
Abstract
Splenocytes obtained from mice immunized with whole purified elementary bodies of Chlamydia trachomatis were used for hybridoma construction. The resulting clones were screened with ELISA using chlamydial lipopolysaccharide (LPS) and full-length human apolipoprotein B (ApoB). One analyzed clone producing IgG1 (MAb 7B5) showed simultaneous recognition of chlamydial LPS and human ApoB, suggesting the presence of common antigenic epitopes in their structures. MAb 7B5 exhibited agreeable activity in immunoblot analysis conducted using chlamydial extracts or full-length human ApoB as well as in immunofluorescence (IF) detecting typical inclusion bodies of C. trachomatis and C. pneumoniae in the infected eukaryotic host cells. The removal of LPS from chlamydial suspensions with lauroyl sarcosyl led to a complete disappearance of IF associated with the elementary bodies of C. trachomatis. Therefore, immunologic response to chlamydial antigen may be associated with the generation of ApoB-specific antibody. Molecular mimicry and subsequent formation of cross-reactive antibodies might be an essential mechanism explaining the appearance of circulating auto-antibodies against low density lipoproteins (LDL) in patients with atherosclerosis. Moreover, newly generated MAb 7B5 can be a useful tool in the laboratory diagnosis of chlamydial infections.
Introduction
In the present study, we report a new monoclonal antibody recognizing chlamydial lipopolysaccharide (LPS) with cross reactivity to human ApoB. The hybridoma was constructed using splenocytes from mice immunized with elementary bodies (EBs) of Chlamydia trachomatis.
Materials and Methods
Bacterial culture
All reagents were obtained from Sigma-Aldrich (St. Louis, MO) unless specifically mentioned otherwise. Strain L2/Bu434 of C. trachomatis was kindly provided by Dr. P. Saikku (University of Oulu, Finland). The pathogen was propagated in McCoy cells and purified by renografin gradient centrifugation as widely described.(5) Purified EBs were suspended in sucrose-phosphate-glutamic acid buffer and kept frozen until immunization or tissue culture work.
Production of MAb
All animal procedures were approved by the local Animal Research Ethical Committee. BALB/c mice were intraperitoneally immunized with UV-inactivated EB suspensions of C. trachomatis with Freund's adjuvant. After 72 h following the final intravenous boost, the spleens of anesthetized mice were removed and dispersed and the resulting splenocytes were fused with Sp-2 cells and incubated in HAT medium according to conventional protocol.(6) Positive wells were inspected microscopically for cluster formation and the supernatants were tested for antibody presence by ELISA. Promising clones were subcloned by limiting dilution protocol using spleen feeders. Antibodies were isotyped with Sigma mouse monoclonal antibody isotyping reagent.
ELISA testing was conducted using Nunc MaxiSorp flat-bottom 96-well plates (eBioscience, Hatfield, United Kingdom) coated with chlamydial LPS or human ApoB. Chlamydial LPS was isolated and purified from EBs of C. trachomatis as described previously.(7) The plates were coated by the addition of 7 μg/mL of LPS (or native human ApoB; #A5553) reconstituted in carbonate buffer (pH 9.6). After washing with PBS containing 0.05% Tween-20 (PBST) and blocking with 0.1% BSA in PBS for 30 min at 37°C, 0.1 mL of incubation medium was added to the wells. Next, the plates were washed with PBST with the subsequent addition of goat-anti mouse secondary IgG-HRP antibody at 1:1000 dilution in PBS and 1 h incubation at 37°C. Washed plates were exposed to TMB substrate and the reaction was stopped by the addition of 1 N H2SO4. ELISA results were documented on the plate reader at 450 nm.
Immunoblot analysis
EBs of C. trachomatis or human ApoB were heated at 95°C in 62.5 mM Tris-HCL (pH 7.8) containing 4 M urea, 10% SDS, 10% glycerol, 100 mM DTT, and 0.05% bromophenol blue. Aliquots of protein (20 μg per lane) were loaded on 4−15% gradient SDS polyacrilamide gel and electrophoresed at 200 V. Proteins were transferred on Hybond nitrocellulose membranes (Amersham, Piscataway, NJ) and blocked overnight at 4°C in 5% non-fat dry milk in PBST. Primary antibody solutions (5 μg/mL in PBST) were added to the membranes with further incubation at room temperature for 2 h. After three 15 min washes in PBST, the membranes were exposed to anti-mouse HRP-conjugated IgG (1:1000 dilution, Amersham), washed again, incubated in SuperSignal West Substrate Working Solution (Pierce, Rockford, IL), and exposed to X-ray film. Polyclonal antibody against human ApoB (Abcam, Cambridge, UK) was used as positive control for immunoblotting.
