
Abstract
Human inducible co-stimulator ligand (GL50, CD275), also known as B7-H2 or ICOSL, is a positive co-stimulatory molecule of B7/CD28 superfamily, which plays a critical role in immune response. Here we generated two novel mouse anti-human GL50 monoclonal antibodies (MAbs) using classical hybridoma technology. The two MAbs (clones 2B4 and 4D11) were IgG1 (κ) and IgG2a (κ), respectively, and bound specifically to human GL50. Epitope competition assay showed that 2B4 and 4D11 bind to the same epitope of GL50, which is not recognized by the commercially available GL50 MAb (9F-8A4). Functionally, the two MAbs act as a blocker of T cell proliferation. Taken together, as useful tools, these two antibodies might be of great value for further exploration of the immune identification and function of GL50.
Introduction
The B7/CD28 superfamily provides both positive and negative co-signals to immunocytes in immune responses.(8) Mounting evidence has shown that GL50, which is extensively expressed in both non-lymphatic and lymphatic tissues, is an important co-stimulatory molecule in up-regulating and promoting T cell immune responses.(2,4,9–12) Expression of GL50 in naive B cells and monocytes in PBMCs is at a low level. After stimulation by IFN-γ, TNF-α, or LPS, it can be quickly up-regulated.(2,12)
The induced expression of ICOS on activated T cells mainly regulates the secretion of Th2 cytokines and thus shifts the immune response to the Th2 type.(13) It has been reported that the ICOS/GL50 pathway is involved in immunopathogenesis such as infection, hypersensitivity, autoimmune disease (AID), transplantation immunity, and tumor immunity.(3,9,14,15,17,19,20) ICOS-/- mouse has an attenuated antibody response to collagen-induced CIA.(15) Alternatively, ICOS fusion protein significantly prolonged survival of allogeneic cardiac graft.(16) Furthermore, blocking of ICOS/GL50 signal by specific anti-GL50 MAb effectively relieved the inflammatory response in arthritis.(18) The significant inhibitory function of anti-GL50 MAb makes it a promising candidate in immunotherapy although the precise mechanism is still unknown.
In this study we successfully generated two mouse anti-human GL50 monoclonal antibodies (MAbs) (clones 2B4 and 4D11) and their immune characteristics were identified. Both MAbs bind to the same epitope, which is not recognized by commercially available MAb (9F-8A4). Function analysis shows that the two MAbs act as blockers of T cell proliferation. Therefore, the two novel MAbs promote better understanding of GL50-ICOSL signaling in immunoregulation. On the other hand, they are good candidates for clinical application.
Materials and Methods
Animals, cell preparation, reagents, and antibodies
Six-week-old BALB/c mice were purchased from the Department of Experimental Animal, Shanghai Institute of Biological Products, Ministry of Health of China. Murine myeloma cell line SP2/0, murine fibrous cell line L929, and all the human lines (Raji, Daudi, THP-1, HO8910, ECV, Jurkat) were originally obtained from American Type Culture Collection (Manassas, VA). Human peripheral blood mononuclear cells (PBMCs) were prepared from healthy donors by Ficoll-Hypaque (Hengxin Chemistry Company, Shanghai, China) density-gradient centrifugation. The T cells were enriched from PBMCs by EasySep and the purity was >90%. The above cells were cultured in RPMI 1640 medium (Gibco BRL, Grand Island, NY) or standard DMEM (Life Technologies, Grand Island, NY) supplemented with 10% FCS (Hyclone, Logan, UT), 100 U/mL penicillin, 100 μg/mL streptomycin, and 2 mM L-glutamine. Cells were cultured in a 5% CO2, 37°C incubator. Cell counting kit-8 (CCK-8) was purchased from Dojindo (Kumamoto, Japan). Commercial recombinant GL50-Ig protein was obtained from R&D Systems (Minneapolis, MN). Available PE-conjugated mouse anti-human GL50 MAb (clone 9F-8A4) was purchased from eBioscience (Woburn, MA). PE-conjugated goat anti-mouse IgG, biotinylated mouse IgG1, agonistic mouse anti-human CD3 MAb (clone UCHT1), and mouse IgG1 (clone 679.1Mc7) were all purchased from Immunotech (Marseille, France). Functional mouse anti-human CD28 MAb was prepared in our laboratory.(13)
