
Abstract
In addition to conventional antibodies, cartilaginous fish have evolved a distinctive type of immunoglobulin, designated as IgNAR, which lacks the light polypeptide chains and is composed entirely by heavy chains. IgNAR molecules can be manipulated by molecular engineering to produce the variable domain of a single heavy chain polypeptide (vNARs). These, together with the VHH camel domains, constitute the smallest naturally occurring domains able to recognize an antigen. Their special features, such as small size, long extended finger-like CDR3, and thermal and chemical stability, make them suitable candidates for biotechnological purposes. Here we describe the generation of two mouse monoclonal antibodies (MAbs), MAb 370-12 and MAb 533-10, that both specifically react with vNAR domains of the horn shark Heterodontus francisci. While the former recognizes a broad spectrum of recombinant vNAR proteins, the latter is more restricted. MAb 370-12 precipitated a single band from whole shark serum, which was identified as IgNAR by mass spectrometry. Additionally, we used MAb 370-12 to follow the IgNAR-mediated immune response of sharks during immunization protocols with two different antigens (complete cells and a synthethic peptide), thus corroborating that MAb 370-12 recognizes both isolated vNAR domains and whole IgNAR molecules. Both MAbs represent an affordable molecular, biochemical, and biotechnological tool in the field of shark single-domain antibodies.
Introduction
Typically vNAR domains against a wide variety of antigens were isolated by screening phage display libraries derived either from spleen or from peripheral blood lymphocytes.(8) The IgNAR-mediated immune response has been mainly characterized in two shark species, the nurse shark (G. cirratum)(1,5) and the wobbegong shark (Orectolobus maculatus).(8,9) However, not much is known about the IgNAR immune response of horn sharks (H. francisci). Their small size (<1 meter in length) and solitary behavior make it possible to keep these animals at low cost in small indoor sea water tanks contrary to nurse or wobbegong sharks, which require specialized aquarium facilities. These features make them an excellent model organism for raising vNAR antibodies for biotechnological processes.
Since animals have to be euthanized at the end of the immunization process, it would be desirable to follow the time course of the specific IgNAR-mediated immune response in response to an immunization. The monoclonal antibodies described in this report meet this need, and, in addition, represent a valuable biotechnological tool for the purification and detection of the isolated vNARs.
Material and Methods
Preparation of recombinant vNAR proteins
Variable domains (vNARs) from IgNAR shark antibodies were isolated from a cDNA library prepared by cloning cDNA derived from the spleen of an adult H. francisci horn shark into the pHEN-2 vector. Individual clones were sequenced to determine the size and variability of the vNAR library, and six representative clones determined to be independent on the basis of distinct sequence motifs in their CDR3 regions (HADYS, DIEGO, SAALAK, SAGTK, GRIL, and PIFQ) were selected for further analysis. For protein expression, colonies were transformed into Escherichia coli HB2151 cells and grown overnight in SB media containing ampicillin (50 mg/mL). Recombinant vNARs containing C-terminal c-Myc and 6x-histidine tags were extracted from the periplasmic fractions by sucrose osmotic shock, as previously described.(10) Periplasmic extracts were passed through a 0.22 μm syringe filter and purified by metal affinity chromatography using the Hi-Select Nickel Affinity Gel (Sigma-Aldrich, St. Louis, MO). Columns were packed with 1 mL of nickel matrix, washed with 5 mL of elution buffer (50 mM NaPO4 [pH 8.0], 250 mM imidazole, 0.3 M NaCl), equilibrated with 5 mL of washing buffer (50 mM NaPO4 [pH 8.0], 10 mM imidazole, 0.3 M NaCl), and eluted with 5 mL of elution buffer collected in individual fractions of 1 mL. Fractions containing recombinant vNARs were pooled, equilibrated in PBS by passage through PD-10 desalting columns (Amersham Biosciences, Uppsala, Sweden), and quantified using the Pierce BCA Protein Assay kit (Thermo Scientific, Rockford, IL). For immunization, endotoxin-free vNAR proteins were obtained by filtering with the Sartobind Q SingleSep filter (Sartorius, Goettingen, Germany), according to the manufacturer's instructions.
