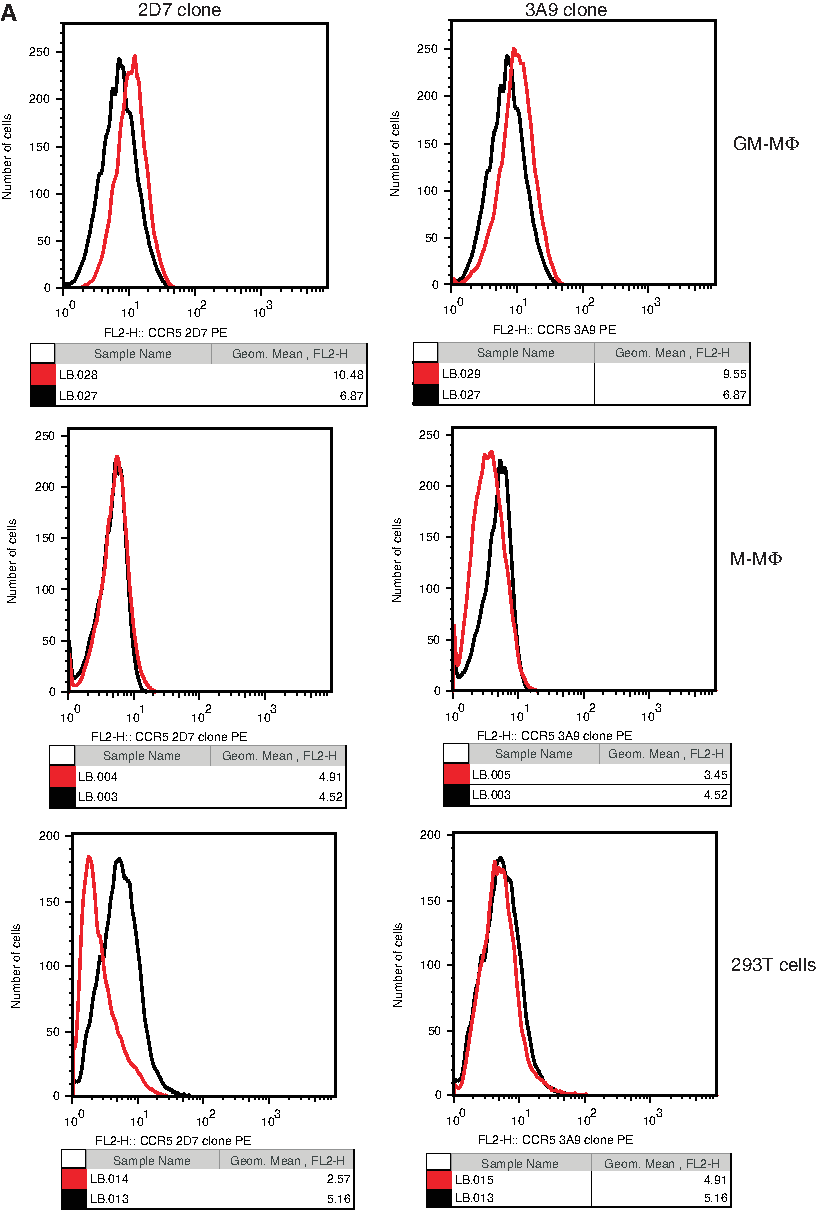
Anti-CCR5 MAb clones 2D7, 3A9, and R22/7 do not bind efficiently to CCR5 on MΦ or to CCR5-negative 293T cells. GM-MΦ, M-MΦ, and 293T were incubated with anti-CCR5 MAb clones (
We then tested an antibody supplied by Santa Cruz Biotechnology (clone R22/7, PE conjugated; Santa Cruz, CA). Compared to an isotype control, very low levels of staining were seen with M-MΦ and GM-MΦ from two donors (Fig. 2B) using 1:40 dilution. Doubling the concentration did not appreciably increase the level of staining seen (data not shown). No staining was seen with 293T cells as expected (Fig. 2B).
