
Abstract
ST2L is a transmembrane receptor that belongs to the IL-1 receptor family. The receptor is expressed on various cell types including Th2 cells, mast cells, basophils, growth-activated fibroblasts, and vascular endothelial cells. ST2L activation by its ligand IL-33 has been implicated in Th2-mediated immunity, inflammation, and allergic responses in vivo. Inhibition of ST2L activity can attenuate Th2-dominated immune responses such as lung eosinophilia, airway hyper-responsiveness, and arthritis in animal models. Here we report the generation and in vitro characterization of a panel of rat anti-mouse ST2L monoclonal antibodies. We demonstrate that the antibodies specifically bind to recombinant receptor protein and that a subset of the binders inhibits mouse ST2L activity in multiple in vitro assays. Four of the identified anti-mouse ST2L antibodies were shown to prevent IL-33 from binding to ST2L, down-regulate IL-33-induced NF-κB signaling, and neutralize the ability of IL-33 to stimulate mouse Th2 cell proliferation. The characterized monoclonal antibodies are important tools that will be used to study mouse ST2L receptor functionality in vivo.
Introduction
IL-33 has been identified as a ligand for ST2L.(8) Upon IL-33 binding, cell surface ST2L forms a signaling complex with IL-1R accessory protein (IL-1RAcP) that activates intracellular pathways, including MAP kinase and NF-kB signaling, and induces the synthesis of Th2-type cytokines.(16) sST2, which also binds to IL-33, has been reported to be a negative regulator of ST2L signaling in vitro and in vivo, possibly as a decoy receptor for IL-33.(11,17)
A significant amount of evidence supports a role for ST2L in inflammation and autoimmune diseases. ST2L is selectively expressed on Th2 but not Th1 lymphocytes.(2) TH2 cells are required for the development of allergic inflammation, and ST2L activation on Th2 cells has been shown to contribute to the maintenance of an allergic response in antigen-induced models of asthma.(12,18) Blocking IL-33/ST2 signaling using an antibody against ST2 down-regulates persistent airway hyper-responsiveness.(12) ST2 knock-out mice develop attenuated collagen-induced arthritis (CIA), and IL-33 has been shown to exacerbate CIA in ST2 knock-out mice engrafted with mast cells from ST2 wild-type mice but not from ST2 knock-out mice.(19)
Systemic IL-33 administration in ST2 wild-type but not knock-out mice induces the secretion of TH2-type cytokines IL-5 and IL-13, eosinophilia, increased serum IgE and IgA levels, lung epithelial cell hyperplasia, and increased mucus production.(8) Thus, modulators of ST2L signaling, such as neutralizing anti-ST2L antibodies, could be used as a tool for investigating the role of this receptor in inflammatory disease models. We have generated a panel of rat anti-mouse ST2L monoclonal antibodies and characterized these antibodies for the ability to bind and neutralize ST2L activity in vitro. Based on in vitro antibody characterization assays, a potent neutralizing anti-mouse ST2L antibody was selected for investigation of the role of ST2L signaling in vivo.
Materials and Methods
Recombinant proteins
Mouse recombinant IL-33 (aa121–270), mouse recombinant ST2-Fc chimera, anti-mouse ST2 MAb MAB10041, and rat IgG isotype controls were purchased from R&D Systems (Minneapolis, MN).
Biotin conjugations
mIL-33 and mST2-Fc were conjugated to biotin using EZ-Link NHS-Biotin kit (Thermo Scientific, Rockford, IL) following the manufacturer's instructions. Bioactivity of biotinylated mIL-33 was confirmed in an ST2 binding ELISA assay. Bioactivity of biotinylated mST2/Fc was confirmed by its ability to neutralize IL-33 activity in an NF-κB reporter gene assay.
Cell culture
The mouse TH2 cell line D10.G4.1 (ATCC, Manassas, VA) was cultured in RPMI 1640 supplemented with 10% FBS, 2 mM L-glutamine, 0.5 g/L sodium bicarbonate, 4.5 g/L glucose, 1.0 mM sodium pyruvate, 0.05 mM 2-mercaptoethanol (Invitrogen, Carlsbad, CA), 10 pg/mL IL-1alpha (R&D Systems), and 10% rat T-STIM factor with Con A (Becton Dickinson, Franklin Lakes, NJ).
The HEK293 cell line was cultured in DMEM with 10% FBS (Invitrogen).
Monoclonal antibody production
Rats were immunized intraperitoneally with mouse ST2-Fc (R&D Systems) and assessed for specific IgG titers. Once sufficient titers were obtained, splenocytes were isolated and fused with FO cells. The resulting hybridomas were plated in 96-well plates or methylcellulose and cultured for 10 days. Antigen-specific clones were identified by standard capture ELISA for binding to mST2-Fc and cross-screened against the Fc protein alone. Murine ST2-specific hybridomas were further tested for the inhibition of IL-33 binding to ST2 in an ELISA and for the inhibition of IL-33-induced D10.G4.1 mouse TH2 cell proliferation. Hybridomas exhibiting neutralization in both receptor-ligand binding and cell-based proliferation assays were cloned by limiting dilution.
ST2L binding ELISA
Hybridoma supernatants containing rat anti-mST2 antibodies were added to 96-well plates (VWR, West Chester, PA) coated with 1 μg/mL of anti-rat Fc-specific antibodies (Jackson Immunoresearch, West Grove, PA) followed by the addition of 0.2 μg/mL biotinylated mST2-Fc chimeric protein. Biotinylated protein binding was detected with streptavidin-HRP (Jackson Immunoresearch) and TMB substrate (RDI Division of Fitzgerald Industries, Concord, MA), following the manufacturer's instructions. The following ELISA assay format was used for testing purified antibodies. Murine ST2-Fc was coated on the plate at 2 μg/mL. Wells were incubated with blocking buffer and washed and various amounts of purified anti-mouse ST2 antibodies were added. The detection was done with anti-rat IgG conjugated to HRP (Jackson Immunoresearch) and development with TMB substrate (RDI Division, Fitzgerald Industries) following the manufacturer's instructions.