Construction of transfectants
The complete cDNA encoding human GL50 was cloned from the human tumor cell line Daudi by reverse transcription polymerase chain reaction (RT-PCR) method with special primers (forward: 5′-CG
Generation of anti-GL50 specific monoclonal antibodies
Using classical hybridoma technology, the MAbs were prepared. Six-week-old BALB/c mice were immunized a total of four times with injections of 1 × 107 mitomycin-treated L929/GL50 cells in 0.5 mL phosphate-buffered saline (PBS) per mouse at 21-day intervals. The first subcutaneous injection was accompanied by complete Freund's adjuvant (Sigma-Aldrich, St Louis, MO). Four days after final boost injection, the splenocytes of immunized mice were fused with murine myeloma SP2/0 cells according to the method described by Groth and Zhang.(13) After hybridomas were selected by hypoxanthine-aminopterin-thymidine (HAT, Sigma-Aldrich), flow cytometry (Altra, Beckman Coulter, Miami, FL) was performed to screen positive clones. Hybridomas secreting mouse anti-human GL50 MAbs were obtained by their strong reactivity with L929/GL50 cells, but not with L929/mock, L929/CD28, or L929/ICOS transfectants. The hybridomas were injected intraperitoneally into pristane-primed BALB/c mice for 8–10 days. The MAbs were purified from ascites of a BALB/c mouse using protein G-sepharose affinity columns (Pharmacia, Uppsala, Sweden).
Characterization of anti-GL50 MAbs
For flow cytometric analysis, the transfected cells (1 × 106) mentioned above were incubated with MAb 2B4 or 4D11 for 30 min at 4°C and washed with PBS. For indirect staining, PE-conjugated goat anti-mouse antibody was added and incubated for another 30 min. After being washed with PBS, the stained cells were analyzed by flow cytometry and the Beckman-Coulter's Expo32 Multicomp software. The Ig isotypes were identified with the rapid test paper (Roche, Mannheim, Germany) according to the method provided by the manufacturer. The competition assay was performed by MAbs 2B4 and 4D11, respectively, with commercially available anti-GL50 MAb 9F-8A4 to determine their GL50 binding epitopes through mutual competition. The L929/GL50 cells (1 × 106) were incubated with 2B4 or 4D11 (0, 0.5, 1, and 2 μg/10 μL) for 45 min at 4°C. After being washed with PBS, the cells were incubated with 1 μg biotinylated identical MAb followed by streptavidin-PE for 30 min for flow cytometry to ascertain the saturating amounts of each MAb. For the epitope competition assay, saturating amounts of unlabeled 2B4 or 4D11 was incubated with L929/GL50 cells (5 × 105 in final volume of 50 μL) at 4°C for 30 min. Without washing, biotinylated 2B4 or 4D11 or PE-conjugated 9F-8A4 were added at 1 μg/test and incubation at 4°C proceeded for another 30 min. After being washed with PBS, cells stained with biotinylated 2B4 or 4D11 were further incubated with streptavidin-PE at 4°C for 30 min for flow cytometry.
Flow cytometry analysis
For flow cytometry analysis, the cells mentioned above (1 × 106/mL) were incubated with the MAbs 2B4 and 4D11, respectively, for 30 min at 4°C and washed. For indirect staining, PE-labeled goat anti-mouse antibody as secondary antibody was incubated for another 30 min at 4°C and washed. The results were analyzed by flow cytometry facility and the Beckman-Coulter's Expo 32 Multicomp software.