Immunofluorescent assay
Direct immunofluorescence (IF) staining was conducted in McCoy cell monolayers infected with reference culture of C. trachomatis as well as in HL cells infected with C. pneumoniae. MAbs were labeled using FITC antibody labeling kit (Pierce). FITC-conjugated MAb against major outer membrane protein (MOMP) of C. trachomatis (NearMedic, Moscow, Russia) was used as irrelevant control. The cells were grown for 48 h on coverslips in 24-well plates after inoculation of pathogens at multiplicity of infection of 2:1. Monolayers were fixed with methanol, permeabilized with 0.1% Triton X-100, and blocked in 0.1% BSA in PBST. After antibody addition (5 μg/mL in PBST), the coverslips were incubated for 2 h at 37°C, washed with PBST, and visualized under Nikon Eclipse 50 fluorescence microscope at x1350 magnification.
Specificity of the newly generated antibody was also determined using the sodium lauroyl sarcosinate (sarcosyl) treatment protocol, which is known to remove LPS from chlamydial suspensions.(5) Briefly, purified elementary bodies of C. trachomatis were centrifuged at 100,000 g for 20 min, and the resulting pellet was gently resuspended with 2% lauroyl sarcosyl in PBS buffer (pH 8.0, 1.5 mM EDTA) or PBS alone. The tubes were placed on a rotating wheel for 1 h at 37°C. After washing in PBS, the cell suspensions were analyzed in IF assay with the newly generated FITC-labeled antibodies against chlamydial LPS or commercial antibody against MOMP.
Results
A multiple step cloning strategy has been employed in our work. Firstly, we used a corpuscular antigen containing a whole array of C. trachomatis-specific constituents to obtain immune splenocytes for the generation of specific hybridoma clones. After constructing hybridoma clones, we succeeded in getting multiple individual clones producing antibodies with specific recognition of chlamydial LPS in ELISA. Clones with the highest reactivity to LPS were screened in ELISA test with human ApoB and further expanded. Finally we narrowed our choice to a single hybridoma clone identified as MAb 7B5, which produced IgG1 recognizing both antigens of interest (chlamydial LPS and human ApoB) in immunoblot analysis, as well as in ELISA. To our advantage, MAb 7B5 seems to work very efficiently in direct IF detection of Chlamydia-specific antigen.
As seen in Figure 1, MAb 7B5 was capable of staining chlamydial inclusion bodies generated in the host cells by both chlamydial species (C. trachomatis and C. pneumoniae). Direct IF protocol allowed registering inclusion body signal formation in McCoy (C. trachomatis) and HL cells (C. pneumoniae) as early as 20 h into the post-infection period. The initial IF signal was associated with punctuate and granular structures in slightly opalescent perinuclear areas of the host cells. Typical inclusion morphology started to emerge at 48 h of the post-infection period. C. trachomatis inclusions were large with irregular borders not uniformly stained with some empty pockets. C. pneumoniae inclusions had regular borders, were also smaller in size, and were surrounded by multiple singular EBs. In both cases, the inclusion formation was accompanied by nuclear dislocation in the host cells. An immunohistochemical signal in extracellular space started to appear after 48 h of the post-infection period when the lytic phase of the infection cycle became apparent. The signal was represented by granular structures resembling typical chlamydial EBs in their size and shape. Uninfected cells remained entirely IF negative with no background staining.

Immunofluorescence assay with newly generated anti-LPS MAb 7B5 in cultured cells infected with chlamydial pathogens. McCoy (
Next we checked if removal of LPS with lauroyl sarcosyl diminishes the intensity of IF staining found in the reference culture of C. trachomatis. Figure 2 shows that LPS removal from bacterial suspensions completely abolished IF signal associated within EBs. On the other hand, EBs remained strongly positive for irrelevant protein constituent (MOMP) regardless of LPS elimination (Fig. 2C). This implies high specificity to LPS and the lack of background activity for MAb 7B5.