Receptor-ligand inhibition ELISA
A 96-well clear plate (VWR) was coated with 50 μL of 2 μg/mL goat anti-human IgG, Fcγ fragment-specific (Jackson Immunoresearch) antibody for approximately 16 h at 4°C. The remaining steps were completed at room temperature. Wells were incubated with blocking buffer and washed, and 50 μL of 2 μg/mL mouse ST2-Fc were added for 1 h. The plate was washed and 1 μg/mL of biotinylated mIL-33 with or without anti-mST2 antibodies was added. The plate was washed and detection was done with streptavidin-HRP (Jackson Immunoresearch) and signal developed with TMB substrate (RDI Division, Fitzgerald Industries) following the manufacturer's instructions.
D10.G4.1 mouse TH2 cell proliferation assay
D10.G4.1 cells were washed twice with assay media (RPMI, 10% FBS, no IL-1, no T-STIM), resuspended at 1.25 × 105 cells per mL, and plated in 80 μL of medium in white clear-bottom TC-treated 96-well plates (NUNC, Rochester, NY). Various amounts of mouse IL-33 were added to the cells for the final assay volume of 100 μL. When testing antibody neutralization, control antibodies (spiked in spent hybridoma medium) or hybridoma supernatants were added to the cells and incubated for 1 h followed by addition of 20 pg/mL mIL-33. The plates were cultured for 24 h in humidified incubator at 37°C, 5% CO2. Quantitation of viable cells was achieved with CellTiter-Glo reagents (Promega, Madison, WI); protocol was performed according to the manufacturer's instructions.
DNA construct, transient transfections, and reporter gene assay
The murine ST2L expression vector was generated by isolating full-length murine ST2L using mouse polyA RNA from mouse spleen DNA (Ambion, Austin, TX) and the Superscript One Step kit (Invitrogen), and following gene-specific primers with restriction sites for cloning:
5′-TTT ATA AAG CTT GCC GCC ACC ATG ATT GAC AGA-3′ 5′-TAT AAA CTC GAG TCA AAA GTG TTT CAG GTC TAA-3′
The isolated DNA was then run through a 1% agarose TBE gel and extracted using the Qiagen gel-extraction kit (Valencia, CA). Extracted DNA was then cloned into a vector for sequencing using the pcDNA3.1V5HIS-TOPO kit from Invitrogen. TOP10 chemically competent Escherichia coli (Invitrogen) were then transformed with the cloned DNA and plated on Teknova agar AMP100 plates (Hollister, CA). Individual colonies were grown in Luria broth + 100 μg/mL ampicillin overnight. Plasmid DNA was extracted from the cultures using a Qiagen kit (#27106) and sequenced using gene specific primers (listed below), T7 forward and EBV reverse vector primers using the MJ research tetrad thermal cycler and ABI3100 sequencer. The output was aligned to NCBI accession #NM_001025602 murine ST2L variant #1. The clone was verified as being full-length murine ST2L.
Gene-specific sequencing primers:
5′-GTA CTC GAC AGT ACG TGG ATC AG-3′ 5′-CCT CAG TGT TAA GGA TAA CTG GTG-3′ 5′-GTT CAC CAC ACT CTG CCC GAC GTT C-3′
For the reporter gene assay HEK293 cells were plated at 50,000 cells per well in white clear-bottom tissue culture-treated 96-well plates (NUNC) in DMEM and 10% FBS, and incubated in humidified incubator at 37°C, 5% CO2 for 24 h. Transfection mixtures of mST2L vector, NF-κB-Luciferase vector (Stratagene, Agilent Technologies, Santa Clara, CA), and Lipofectamine 2000 in Opti-MEM media (Invitrogen) were added to the cells and incubated for an additional 24 h at 37°C, 5% CO2. Cells were then treated with mIL-33 with or without antibodies for 16 h at 37°C, 5% CO2, and luciferase activity was measured using Steady-Glo reagent (Promega), according to the manufacturer's instructions.
Antibody purification
Hybridoma culture supernatants were mixed with MabSuRe (GE Healthcare Bio-Sciences, Piscataway, NJ) Protein A resin, which was pre-equalibrated with 1x D-PBS. The protein bound resins were then washed with the same 1x D-PBS buffer to remove the nonbinding components. All samples were then eluted with 0.1 M sodium acetate (pH 3.5). The eluted proteins were immediately neutralized with the addition of a concentrated buffer of 2.5 M Tris (pH 7.2). The samples were subsequently dialyzed in 1x D-PBS buffer.
Results
Generation of anti-mouse ST2L monoclonal antibodies
The extracellular domain of mouse ST2 fused to human IgG Fc protein was used for immunizations in rats. A panel of hybridomas was generated by fusing splenocytes from immunized animals with the myeloma cell line F0. In order to identify anti-ST2 secreting hybridomas, supernatants were screened for the binding to mST2-Fc in ELISA format. All the binders were cross screened for binding to human IgG Fc protein by ELISA. Forty-eight ST2-specific supernatants were identified and tested for the inhibition of ST2/IL-33 binding.
Screening for anti-mouse ST2L neutralizing hybridomas
Forty-eight mouse ST2-specific hybridoma supernatants were tested for the ability to neutralize binding between mouse ST2-Fc coated on the plate and biotinylated mouse IL-33. Antibody concentrations in the supernatants were not determined prior to screening. Results of the binding inhibition analysis are presented in Figure 1 with the supernatants exhibiting various degrees of inhibition. Six of the strongest inhibitors (4, 24, 25, 29, 45, 46) showed nearly 100% inhibition of binding.