Dot-blot and Western blot analysis
The isotypes of the two MAbs were identified using Mouse Monoclonal Antibody Isotyping Kit (Roche) according to the manufacturer's instructions. Next, dot-blot and Western blotting assay were used to analyze the binding ability of the two MAbs to recombinant GL50-Ig. Briefly, drops of purified GL50-Ig were spotted onto circled dot (5 mm in diameter) on nitrocellulose membranes (Bio-Rad, Hercules, CA). After 30 min of drying at room temperature (RT), the membrane was blocked for 2 h at RT with 5% non-fat dry milk in PBS (pH 7.2) on a rocking platform. The membrane was then washed three times with PBS containing 0.1% Tween-20 (PBST), and incubated with PBS, 5 μg/mL anti-GL50 MAbs (9F-8A4, 2B4, or 4D11) or mouse IgG1 for 2 h at RT. After being washed, the membrane was sealed in 5% milk containing 1:2000 dilution of horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG and incubated at RT for 1 h with agitation. The membrane was subjected to an enhanced chemiluminescence (ECL) kit (Roche) and then exposed to film. Western blot analysis was performed to analyze the binding capacity of 2B4 and 4D11 MAbs to purified GL50-Ig and GL50 protein on L929/GL50 and Daudi. L929/mock cells were set as negative control. 10 μg purified GL50-Ig and the cell lysates of L929/GL50, L929/mock, and Daudi were mixed with loading buffer and boiled at 95 − 100°C for 5 min, fractionated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (10%), and transferred onto the nitrocellulose filter, which was treated as described above for dot-blot.
Enzyme-linked immunosorbent assay of MAbs
The application of the obtained MAbs was determined by indirect ELISA. Briefly, the recombinant GL50-Ig fusion protein was diluted serially two-fold to different concentrations (200, 100, 50, and 25 ng/mL) with carbonate buffer (pH 9.6) and pre-coated on a 96-well ELISA plate at 4°C overnight. After being washed three times with PBS, 2% bovine serum albumin (BSA) was added to the wells for blocking at room temperature to avoid non-specific binding. Then the obtained anti-GL50 MAbs (1 μg/mL) were added for 1 h at room temperature, followed by HRP-goat anti-mouse IgG at 1:1000 for 1 h at 37°C. The absorbance of the wells was revealed with the substrate 3, 3, 5, 5-tetramethylbenzidine (TMB, Sigma). The reaction was stopped with 2 M H2SO4, and the plate was measured at 450 nm by a microplate reader (Bio-Rad). In this experiment, the plate was washed five times with PBS containing 0.05% Tween-20 after each of these steps.
Analysis of T cell proliferation
Purified human T cells (1 × 105/well) from peripheral blood mononuclear cells (PBMCs) were seeded into 96-well plates pre-coated with anti-CD3 MAb (0.5 μg/mL), anti-CD3 MAb (0.5 μg/mL) + L929/mock (1 × 104/well), and anti-CD3 MAb (0.5 μg/mL) + L929/GL50 (1 × 104/well). Meanwhile, different concentrations of the anti-GL50 MAbs and soluble anti-CD28 MAb were added to the medium and cultured at 37°C, 5% CO2. Mouse IgG1 was used as control in this experiment. After 3 days in culture, the reagent of cell counting kit-8 (CCK-8, Dojindo, Kumamoto, Japan) was added to the culture system and incubated for 6 h. The absorbance of every well was measured at 450 nm by a microplate reader (Bio-Rad). The procedure was performed as described previously.(26) The Student's t test was applied for statistical analysis among the groups.
Results
Establishment of two novel anti-human GL50 MAbs
As shown in Figure 1A, the L929/GL50 transfectants expressed on their membrane high amounts of target molecules in comparison with L929/mock cells. To generate MAbs specific for human GL50, the splenocytes of mice immunized by L929/GL50 cells were fused with myeloma SP2/0 cells. After repeated screening and cloning, two MAbs, named 2B4 and 4D11, were obtained. Immunophenotyping showed that 2B4 and 4D11 could recognize transfected cells L929/GL50, but not L929/mock, L929/CD28, and L929/ICOS cells, indicating that 2B4 and 4D11 are specific for GL50 (Fig. 1B). The isotype of 2B4 and 4D11 were IgG1 (κ) and IgG2a (κ), respectively.