Immunization of mice and generation of monoclonal antibodies
One BALB/c mouse (designated as 3537) was immunized intramuscularly every third week for 3 months with 20 mg of the endotoxin-free vNAR protein SAALAK emulsified in Specol adjuvant. Sera were obtained from the mouse before and after completing the immunization protocol. Spleen, axial, and inguinal lymph nodes from the immunized mouse were pooled and fused to the myeloma cell line Sp2.0, as described previously.(11) Cells were plated in 96-well plates in RPMI media supplemented with hypoxanthine, aminopterin, and thymidine (HAT), and incubated at 37°C and 5% CO2 for 14 days. Grown colonies were picked and transferred to new 96-well plates containing HAT medium. Clones secreting anti-vNAR antibodies were identified by ELISA (as described below) and subcloned by limiting dilution in RPMI-HT media. Positive clones were adapted to RPMI low-IgG medium for production of monoclonal antibodies. The isotypes of clones were determined using the Mouse Immunoglobulin Isotyping ELISA kit (BD Biosciences, San Jose, CA) following the manufacturer's instructions.
Detection of vNAR recombinant proteins by ELISA
The reactivity of sera or MAbs against different recombinant vNAR proteins was determined by an ELISA assay. The wells of a 96-well plate were coated for 2 h at 37°C with 120 ng of purified vNAR proteins. Lama VHH proteins or 3% skim milk in PBS were used as specificity controls. After blocking for 1 h at 37°C with 3% skim milk in 1x PBS-Tween (MPBS-T; 20 mM Tris HCl [pH 7.5], 150 mM NaCl, and 0.05% Tween-20), the blocking solution was replaced by 100 μL mouse serum (diluted 1:1000 in MPBS-T) or hybridoma supernatant (diluted 1:100). After 2 h of incubation at 37°C, the plate was washed, and detection was performed using a sheep anti-mouse IgG horseradish peroxidase (HRP) conjugate at 1:10,000 dilution (Amersham Biosciences).
Western blot analysis
Purified vNAR proteins were denatured by boiling for 10 min, separated on a 12% SDS-PAGE gel, and transferred to a PVDF membrane (0.45 μm) (Amersham Biosciences). Membranes were blocked with 5% MPBS-T for 1 h at room temperature, incubated with MAb 370-12 for 1 h at room temperature, washed four times with 1x PBS/0.05% Tween-20 (PBS-T), and incubated with a sheep anti-mouse IgG-HRP conjugate at 1:5000 dilution for detection. The signal was visualized with the ECL Western Blotting Detection reagents kit (Amersham Biosciences) on Amersham Hyperfilm ECL High Performance chemiluminescence film (Amersham Biosciences).
Dot blot
Dot blots were performed by spotting 2 μL of animal serum (shark, llama, or goat) onto nitrocellulose Hybond ECL membrane (Amersham Biosciences). The membrane was dried at room temperature, blocked overnight at 4°C with 3% MPBS-T, and incubated for 2 h with mouse serum (1:1000) or hybridoma supernatant (1:10) diluted in PBS-T. Secondary antibody incubation and developing was achieved as described above for Western blot analysis.
Affinity chromatography of shark immunoglobulins
A 1 mL column with immobilized MAb 370-12 was prepared using the AminoLink Plus immobilization kit (Thermo Scientific). The column was loaded with 1 mL shark serum diluted 1:10 in PBS, washed with PBS, and eluted with elution buffer (0.1 M glycine [pH 2.7]). Eluted fractions were immediately neutralized with 1:10 volume of neutralization buffer (1 M Tris-HCl [pH 9.0]). Obtained fractions were separated on a 10% SDS-PAGE gel, transferred to a PVDF membrane, and subjected to Western blot analysis using the MAb 370-12.