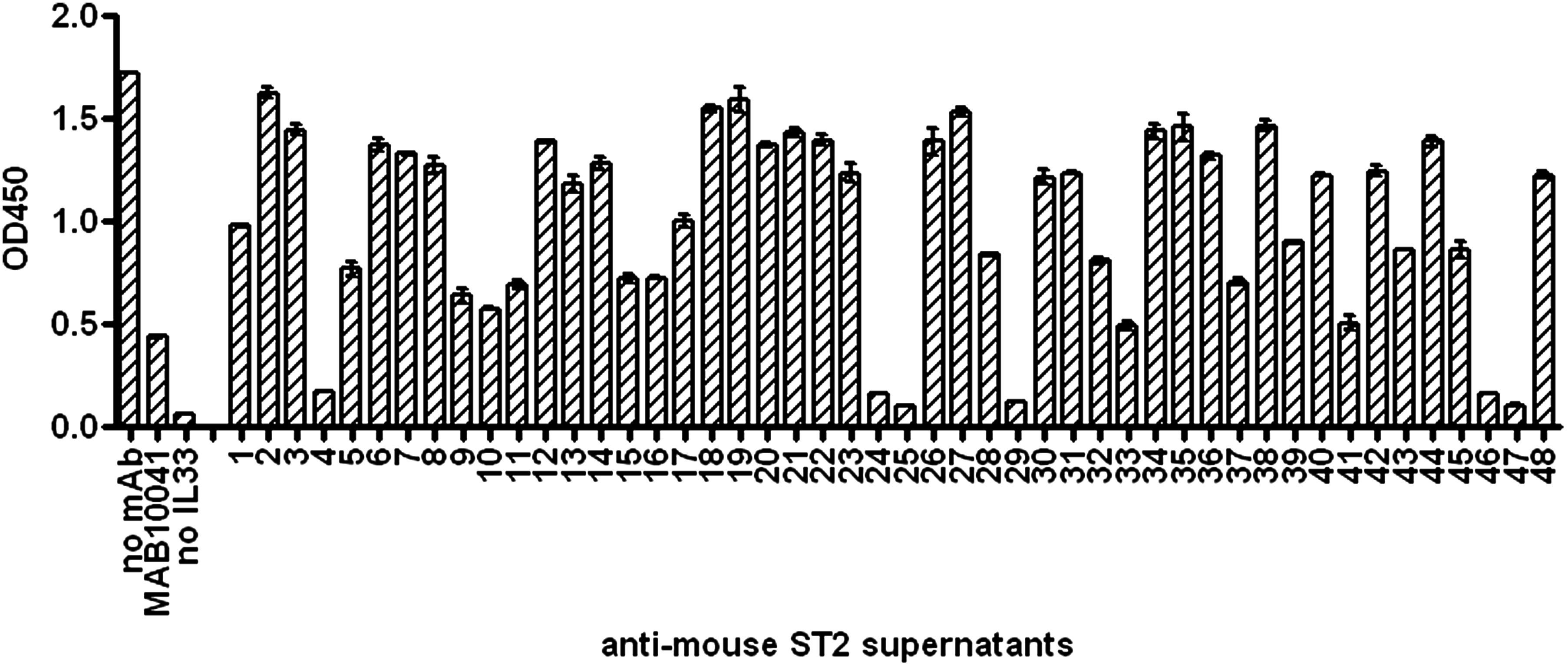
Screening anti-ST2L hybridoma supernatants in the receptor-ligand binding ELISA. Hybridoma supernatants containing anti-mouse ST2 antibodies were added to the ST2-Fc chimera presented on the assay plate following the addition of 1 μg/mL biotinylated mIL-33. Controls containing 10 μg/mL of MAB10041, no IL-33, and no antibody were added to spent hybridoma medium in the volume equal to the volume of tested hybridoma supernatants. Data are presented as average from duplicate wells (n = 2) with error bars representing variability.
All 48 anti-ST2L clones were also tested for the ability to neutralize mIL-33-induced proliferation of D10.G4.1 cells, a mouse TH2 cell line. We confirmed that D10.G4.1 cells do express mST2L on the cell surface by flow cytometry (data not shown), and R&D Systems have reported that their recombinant mIL-33 protein does induce D10.G4.1 cell proliferation. The cells were treated with various concentrations of mIL-33, and they demonstrated a dose-dependent increase in proliferation as measured by the amount of intracellular ATP in a Promega CellTiter-Glo assay (Fig. 2). Anti-mST2L hybridoma supernatants were added to D10.G4.1 cells in combination with mIL-33 and cell proliferation was measured after 24 h (Fig. 3). All six supernatants identified as strong inhibitors in the receptor-ligand binding assay exhibited strong anti-proliferative activity in the cell-based assay as well. In addition to the six strong neutralizers, there were supernatants exhibiting various degrees of inhibition in both the receptor-ligand binding and cell proliferation assays. Supernatants 18, 26, and 27 inhibited cell proliferation but had no effect in the receptor-ligand binding assay and therefore could be inhibitors of IL-1RAcP binding to the ST2L/IL-33 complex. Supernatants 13, 14, and 40 enhanced cell proliferation above the level induced by mIL-33 alone. This effect could be caused by mitogenic agents secreted by hybridomas and present in supernatants or by anti-mST2 agonistic antibodies. Considering the complexity of the hybridoma supernatants, we understood that screening results may identify false positive or false negative hits, especially in cell-based assays assessing cell viability/proliferation. Thus, purified antibody preparations were needed to test for functional activity against the target. Based on the neutralization efficiency in both the receptor-ligand ELISA and D10.G4.1 proliferation assay, 20 clones were selected for establishment of monoclonal cultures. Eleven stable hybridoma clones were generated and monoclonal antibodies were purified to be further characterized in vitro.