(
For further analysis, we conducted dot-blot and Western blotting to confirm the specificity of the two MAbs and explore whether structural folding is indispensable for their recognition of GL50. As shown in Figure 2A, just as with 9F-8A4, both MAbs were proven in dot-blot to specifically bind to natural GL50-Ig compared with PBS and mouse IgG1 staining (negative control). However, neither of them could recognize the denatured GL50-Ig or GL50 from L929/GL50 and Daudi cell lysates (data not shown). The MAbs were also identified by ELISA. The results indicated that both 2B4 and 4d11 can bind to immobilized GL50 Ig protein in indirect ELISA (Fig. 2B). Thus, the two novel MAbs against human GL50 reported in this work provide additional reagents for the functional characterization of GL50.
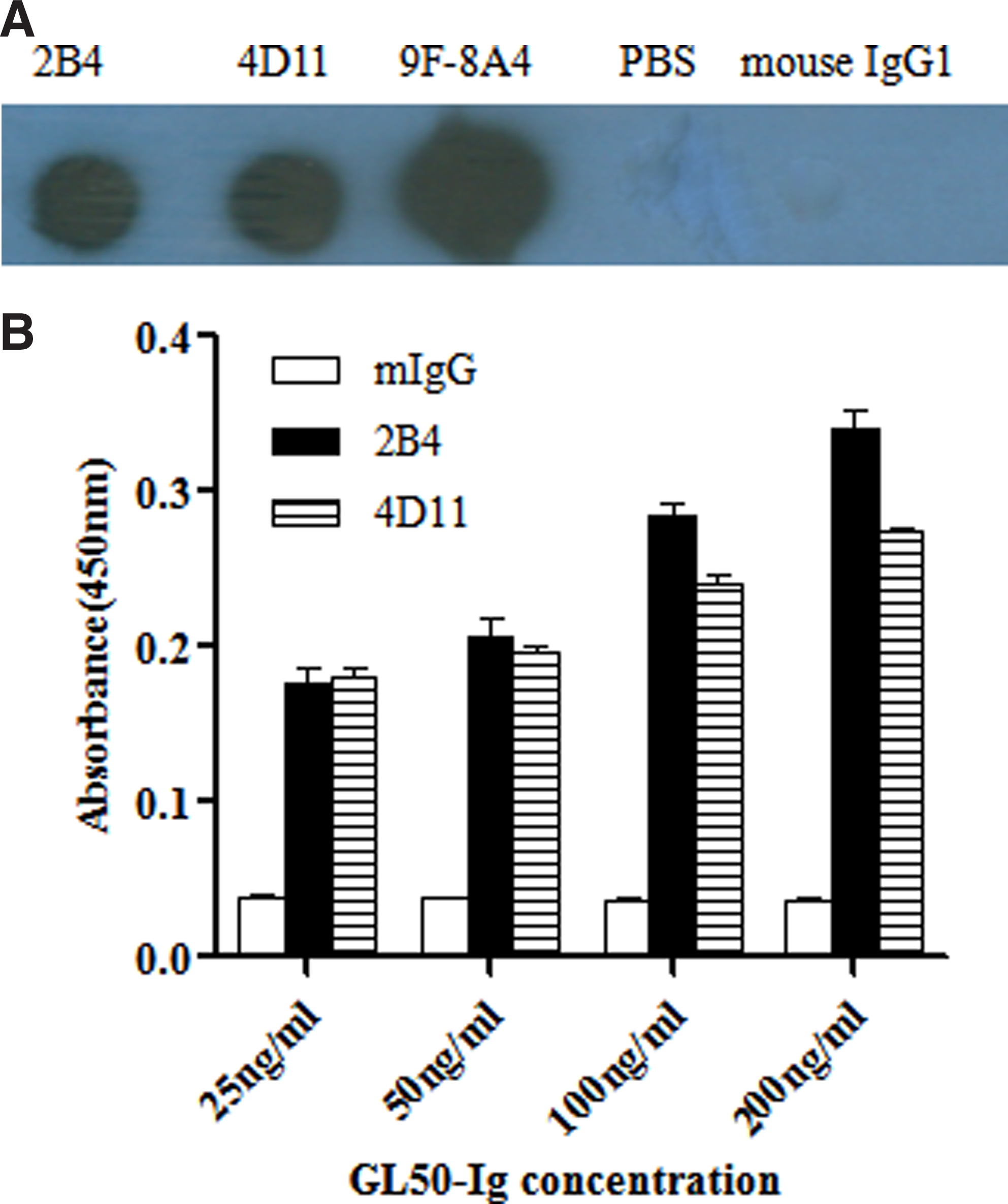
(
Epitope recognized by 2B4 and 4D11 is different from that of commercially available 9F-8A4 MAb
Competition assay was performed to further determine whether the two antibodies recognize the same, overlapping, or distinct epitopes of human GL50 antigen. At first, the L929/GL50 cells (1 × 106) were incubated with 2B4 or 4D11 (0, 0.5, 1, and 2 μg/10 μL) for 45 min at 4°C. After being washed with PBS, the cells were stained with the detecting amounts (1 μg/test) of biotin-conjugated identical antibody