Mass spectrometry assay
Eluted IgNAR molecules obtained from affinity chromatography were analyzed on 10% SDS-PAGE and stained with Coomassie blue. Bands were excised from the gel and analyzed on a QTOF2 tandem mass spectrometer (Micromass) at the mass spectrometry core facility at the University Medical Center Hamburg-Eppendorf.
Specific IgNAR immune response in sharks
To evaluate the specific IgNAR response of sharks during an immunization protocol of 149 days, two animals were independently immunized with human erythrocytes or a synthetic conotoxin. Pre-immune and immune sera (after every boost) were taken and analyzed by ELISA. Briefly, ELISA plates were incubated overnight at 4°C with 1 μg per well of glycophorin-A (the major surface protein in erythrocytes) or a synthetic conotoxin, washed, and blocked for 2 h at 37°C with 200 μL of 3% milk or BSA in PBS-T. Subsequently, 100 μL of shark serum at 1:800 dilution were added to the wells and incubated for 2 h at 37°C. The wells were washed four times, and then incubated for 2 h at 37°C with 100 μL MAb 370-12 hybridoma supernatant. After washing again, the wells were incubated for 2 h at 37°C with 100 μL of anti-mouse IgG-HRP at 1:5000 dilution and developed with 100 μL of TMB substrate (Sigma) for 30 min at room temperature in darkness. The reaction was stopped by adding 50 μL of 0.5 M sulfuric acid, and the plate was read at 450 nm.
Results
MAbs 370-12 and 533-10 specifically recognize a variety of vNAR recombinant proteins
To generate monoclonal antibodies able to react with H. francisci IgNAR and vNAR molecules, a BALB/c mouse (3537) was immunized with recombinant vNAR protein (SAALAK). At the end of the immunization period, serum from the immunized mouse (i.s. 3537) reacted with a broad spectrum of different recombinant vNAR proteins, while pre-immune serum (p.i.s. 3537) showed no