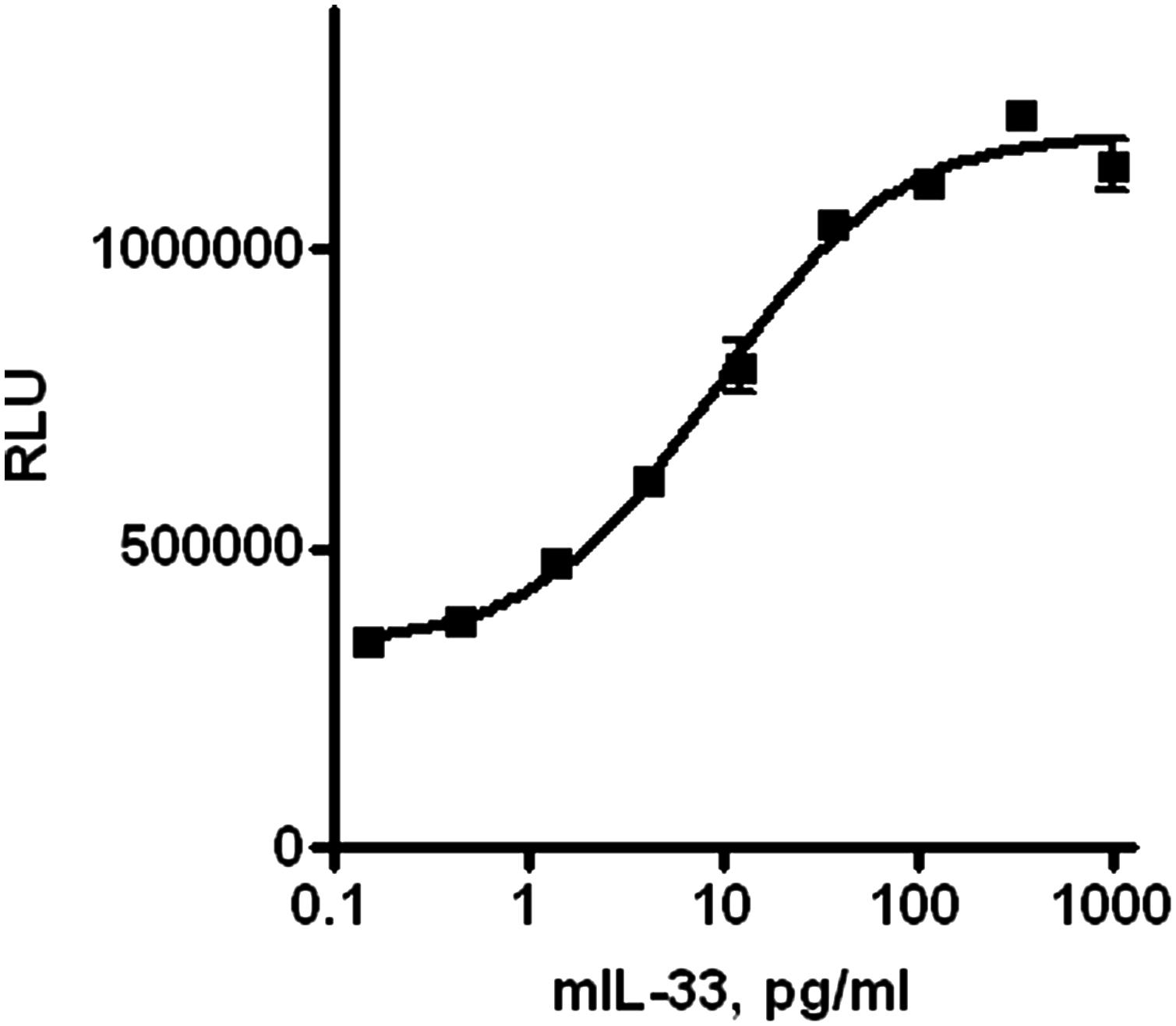
IL-33 caused increase in D10.G4.1 cell proliferation in dose-dependent manner. Cells were seeded in a 96-well plate, treated with various amounts of mIL-33 for 24 h. Proliferation was measured using CellTiter-Glo reagent (Promega). Data are presented as average from duplicate wells (n = 2) with error bars representing variability. One representative experiment out of three independent experiments is shown.

Screening anti-ST2L hybridoma supernatants in the D10.G4.1 mouse TH2 cell proliferation assay. Hybridoma supernatants containing anti-mouse ST2 antibodies were added to the cells on a 96-well plate following addition of 20 pg/mL mIL-33. Controls containing 1000, 100, and 20 pg/mL mIL-33, 10 μg/mL MAB10041, rat IgG (ITC; isotype control), and no IL-33 were added to spent hybridoma medium in the volume equal to the volume of tested hybridoma supernatants. Cellular level of ATP as a factor of proliferation was measured using CellTiter-Glo reagent (Promega) 24 h after treatment. Data are presented as average from duplicate wells (n = 2) with error bars representing variability.
Antibody in vitro characterization
ST2 receptor binding
Eleven purified anti-ST2L MAbs were tested for the binding to mST2 receptor in an ELISA. As shown on the Figure 4, all 11 anti-ST2 MAbs (but not isotype control rat IgG) bound to mST2-Fc coated on the plate. No binding was detected in wells coated with irrelevant protein (data not shown).

Binding of purified antibodies to mST2-Fc receptor by ELISA. Various amounts of purified anti-mouse ST2 antibodies were added to the mST2-Fc protein coated on the plate. Antibodies were detected using an anti-rat HRP-conjugated antibody. Data are presented as an average from duplicate wells (n = 2) with error bars representing variability. One representative experiment out of two independent experiments is shown.
Receptor-ligand binding inhibition assay
Antibody functionality against the ST2 receptor was tested in a number of biochemical and cell-based assays. The monoclonal antibody designated as CNTO8718 did not inhibit receptor-ligand binding (Fig. 5). The rest of the antibodies showed various degrees of inhibition. CNTO2549, CNTO9383, CNTO8195, and CNTO9195 were found to be the strongest inhibitors of ST2/IL-33 binding.