Immunofluorescent staining in C. trachomatis suspensions treated with lauroyl sarcosyl. (
After this experiment, we proceeded with immunoblot characterization of MAb 7B5. As seen in Figure 3, MAb 7B5 produced identical multiple band recognition complexes in PAGE conducted with the extracts of both chlamydial pathogens. Such immunoblot recognition pattern with multiple “ladder-like” bands is well known for chlamydial LPS in SDS-PAGE.(8) The lower band was positioned close to 17 kDa, which allowed positive identification of the bands as LPS. Therefore, the immunoblot analysis is consistent with our clone screening strategy, based on choosing hybridomas recognizing LPS in ELISA, and supports immunohistochemistry results revealing the ability of MAb 7B5 to detect chlamydial LPS. Moreover, MAb 7B5 showed recognition of a single band positioned at the very top of the gel (Fig. 3, lane 3), which presumably is human ApoB. The identity of the band becomes clear when the right side of the panel, blotted with anti-ApoB antibody, is considered (Fig. 3, lanes 4–6). Antibody against human ApoB does not produce recognition bands in chlamydial extracts. However there is an obvious recognition of ApoB in the last lane. Therefore, immunoblot analysis convincingly reveals cross reactivity of MAb 7B5 to chlamydial LPS and human ApoB. Supporting data were obtained with ELISA conducted using purified chlamydial LPS and human ApoB (Fig. 4).
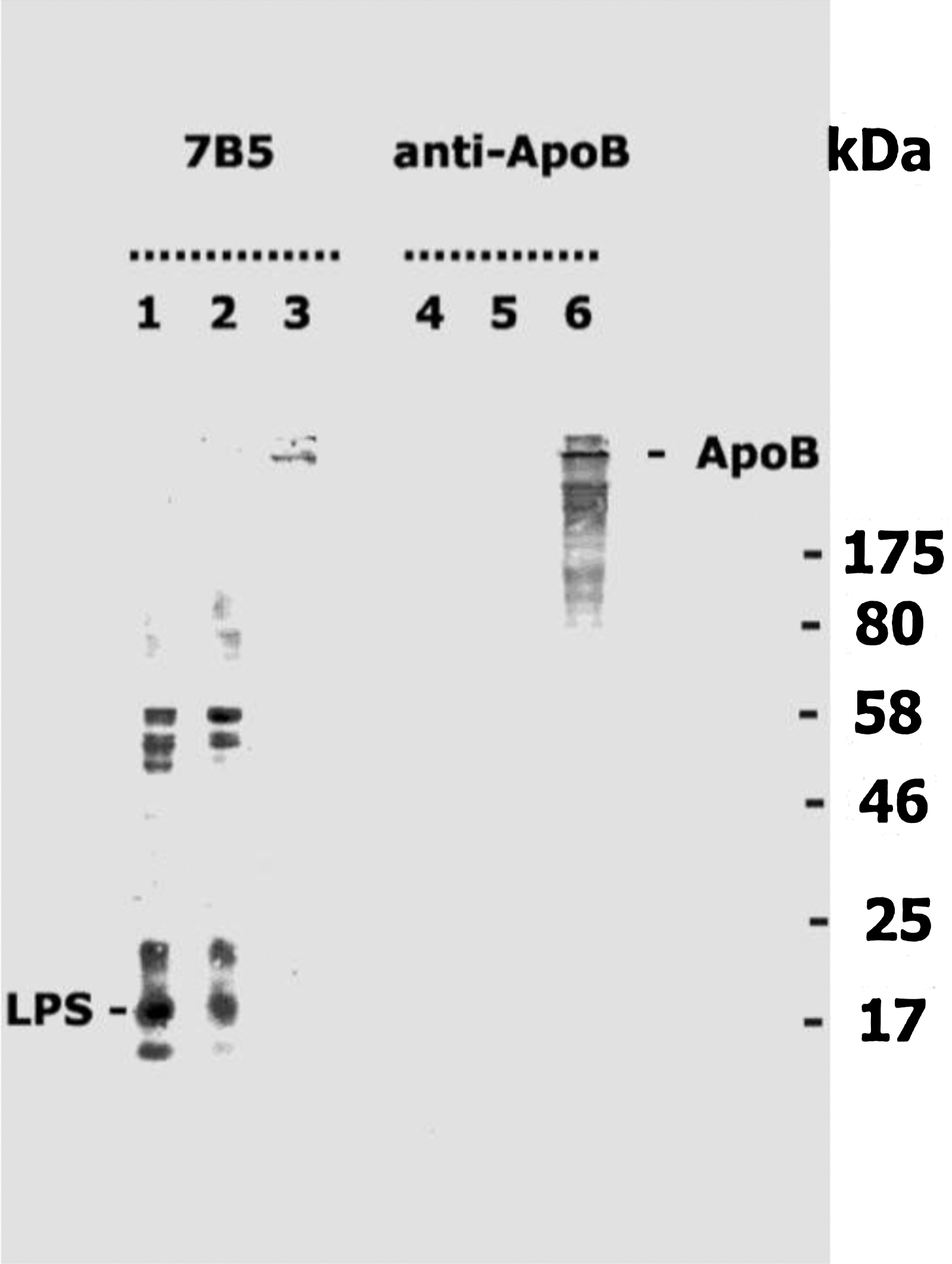
Immunoblot analysis using newly generated anti-LPS MAb 7B5 versus commercial antibody against human ApoB. Lanes 1–3 were blotted with MAb 7B5; lanes 4–6 were blotted with commercial antibody against human ApoB. Lanes 1 and 4 contain 20 μg of C. trachomatis whole cell lysate (see Materials and Methods section); lanes 2 and 5 contain 20 μg of C. pneumoniae lysate; lanes 3 and 6 contain 10 ng of human ApoB.