reactivity (Fig. 1). After cell fusion, 395 hybridoma clones were obtained and screened by ELISA using SAALAK as antigen. Only two stable antibody-secreting clones emerged: 370-10 and 533-12. Although both clones shared the IgG1/kappa isotype, they showed differences in antigen specificity. As shown in Figure 1, MAb 370-12 specifically reacted with all the vNARs tested. However, MAb 533-10 reacted strongly with SAALAK, DIEGO, and SAGTK, but reacted weakly or not at all with HADYS, GRIL, or PIFQ. Comparison of the amino acid sequences of these clones did not reveal any evident sequence motif shared by the former three and absent in the latter three vNARs that might constitute the epitope recognized by MAb 533-10 (Fig. 2), suggesting that MAb 533-10 recognizes a conformational rather than a linear epitope.
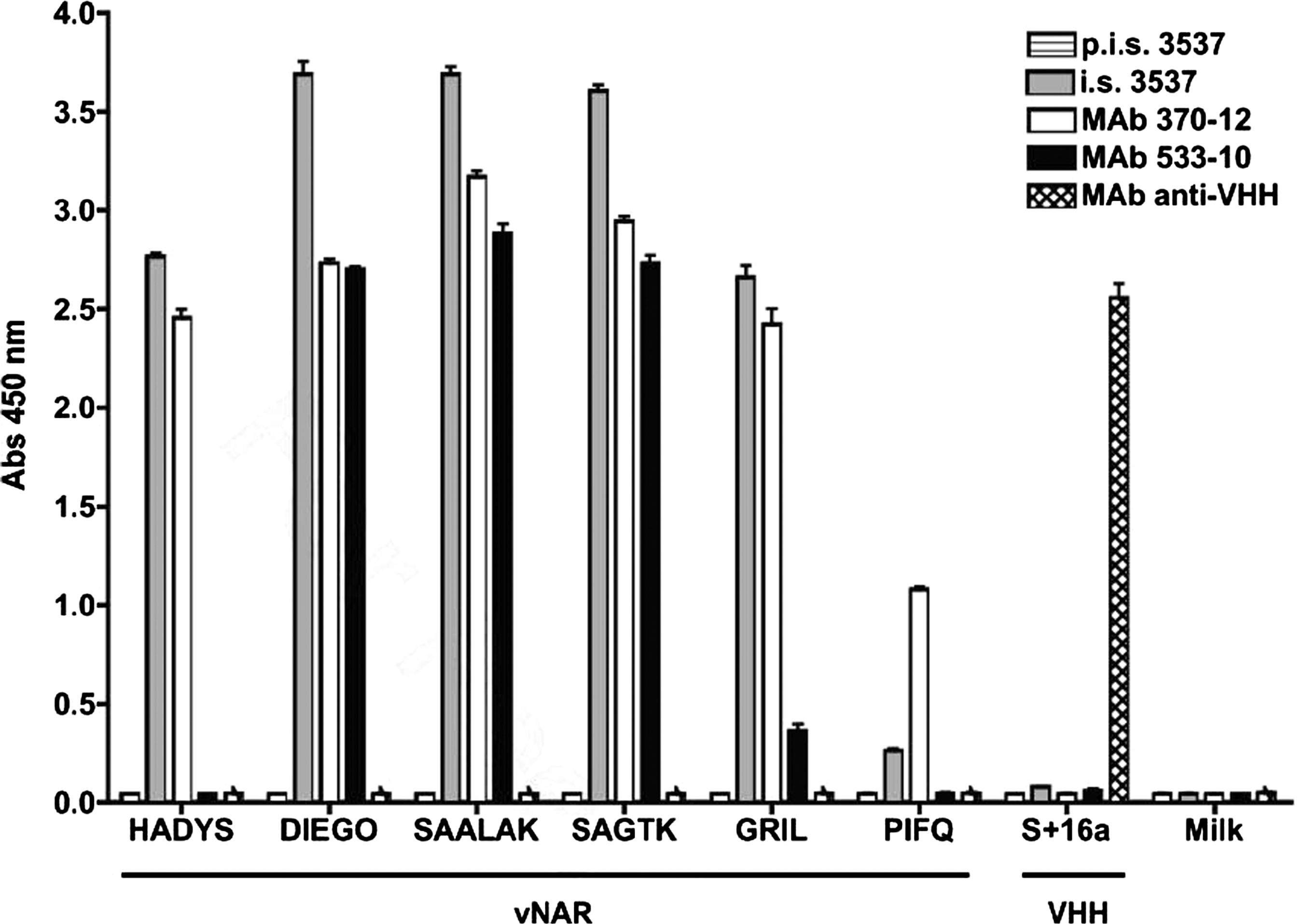
Monoclonal antibodies 370-12 and 533-10 specifically recognize a variety of vNAR recombinant proteins. The capacity of immune serum from mouse 3537 (i.s. 3537) and MAbs 370-12 and 533-10 to react with a variety of vNAR molecules or irrelevant proteins (milk or the llama VHH domain s + 16) was analyzed by ELISA. Pre-immune serum (p.i.s 3537) and an irrelevant monoclonal antibody (raised against llama VHH domain s + 16) were used as controls.
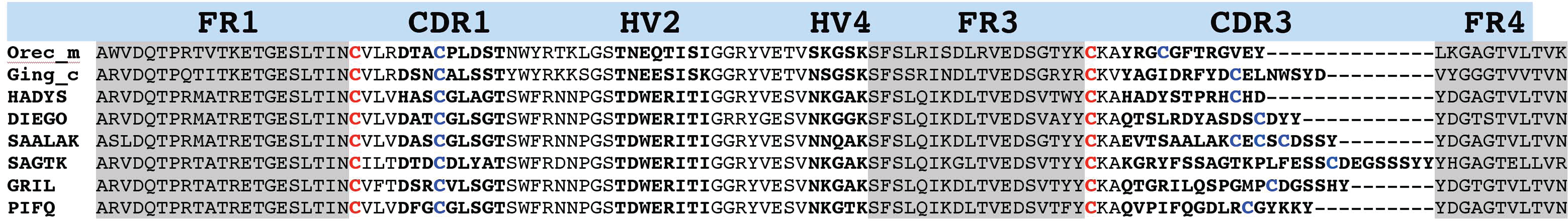
Sequence alignment of Heterodontus francisci vNAR clones. The amino acid sequences of the H. francisci vNAR clones studied in this experiment were aligned with corresponding domains from the wobbegong shark (Orectolobus maculatus, GenBank AF466396.1) and the nurse shark (Ginglymostoma cirratum, GenBank AY114863.1). Framework regions (FR1, FR2, and FR3) are shaded in gray; hypervariable regions (CDR1, HV2, HV4, and CDR3; Stanfield, 2007 #7) are in bold letters; canonical cysteines are shown in red; additional cysteines in blue.