to ascertain the saturating concentration at which all of the binding sites on L929/GL50 were occupied by this MAb. As shown in Figure 3A, graded amounts of unlabeled 2B4 or 4D11 blocked subsequent binding of biotinylated MAb to L929/GL50 cells in a dose-dependent way and 2 μg was sufficient for both antibodies to saturate their interacting sites in GL50, respectively. Secondly, mutual competition assay between 2B4 and 4D11 and commercial 9F-8A4 was performed using the same protocol. The results showed that the incubation of saturating amounts of unlabeled 2B4 MAb as a competitor could inhibit the following binding of biotin-conjugated 4D11 to L929/GL50 cells. Conversely, pre-staining of L929/GL50 transfectants with unlabeled 4D11 as a competitor also inhibited the subsequent recognition of commercial 9F-8A4. However, binding of PE-conjugated 9F-8A4 could not be blocked in 2B4 or 4D11 saturated L929/GL50 cells (Fig. 3B). Taken together, these data indicated that these two MAbs (2B4 and 4D11) did not compete with PE-conjugated 9F-8A4 in binding to human GL50 while 2B4 and 4D11 seemed to recognize the same epitope.
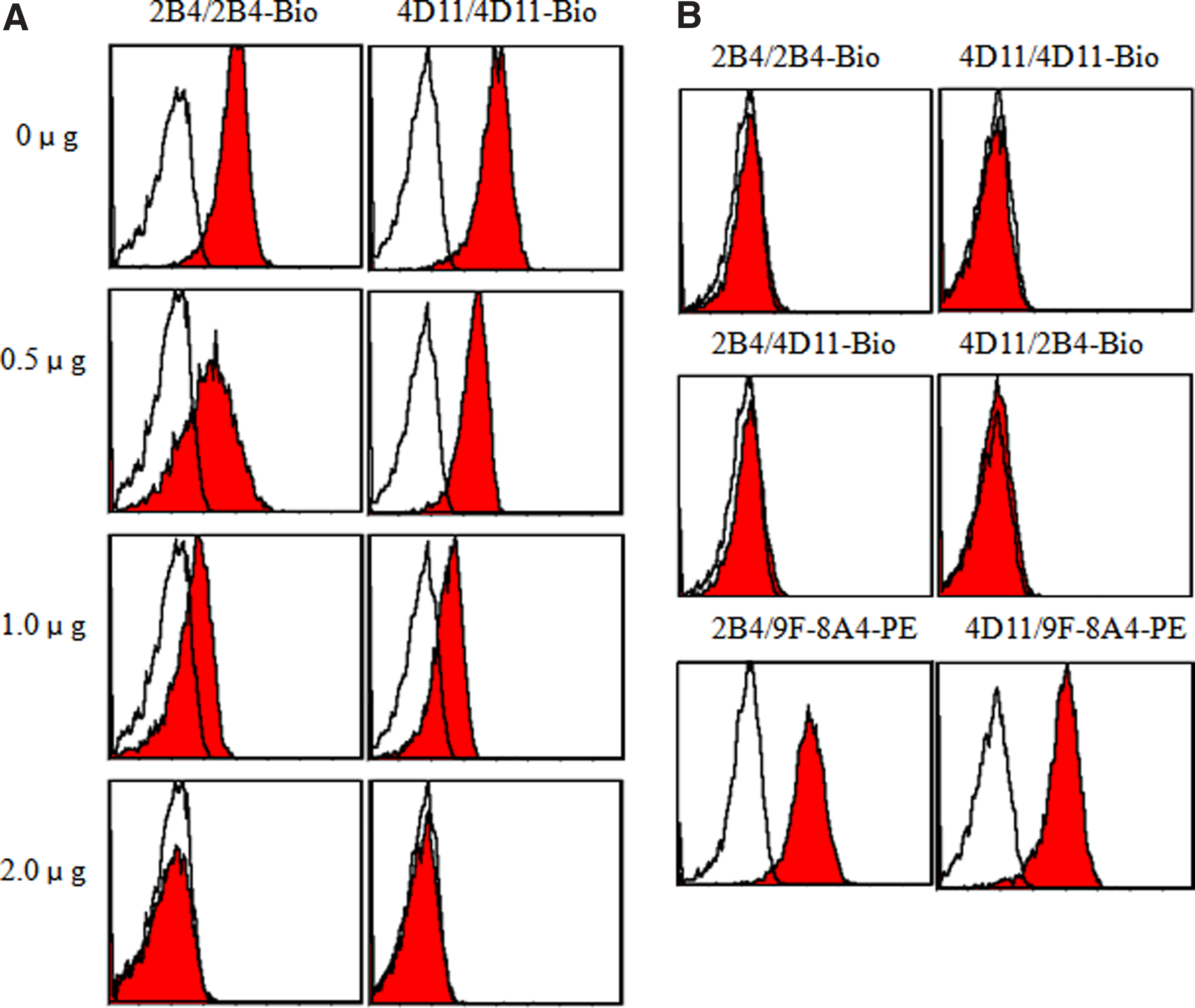
Competitive inhibition test. (
Expression of GL50 on different cell lines