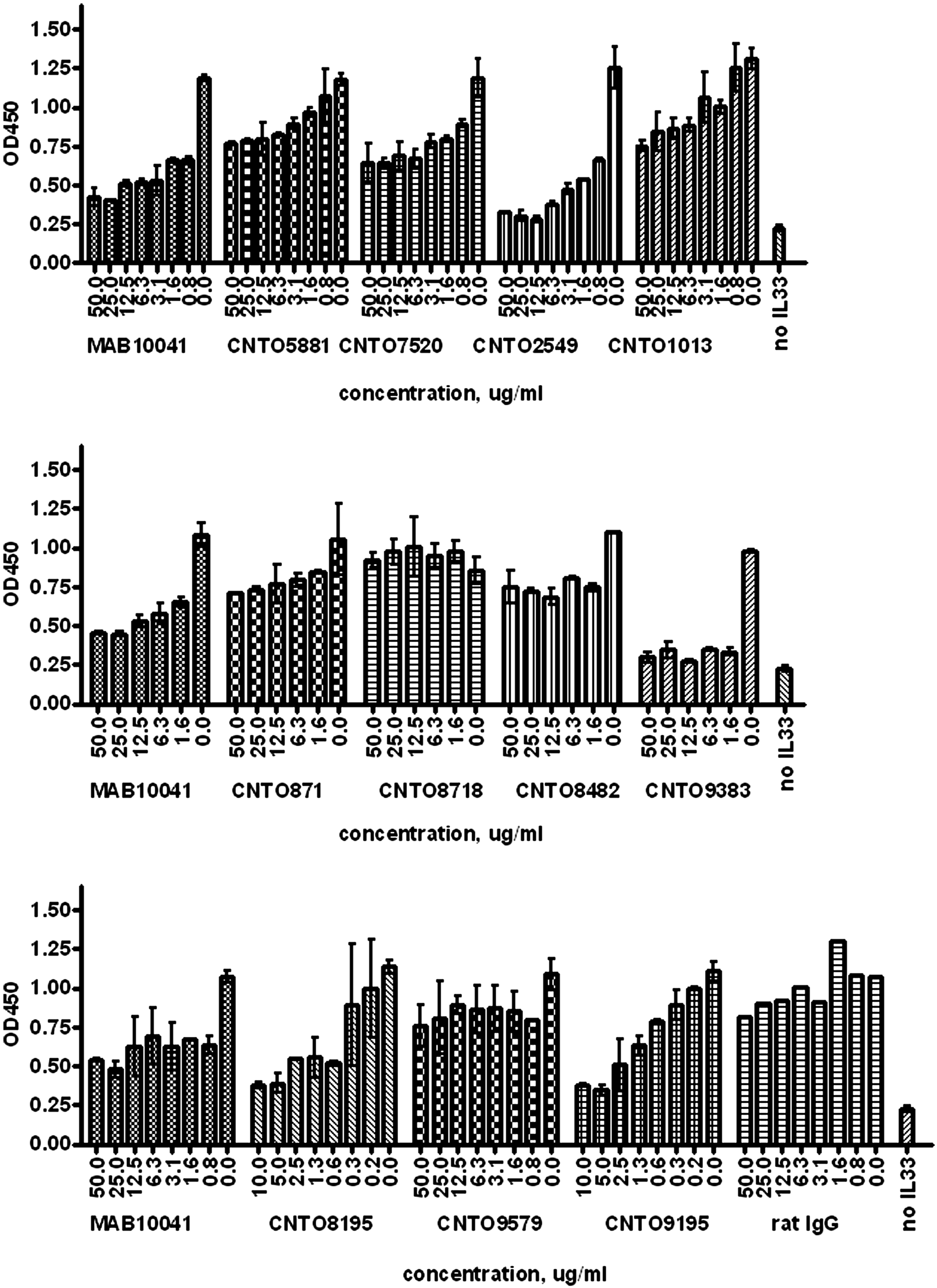
Receptor-ligand binding inhibition assay. Various amounts of purified anti-mouse ST2 antibodies were mixed with biotinylated mIL-33 and added to the mST2-Fc presented on the plate. The detection was done using streptavidin-conjugated HRP. Antibodies were tested on three plates with MAB10041 and no IL-33 controls on each plate. Data are presented as average from duplicate wells (n = 2) with the exception of isotype control-containing wells. Error bars represent variability. One representative experiment out of two independent experiments is shown.
Reporter gene assay
We developed a cell-based assay to measure the effect of anti-mouse ST2L antibodies on mIL-33-induced mST2L signaling. As IL-33 is known to signal through the NF-κB pathway,(8) we set up a reporter gene assay that measures transcriptional activity of NF-κB in response to mIL-33 stimulation. HEK293 cells were transiently transfected with a mouse ST2L receptor expression vector together with a luciferase reporter gene construct driven by an NF-κB-inducible promoter. Using transfected cells, we demonstrated that mIL-33 caused a dose-dependent increase in NF-κB activity in an mST2L-specific manner (Fig. 6), confirming that mST2L is able to signal as a heterodimer with human IL-1RAcP that is endogenously expressed by the HEK293 cells.