Since the monoclonal antibodies were raised against isolated variable domains, we asked whether either the immune serum or one of the monoclonal antibodies was able to specifically recognize immunoglobulins in the context of whole shark serum. To test the recognition of antigen in its native conformation, we employed a dot blot analysis. The results (Fig. 3) confirmed that the immune serum and both monoclonal antibodies specifically reacted with molecules present in whole shark serum, but not in serum derived from other species such as llama or goat.

Monoclonal antibodies 370-12 and 533-10 specifically react with a protein present in shark serum. Sera from shark, llama, and goat were spotted onto a nitrocellulose membrane, dried, and detected with mouse sera (diluted 1:100) or hybridoma supernatants (diluted 1:10). Pre-immune serum (p.i.s. 3537), an immune serum from a mouse immunized against a llama VHH domain (i.s. VHH), and an irrelevant monoclonal antibody (1G4) were used as specificity controls.
MAb 370-12 recognizes a linear epitope in vNAR domains
In the previous experiments, MAb 370-12 had specifically reacted with all of the vNAR domains tested. It was therefore not possible to predict whether the epitope recognized was linear or conformational in nature. We thus tested whether MAb 370-12 would react with denatured vNAR domains present on a Western blot (Fig. 4). Indeed, MAb 370-12 reacted with all vNAR domains tested in a similar rank order of potency as had been observed previously in the ELISA assay, but did not react with an isolated heavy-chain V-domain from llama, demonstrating that this antibody was suitable for use in Western blot analyses.

Monoclonal antibody 370-12 reacts with vNAR molecules in Western blots. The indicated recombinant vNAR proteins and a VHH control were subjected to SDS-PAGE and visualized by staining with Coomassie Blue (upper gel) or immunodetection with MAb 370-12 (lower gel).
MAb 370-12 specifically precipitates IgNAR from H. francisci whole serum
The previous experiments had shown that MAb 370-12 recognized an epitope accessible both in linear and native conformations, and present in a broad spectrum of vNAR domains as well as in whole shark serum. It was unclear, however, if the antibody specifically reacted with IgNAR molecules, or whether it cross-reacted with other immunoglobulin classes. To resolve this question, immunoprecipitation experiments were performed. Diluted shark serum was passed through a column containing MAb 370-12 immobilized to a solid matrix, the column was washed, and bound proteins were eluted by pH shift. The resulting fractions were separated by SDS-PAGE and stained for total protein (Fig. 5A) or subjected to immunodetection by MAb 370-12 (Fig. 5B). MAb 370-12 specifically precipitated a minor protein of approximately 90 kDa (Fig. 5A, lane 5), corresponding to the expected molecular weight of IgNAR,(9) from whole shark serum. This band was also recognized by MAb 370-12 upon immunodetection (Fig. 5B, lane 5). To determine their identity, the two major bands observed in whole shark serum and the band precipitated by immobilized MAb 370-12 were excised from the gel, digested with trypsin, and analyzed by mass spectrometry. The analysis showed that the two major bands present in whole shark serum (Fig. 5A, lanes 1 and 2, arrows) corresponded to the heavy and light chains of conventional immunoglobulins from Heterodontus francisci. The band specifically precipitated by MAb 370-12 could be identified as IgNAR. Three peptides derived from this protein could be assigned to shark immunoglobulin sequences. One (DLTVEDSVTYYCK) perfectly matched a peptide contained in the third framework region of a previously submitted vNAR domain sequence from H. francisci (GenBank AAX10139), while two others (NEGVQLEAATK and SNQSGFTTTSPTK) showed significant homology to sequences NEGVQMEPTK from the C2 and TITSGFATTSPVK from the C1 constant regions of IgNAR of the nurse shark G. cirratum (GenBank AAB42621, Fig. 6).