Anti-CCR5 MAb clones from R&D Systems bind to M-MΦ and GM-MΦ
R&D Systems supplies six clones of anti-CCR5 MAbs. Detailed epitope mapping of these clones has been performed in 293T cells by Lee and colleagues(11); two were reported to recognize linear epitopes in the N-terminus (clones CTC5 and CTC8), one recognized a non-linear N-terminal epitope (45502), clone 45531 bound to extracellular loop 2B (ECL-2B), and the remaining two clones (45523 and 45549) recognized epitopes comprising multiple domains.(11)
We tested unconjugated versions of CTC5 and CTC8 with M-MΦ (Fig. 3A) and GM-MΦ (Fig. 3C), and found that both of these clones stain both cell types. This staining revealed two populations of cells, a low-staining group representing the majority and a minor population of more highly stained cells. The pattern was identical when we used a FITC-conjugated version of CTC5 to stain M-MΦ and GM-MΦ (Fig. 3B, D). We next tested the four remaining anti-CCR5 clones from R&D Systems: 45502, 45523, 45531, and 45549 (all unconjugated MAbs). All four resulted in a higher level of fluorescence than the isotype control with both M-MΦ and GM-MΦ (Fig. 3A, C). Clones 45531 and 45549 produced the highest levels of staining, and two populations with differential levels of binding were revealed by 45531 staining of M-MΦ and 45549 staining of GM-MΦ (Fig. 3A, C). In contrast, staining with directly conjugated MAb clone 45531 showed a single population for both M-MΦ and GM-MΦ, and suggests that CCR5 expression is normally distributed (Fig. 3B, D).

Anti-CCR5 MAb clones from R&D Systems bind to M-MΦ and GM-MΦ. (
To demonstrate that PE-conjugated clone 45531 staining is specific for CCR5, we pre-treated M-MΦ with the CCR5-binding chemokine RANTES. RANTES induces the internalization of a proportion of CCR5 from the surface of macrophages(13) and treatment at 37°C should result in reduced anti-CCR5 antibody binding. After a 30 min incubation at 37°C with 100 nM RANTES, a 28% reduction in mean fluorescence intensity was seen with PE-conjugated clone 45531 compared to untreated cells (Fig. 3E). When the experiment was repeated at 4°C, an 11% reduction was seen (Fig. 3E). This indicates that at least a proportion of PE-conjugated clone 45531 binding is specific for CCR5.
Anti-CCR5 MAb clones from R&D Systems stain CCR5 negative 293T cells
Untransfected 293T cells should not express CCR5, therefore they can be used to confirm the specificity of anti-CCR5 MAbs for CCR5 positive cells. Surprisingly, we found that CTC5 and CTC8 stained 293T cells substantially above the levels of an appropriate isotype control (Fig. 4A). In the case of CTC5, the geometric mean of the histogram plot was almost 9-fold above that of the isotype control (and the histogram showed an additional high-staining shoulder), and for CTC8 it was 6.5-fold higher. Unconjugated 45531 stained 293T cells very highly, as did clone 45549 (Fig. 4A).

Anti-CCR5 MAb clones from R&D Systems stain CCR5-negative 293T cells. (
To determine whether the high background seen with CTC5 was due to a manufacturing issue, we tested CTC5 from another manufacturer, Leinco Technologies (St. Louis, MO). This antibody also stained 293T cells above the level of the isotype control (Fig. 4B); however the geometric mean was lower and the population was normally distributed. Finally, we tested fluorophore-conjugated clone 45531 with 293T cells. This resulted in low levels of staining, which were only slightly above isotype control background levels (Fig. 4B). Strikingly, this PE-conjugated antibody behaves very differently from the unconjugated version of the same clone, despite both being obtained from the same supplier.