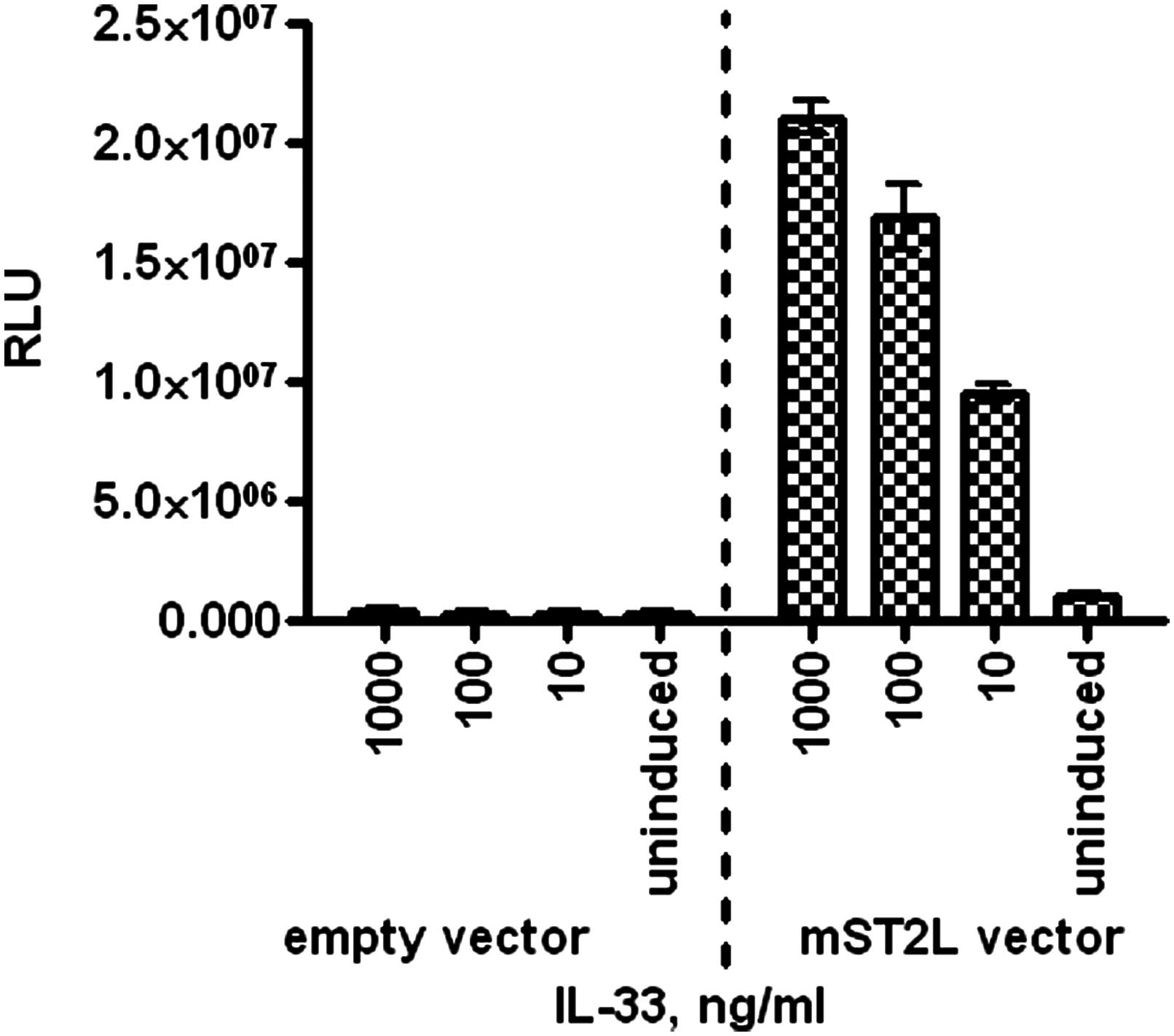
HEK293-mST2L reporter gene assay. HEK293 cells were co-transfected with mST2L or empty vector along with the NF-κB-inducible luciferase reporter vector and stimulated with mIL-33 for 16 h. Reporter gene activity was measured with SteadyGlo reagent (Promega). Data are presented as an average from triplicate wells with error bars presenting standard deviation.
Next, the panel of anti-mouse ST2L antibodies was tested for their ability to inhibit mST2L-specific NF-κB activation. Four antibodies (CNTO2549, CNTO9383, CNTO8195, and CNTO9195), which were shown to be the strongest inhibitors of the receptor-ligand interaction in the biochemical assay, were confirmed as neutralizers of ST2L signaling in the cell-based reporter gene assay (Fig. 7). The antibody panel was also tested in the reporter gene assay in the absence of mIL-33 for agonistic activity. Two of the non-neutralizing MAbs, CNTO7520 and CNTO8482, were found to strongly increase NF-κB activity in the absence of IL-33 in concentration-dependent manner (Fig. 8).

Testing anti-mST2 antibody neutralizing activity in HEK293-mST2L reporter gene assay. Various amounts of antibodies were added to transfected cells following by mIL-33 addition and a 16-h incubation. Reporter gene activity was measured with SteadyGlo reagent (Promega). Antibodies were tested on three plates with MAB10041 and no IL-33 controls on each plate. Data presented as average from duplicate wells (n = 2) with error bars presenting variability. One representative experiment out of two independent experiments is shown.
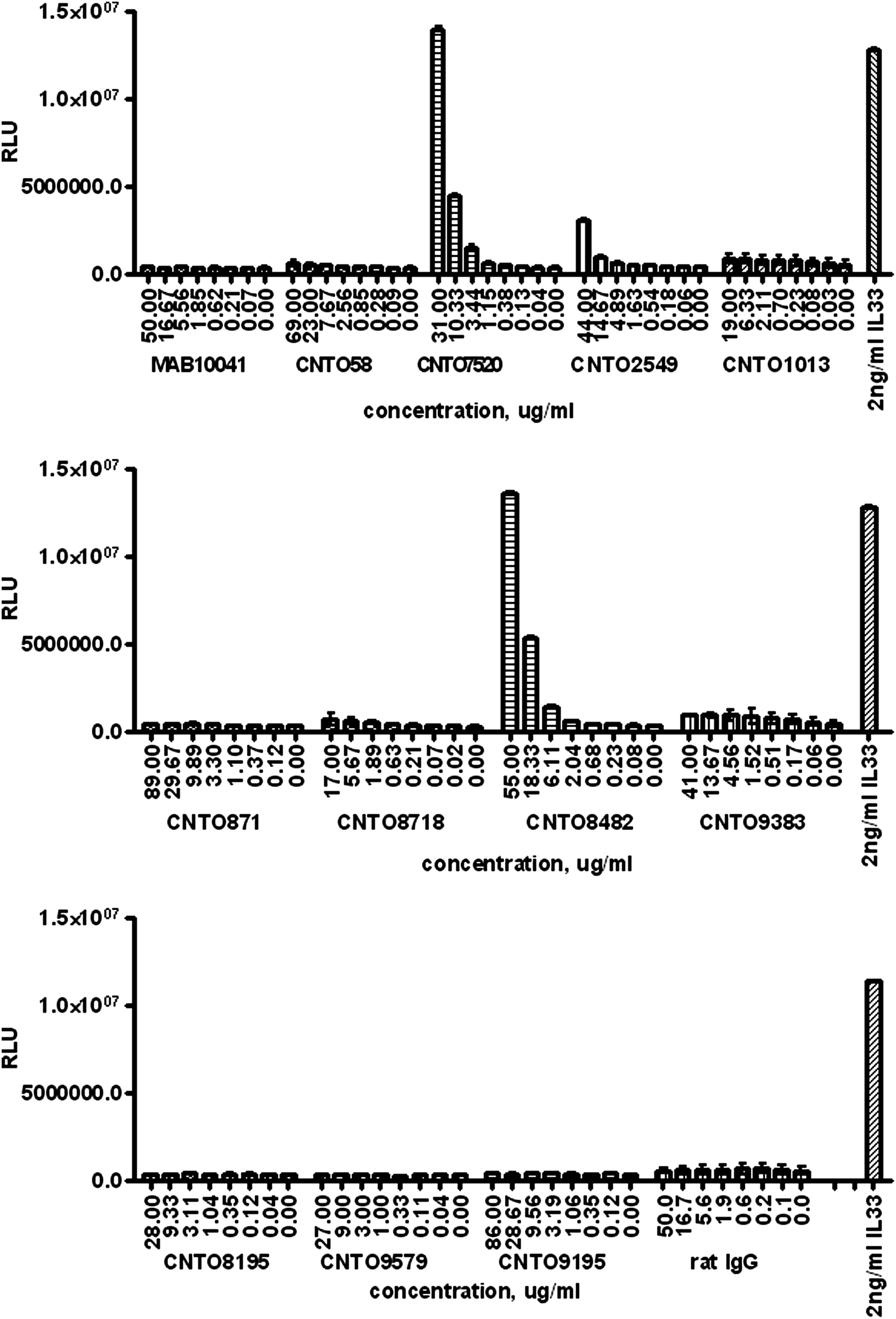