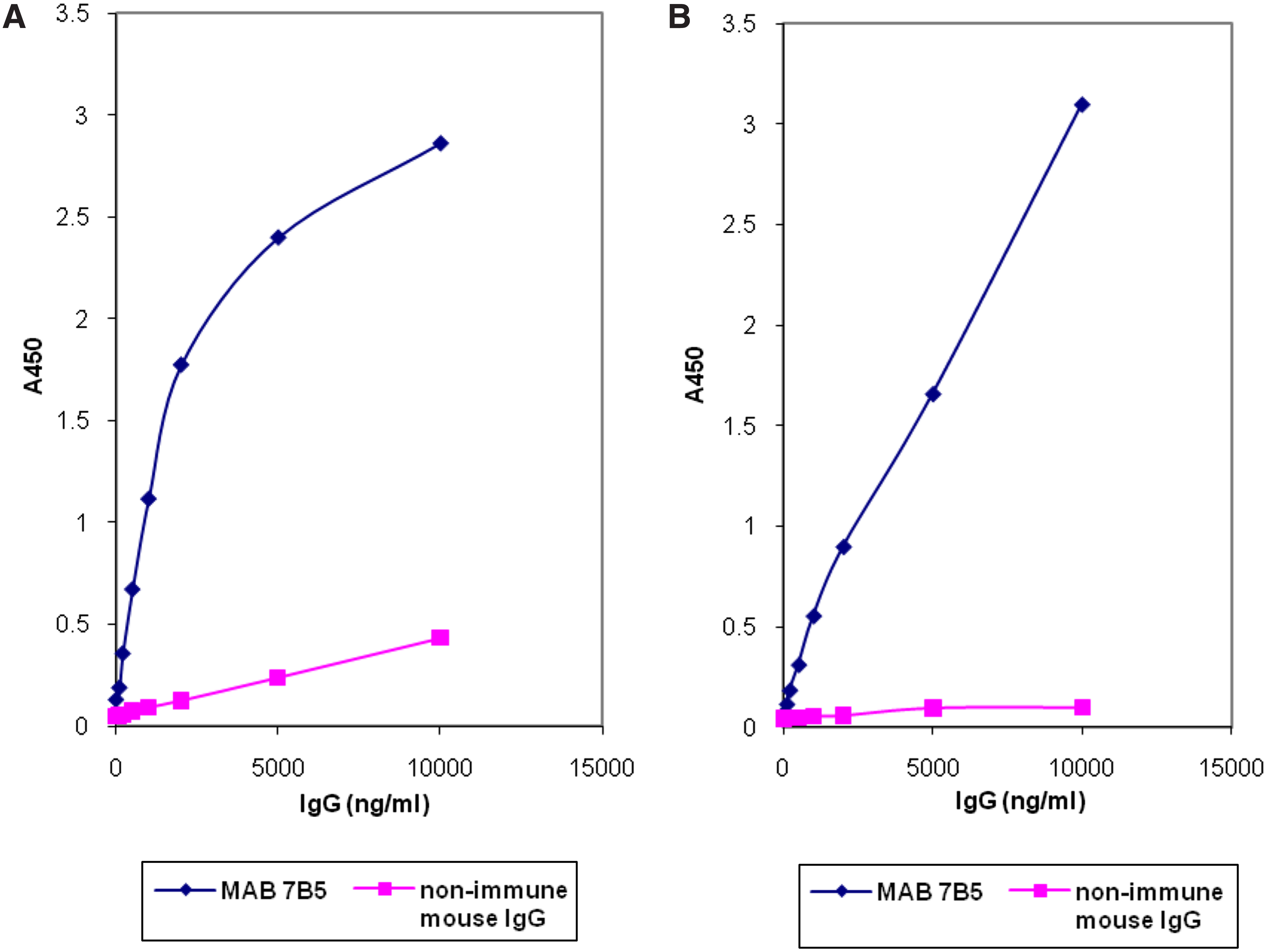
Recognition of chlamydial LPS (
Discussion
LPS is a universal constituent of the outer envelope in gram-negative bacteria including the genus Chlamydia. Chlamydial LPS plays an extremely important role in the pathogenesis of chlamydiosis. Despite relatively weak endotoxic activity, chlamydial LPS retains high mitogenic activity, triggering a cascade of immunologic reactions in infected individuals.(9) The ability to induce cytokine production (TNFα, IL-1, IL-6, IL-12), pro-inflammatory, pro-coagulating, and pyrogenic effects are other crucial features of LPS.(10–12) Its core structure and genus-specific epitopes are fairly well conserved in spite of some variations among different chlamydial species.(13)
In the present study, we report a new genus-specific monoclonal antibody against chlamydial LPS raised using bacterial suspensions of C. trachomatis. The resulting antibody cross-reacts with native human ApoB. MAb 7B5 showed an agreeable activity in indirect IF protocol detecting LPS in C. trachomatis and C. pneumoniae in the infected host cells as well as in immunoblot analysis conducted using chlamydial extracts and human ApoB. Obtained results were confirmed with ELISA. Newly produced antibody might be a useful tool in the laboratory diagnostics of chlamydial infections. Overall appearance of chlamydial intracytoplasmic vacuoles visualized with MAb 7B5 in HL and McCoy cells corresponds to the classic inclusion body morphology described for chlamydial species elsewhere.(14,15) MAb 7B5 can also be implemented in apolipoprotein analysis for detecting human ApoB in plasma and some tissues expressing Apo B mRNA (small intestine, liver, adrenal glands).
Beyond its significance in laboratory practice, there is another important dimension emerging from our work. There is a clear association between C. pneumoniae in atherosclerosis proven by sero-epidemiological analysis, detection of the organism within atherosclerotic lesions and animal experiments.(16) Whether C. pneumoniae is a cause, a co-factor, or an innocent bystander residing in atherosclerotic plaques remains unanswered and is under intensive investigation.