Detailed analysis of specificity of anti-CCR5 MAbs
An ideal way to confirm antibody specificity is to use cell lines that are identical in every way except in the expression of the target protein, as this removes the possibility of differential non-specific binding to other components of the cells. In order to do this, we used two cell lines called GHOST and GHOST Hi-5. GHOST cells were derived from a human osteosarcoma cell line and they stably express CD4. From these, GHOST Hi-5 cells were derived, with stable expression of CCR5.(14) Antibody staining was performed on both cell lines in parallel, analyzed using the same flow cytometer settings, and the results compared.
Clone CTC5
Unconjugated CTC5 gave a small amount of background staining on parental GHOST cells (Fig. 5A), but the peak moved substantially to the right for CCR5 positive cells (Fig. 5C). Overlaying the profiles of the parental and CCR5 positive cells shows a good distinction between the two cell types (Fig. 5E).

Detailed analysis of the specificity of anti-CCR5 MAbs using GHOST and GHOST Hi-5 cell lines. (
Clone CTC8
Staining with unconjugated CTC8 resulted in low levels of background on parental GHOST cells (Fig. 5A), and high levels of staining for GHOST Hi-5 cells (Fig. 5C). The distinction between CCR5 negative and positive cells was very good (Fig. 5E), and the populations appeared to be normally distributed. CTC5 and CTC8 both recognize a linear epitope and can therefore be used for Western blotting of CCR5. Based both on these results and on the 293T data (Fig. 4A), CTC8 is more specific and should be used preferentially to CTC5 for Western blotting.
Clone 45531
Staining with unconjugated 45531 revealed a large amount of background binding to parental GHOST cells (Fig. 5A) compared to the isotype control staining. In contrast, background was minimal with the PE-conjugated version of 45531 (Fig. 5A). The geometric mean fluorescence more than doubled for unconjugated 45531 when it was used to stain GHOST Hi-5 cells, and staining also increased substantially (6.5-fold increase) for the conjugated version (Fig. 5C). Figure 5E shows the overlay of CCR5 negative and positive results for these antibodies.
Clone 45549
Unconjugated 45549 bound to CCR5 negative GHOST cells (Fig. 5A) more than the isotype control. The geometric mean value increased with CCR5 positive GHOST cells (Fig. 5C). However, an overlay of the histograms shows poor differentiation between negative and positive cells, as the negative peak completely encompasses the positive peak (Fig. 5E).
Clone 45502
This clone is reported to recognize CCR5 on transfected cell lines, but not CCR5 on PBMCs unless the cells are first fixed. We therefore tested 45502 on both formaldehyde fixed and unfixed GHOST cells. No background was seen on unfixed parental GHOST cells and the majority of fixed cells, but a second peak representing a slightly higher staining population was also observed with fixed cells (Fig. 5B). In the case of CCR5 positive GHOST cells, no staining was seen for unfixed cells, showing that, contrary to the information provided by R&D Systems, fixation is necessary with CCR5 transfected cell lines (although surprisingly, clone 45502 produced low level staining of unfixed macrophages; Fig. 3A, C). When the cells were fixed prior to staining, a small amount of staining was seen, with a bimodal profile (Fig. 5D). Figure 5F shows the overlay of the histograms for parental and Hi-5 cells.
Clone 45523
As with the previous clone, 45523 is thought to recognize CCR5 on transfected cell lines, but not on unfixed PBMCs. No staining was seen with CCR5 negative GHOST cells (either fixed or unfixed, except for a small shoulder with fixed cells; Fig. 5B). Staining increased dramatically for CCR5 positive GHOST cells, both fixed and unfixed (Fig. 5D). Overlays of the parental and Hi-5 histograms show excellent differentiation (Fig. 5F). As the data were normally distributed for unfixed cells, we would recommend not fixing cells prior to staining with this antibody, particularly as 45523 was able to stain unfixed macrophages (Fig. 3A, C).
Discussion
The data presented here show that several anti-CCR5 MAbs stain cells that do not express CCR5, often to high levels. This issue was not resolved despite extensive trouble-shooting discussions with R&D Systems. Table 2 summarizes the results of Figures 1–5. Based on these findings, we would recommend using CTC8 for Western blot analysis (or other uses requiring a linear epitope), and PE-conjugated clone 45531 or unconjugated clone 45523 (both with unfixed cells) for general flow cytometry staining of CCR5.