Testing anti-mST2 antibody agonistic activity in HEK293-mST2L reporter gene assay. Antibodies were added to the transfected cells at various concentrations. mIL-33 was added to separate wells as positive control. Plates were incubated for 16 h and reporter gene activity was measured with SteadyGlo reagent (Promega). Antibodies were tested on three plates with mIL-33 controls on each plate. Data are presented as average from duplicate wells with error bars representing variability.
Mouse TH2 cell proliferation assay
The mouse TH2 cell proliferation assay that was used for initial functional screening of anti-ST2L hybridomas was also used for characterization of purified antibodies. All 11 purified anti-mouse ST2L MAbs were tested for the ability to inhibit mIL-33- induced proliferation of D10.G4.1 cells. An amount of mIL-33 that caused a sub-maximal increase in cell proliferation was used in the antibody neutralization experiment. Again, four antibodies identified in previous assays as ST2L inhibitors exhibited neutralization of D10.G4.1 cell proliferation (Fig. 9). CNTO9383 showed the most potent and complete inhibition of mIL-33-induced proliferation. All other MAbs tested showed a low level or no inhibition of D10.G4.1 cell proliferation (data not shown). None of the antibodies were found to induce cell proliferation when tested in the absence of mIL-33 (data not shown).
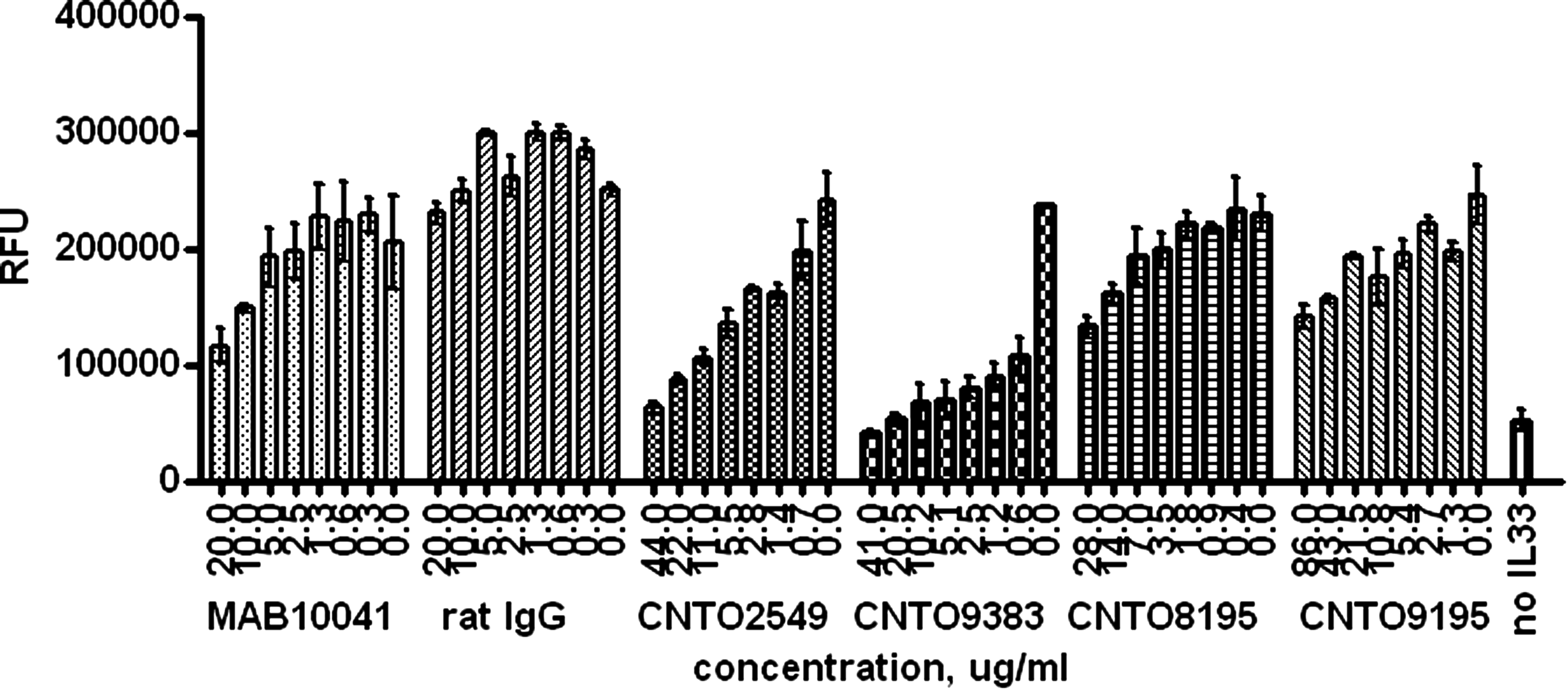
Testing anti-mST2 antibody neutralizing activity in mouse TH2 cell proliferation assay. Varying amounts of antibody were added to D10.G4.1 cells followed by mIL-33 addition and a 24-h incubation. Cellular level of ATP as a factor of proliferation was measured using CellTiter-Glo assay (Promega). Data are presented as average from triplicate wells with error bars representing standard deviation.