(17) Causative relationship between Chlamydia and atherosclerosis requires complete fulfillment of Koch's postulates, which is a lengthy ongoing process. However some clinical manifestations of atherosclerosis can be explained with high confidence by the host-pathogen interaction even now. Atherosclerosis as a disease involves a combination of pathological changes—–cholesterol deposition, inflammation, fibrosis, and autoimmunity.(3,18) The appearance of ApoB-specific antibodies in plasma of the patients with atherosclerosis is a well-known detrimental feature of atherosclerosis because LDL-containing immune complexes taken up by lipid-laden macrophages are known to be massively deposited in atherosclerotic plaque.(1,19) However the mechanism triggering lipoprotein-specific antibody production remains unclear since native apolipoproteins cannot induce an antibody response due to immune tolerance.(3,4) Our observation spells out first the possibility of antibody formation against ApoB in response to bacterial antigen particularly to chlamydial LPS. This phenomenon may occur due to the presence of common antigenic epitopes in the structure of chlamydial LPS and ApoB. Therefore it becomes clear that the immunological response to chlamydial antigens may lead to the emergence of lipoprotein–specific antibodies, which is a pathophysiological feature of atherosclerosis. Molecular mimicry involving bacterial pathogens might be a crucial mechanism initiating autoimmune reactions in atherosclerotic patients. Such a conclusion agrees well with the report describing cross-reacting antibody directed to oxidized-LDL and antigenic determinants of Streptococcus pneumoniae.(20) The pathophysiological significance of cross-reactive antibodies and antigenic epitope mapping will need to be addressed in future studies.
Footnotes
Acknowledgments
The authors wish to thank Professor P. Saikku (University of Oulu, Finland) for sharing the strain L2/Bu434 of C. trachomatis. This work was supported by Cambridge Theranostics, Ltd., and Lycotec, Ltd., Cambridge, United Kingdom.
Author Disclosure Statement
The authors have no financial conflicts to disclose.