Anti-CCR5 MAb clones 2D7, 3A9, and R22/7 were found not to stain CCR5 on the surface of M-MΦ and GM-MΦ efficiently. Macrophages are known to express CCR5 and to support entry of R5 tropic HIV-1; blockade of CCR5 with a small molecule inhibitor (Tak-779) is able to inhibit HIV-1 infection of macrophages.(15) Therefore these MAbs should bind to MΦ above the level of an isotype control. A possible explanation for the negative results seen is that the conformation of CCR5 on macrophages is not optimal for recognition by these antibodies, as CCR5 does exist in multiple conformations.(10,11) Epitope mapping has been performed for clone 3A9 and supports conformation-dependent binding; mutation of sequences in the N-terminus, ECL1, and ECL3 inhibited binding(16) although the N-terminal mutations had the largest effect. 2D7 binds exclusively to ECL2 and also depends on a precise conformation of the protein.(17,18) However, 2D7 has also been described as recognizing a large proportion of the CCR5 on human cells,(11) arguing against this explanation for low binding to macrophages. R22/7 was raised against a synthetic peptide near the N-terminus of CCR5, and it recognizes a linear epitope as it is recommended for use in Western blotting. Therefore it is difficult to explain why such low levels of staining were observed with this clone.
CCR5 MAb clones from R&D Systems were all able to bind to M-MΦ and GM-MΦ, at varying levels, and none of the antibodies tested required prior fixation of the cells for binding to occur, despite the fact that datasheets provided by R&D Systems for clones 45502 and 45523 specify that fixation of PBMCs is necessary for CCR5 recognition. RANTES treatment of M-MΦ caused a reduction in staining by PE-conjugated clone 45531, providing evidence for at least some specific staining by this antibody. It is not possible to determine the proportion of staining that is specific because RANTES may not result in complete CCR5 internalization.(13)
In order to test the specificity of the CCR5 MAbs further, we measured the binding of four clones to 293T cells, which are CCR5 negative. Care was taken to use identical concentrations of antibody for isotype controls (which were also from R&D Systems) and anti-CCR5 MAbs, and to run samples in parallel. All of the clones showed significant binding except 45531 (fluorophore conjugated). More extensive testing with GHOST and GHOST Hi-5 cell lines confirmed that high background binding occurred with CCR5 negative cells, except for clones CTC8, 45502, 45523, and fluorophore-conjugated clone 45531. Surprisingly, the unconjugated form of the 45531 MAb bound to 293T cells and GHOST cells at a much higher level. It is difficult to conceive of reasons why identical Abs that differ only in fluorophore conjugation should show such different binding patterns; the presence of the fluorophore does not usually impact antigen binding, and it is likely that this would have been tested by the manufacturer during initial product development. It may be that the antibody is not efficiently labeled; therefore binding could be under-represented by the fluorescent signal being detected.
The high background binding of many of these MAb clones to cells that are negative for the target antigen suggests a lack of specificity. However 293T cells (CCR5 negative and CCR5 transfected) were used for the extensive epitope mapping of these MAbs conducted by Lee and colleagues,(11) the group that initially generated the MAbs, and it would have been largely impossible to perform such fine mapping if high background binding was present. This implies that manufacturing errors have led to reductions in specificity, now that the antibodies are available commercially. Despite this, it is clear that all of the MAb clones from R&D Systems do recognize CCR5, since binding increased substantially with GHOST Hi-5 cells when compared to CCR5 negative GHOST cells. Care should be taken when selecting CCR5 MAbs for use in experiments and in interpreting the results; the clones with better specificity should be used preferentially. It may also be necessary to revisit published studies that relied on these antibodies for key conclusions.
Footnotes
Acknowledgment
This work was supported by a Ph.D. studentship from the UK Medical Research Council.
Author Disclosure Statement
The authors have no financial interests to disclose.