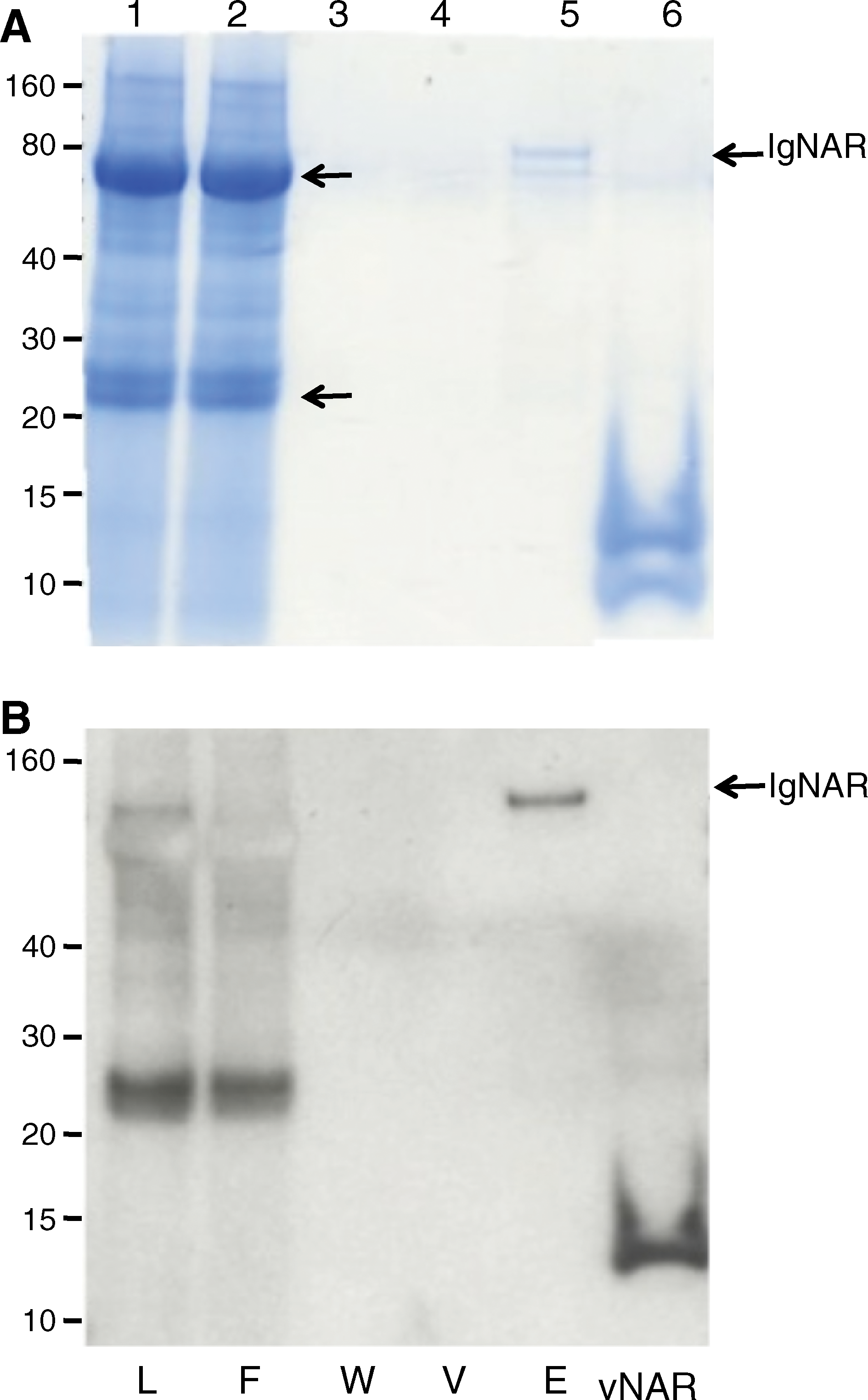
Monoclonal antibody 370-12 specifically recognizes IgNAR immunoglobulin in H. francisci serum. Serum from a H. francisci shark (diluted 1:10 in PBS) was passed through a column containing MAb 370-12 covalently coupled to an AminoLink matrix. Aliquots of the loaded serum (L), flow-through (F), wash (W), void (V), and eluted (E) fractions were subjected to SDS-PAGE (

Peptides identified in IgNAR immunoprecipitated from whole shark serum by MAb 370-12. The band marked as IgNAR in Figure 5 was excised and subjected to trypsin digestion and tandem MS/MS mass spectrometry. The location of the identified peptides (or their homologues) in IgNAR sequences from H. francisci and G. cirratum deposited in GenBank is shown.