Discussion
The goal for this study was to generate a panel of monoclonal anti-mouse ST2L antibodies and to select antibodies inhibiting mIL-33 induced mST2L activity. The panel of antibodies was generated by immunization in rats with recombinant mST2L-extracellular domain (ECD) protein. The hybridoma antibodies were screened as supernatants for binding to mST2L-ECD by ELISA. mST2L-ECD binding clones were identified and further screened in a set of functional assays to select candidates neutralizing mST2L activity. Based on the mechanism of IL-33-induced ST2L activation, there are at least two theoretical classes of anti-ST2L neutralizing antibodies. One mechanism could be binding to ST2L and preventing it from binding to IL-33. A second mechanism could be to prevent IL-1RAcP from binding to the ST2L/IL-33 complex and subsequently inhibit downstream signaling. Each of the mechanisms of neutralization may include either direct or allosteric inhibitors. The set of functional screening assays used in this study included biochemical receptor-ligand binding, reporter gene activation, and TH2 cell proliferation assays. All inhibitors identified in the receptor-ligand binding assay were neutralizers of the ST2/IL-33 interaction. The cell-based proliferation assay allowed identification of neutralizing antibodies inhibiting either receptor-ligand binding or interactions of the IL-1RAcP with the receptor-ligand complex. Candidate selection was open to both types of neutralizing epitopes and was initially based on neutralization efficiencies that may reflect antibody epitope, affinity, and/or concentration in the hybridoma supernatants. However, it is interesting to note that this method of antibody generation only yielded neutralizers with one mechanism: inhibition of ligand binding. It is not known whether the antigen used for immunizations influenced the type of neutralizing antibodies generated. Another possibility is that the epitopes on ST2 important for IL-1RAcP recruitment to the ST2/IL-33 complex are not exposed in the absence of ligand.
After initial screening, anti-mouse ST2L binders that exhibited neutralizing activity in crude supernatants were subjected to clonal selection and purification. Purified antibodies were characterized for the binding and functionality against mST2L in vitro. All of the purified antibodies were shown to bind to mST2L by ELISA. Biochemical receptor-ligand binding inhibition assays identified four antibodies that inhibited ≥90% of mIL-33 binding. We also tested the purified antibodies for the inhibition of mIL-33-induced mST2L-dependentNF-κB activation and mIL-33 induced TH2 cell proliferation assays. Both cell-based assays allowed us to select ST2L inhibitors blocking mIL-33 binding to the receptor or inhibitors blocking IL-1RAcP binding to the receptor-ligand complex. While the mIL-33-induced TH2 cell proliferation assay reflects mST2L activity relevant to the role of the receptor in vivo, the NF-κB reporter gene assay is a mechanistic assay that models mST2L signaling in human HEK293 cells that do not express the human ST2L receptor endogenously. However, we confirmed that HEK293 cells express human IL-1RAcP (data not shown) and we have demonstrated that mIL-33 induces activation of NF-κB in mST2L-transfected HEK293 but not in untransfected cells. We incorporated the NF-κB reporter gene assay as an additional test that allow us to measure antibody effect on mIL-33-induced mST2L signaling and to confirm antibody activity seen in the TH2 cell proliferation assay.
The functional cell-based assays confirmed CNTO2549, CNTO9383, CNTO8195, and CNTO9195 as neutralizing antibodies, with CNTO2549, CNTO8195, and CNTO9195 being less potent when compared to CNTO9383 in all the assays tested. Two non-neutralizing antibodies, CNTO7520 and CNTO8482, showed agonistic activity in the reporter gene assay in the absence of IL-33. Interestingly, the same antibodies did not cause increase in TH2 cell proliferation when tested in the absence of mIL-33. Antibody-mediated receptor agonism may be an effect specific to HEK293-mST2L transfected cells that overexpress receptor on the cell surface compared to the endogenously expressed mST2L in D10.G4.1 cells. Antibody bound to certain epitopes on the high density receptor on the cell surface may result in receptor cross-linking and subsequent activation. Alternatively, ST2L signaling in the HEK293 cells may be dysregulated in this artificial system compared to endogenously expressing D10.G4.1 cells. CNTO2549, CNTO9383, CNTO8195, and CNTO9195 did not show agonistic activity when tested in either the reporter gene assay or TH2 proliferation assay in the absence of mIL-33.
Based on the in vitro characterization tests, we selected anti-mST2L antibody CNTO9383 as the best choice for in vivo studies. This antibody showed 53 pM affinity for binding to mST2L-ECD as measured by Biacore (data not shown). CNTO9383 blocked mIL-33 from binding to mST2L and demonstrated more than 90% inhibition of IL-33 effects in all biochemical and cell-based assays. We propose to use this anti-mouse ST2L neutralizing antibody in a number of in vivo models to assess the functionality of the mouse ST2L receptor.
Author Disclosure Statement
The authors have no financial conflicts to disclose.