Time course of IgNAR-mediated immune response of Heterodontus francisci
A major advantage of an antibody that recognizes IgNAR immunoglobulin is that it makes it possible to follow the specific IgNAR immune response of a horn shark during the course of an immunization, thereby permitting the optimization of immunization protocols and facilitating basic research on the immune response in these sharks. We thus used MAb 370-12 to monitor the immune response of two sharks immunized with a synthetic peptide derived from a conotoxin, and with human erythrocytes, respectively

Monitoring the IgNAR-mediated immune response of the horn shark Heterodontus francisci. Serum samples from two H. francisci sharks immunized with human erythrocytes (circles) or a synthetic conotoxin (squares) were drawn at different time points and probed for the presence of antigen-specific IgNAR using MAb 370-12. ELISA plates were coated with specific antigen (human glycophorin or conotoxin) or non-related targets (milk proteins or BSA) as controls. Serum samples were diluted 1:800 for analysis. Arrows indicate booster immunizations.
Discussion
In this report, we describe the generation of two monoclonal antibodies (370-12 and 533-10) raised against vNAR domains of the horn shark H. francisci. Although a single vNAR domain was used for immunization, the immunized mouse generated antibodies that recognize a broad spectrum of different variable domains. Both monoclonal antibodies derived from this immunization recognize epitopes common to several distinct vNAR domains. While MAb 533-10 recognizes a conformational epitope present in a subgroup of the vNAR domains tested in this study, MAb 370-12 reacts with a linear epitope accessible in the native conformation and present in all of the vNAR domains tested. Interestingly, MAb 370-12 specifically recognizes the IgNAR “heavy-chain-only” class of immunoglobulins in shark serum, and does not significantly cross-react with conventional immunoglobulins that are present in shark serum in much higher concentrations. Although vNAR domains are at the focus of biotechnological research because of their unique properties as single-domain antibodies, IgNAR constitutes only a minor fraction of the total immunoglobulins present in shark serum.(9) This is confirmed by our immunoprecipitation studies, which showed that MAb 370-12 specifically precipitated a minor band running close to, but not identical with, the major bands representing conventional immunoglobulin heavy chains. This precipitated protein was unequivocally identified as IgNAR by tandem MS/MS mass spectrometry.
Monoclonal antibodies have been raised previously against IgNAR molecules from the nurse shark(12) and used to follow the IgNAR immune response in these sharks.(9) Here we show that MAb 370 represents a valuable tool for studying the IgNAR response in H. francisi, a species belonging to a different order. It will be interesting to determine whether the epitope recognized by MAb 370-12 is also present in IgNAR molecules of sharks of other species.
Footnotes
Acknowledgments
This work was supported by grants from CICESE, and by a stipend from the National Council of Science and Technology and the Deutsche Akademische Austauschdienst (KJ).
Author Disclosure Statement
The authors have no financial interests to disclose.