The expression of GL50 was examined on different human tumor cell lines, using flow cytometry (Fig. 4). Results showed that GL50 could be highly expressed on tumor cell lines of Raji, Daudi, THP-1, HO8910, ECV, but not expressed on Jurkat and the two MAbs we generated could be used in FCM analysis. With the expression of GL50 in these tumor cell lines, detected by these two new MAbs 2B4 and 4D11, these two MAbs may have great value for further study of tumor stem cells.

Expression of GL50 antigen on different cell lines was determined by FCM with MAbs 2B4 and 4D11 and commercial MAb 9F-8A4. Different human cell lines were stained with MAb (2B4, 4D11, or 9F-8A4) (grey histograms) or negative control mouse IgG (open histograms) at 4°C for 30 min, followed by goat anti-mouse IgG-phycoerythrin. The figure is a representative of three experiments.
Both 2B4 and 4D11 could inhibit T cell proliferation
To assess whether the blocking MAbs 2B4 and 4D11 exhibited any functional effect on T cells, an in vitro direct proliferative assay was performed in the presence of MAbs 2B4 or 4D11 and L929/mock and L929/GL50. Compared with the control groups, the addition of MAb 2B4 or 4D11 inhibited T cell proliferation in a dose-dependent manner that could be measured by CCK-8 incorporation within 72 h of incubation (Fig. 5). Therefore, MAbs 2B4 and 4D11 are functional antibodies that can inhibit ICOS/GL50 positive signals on T cells.
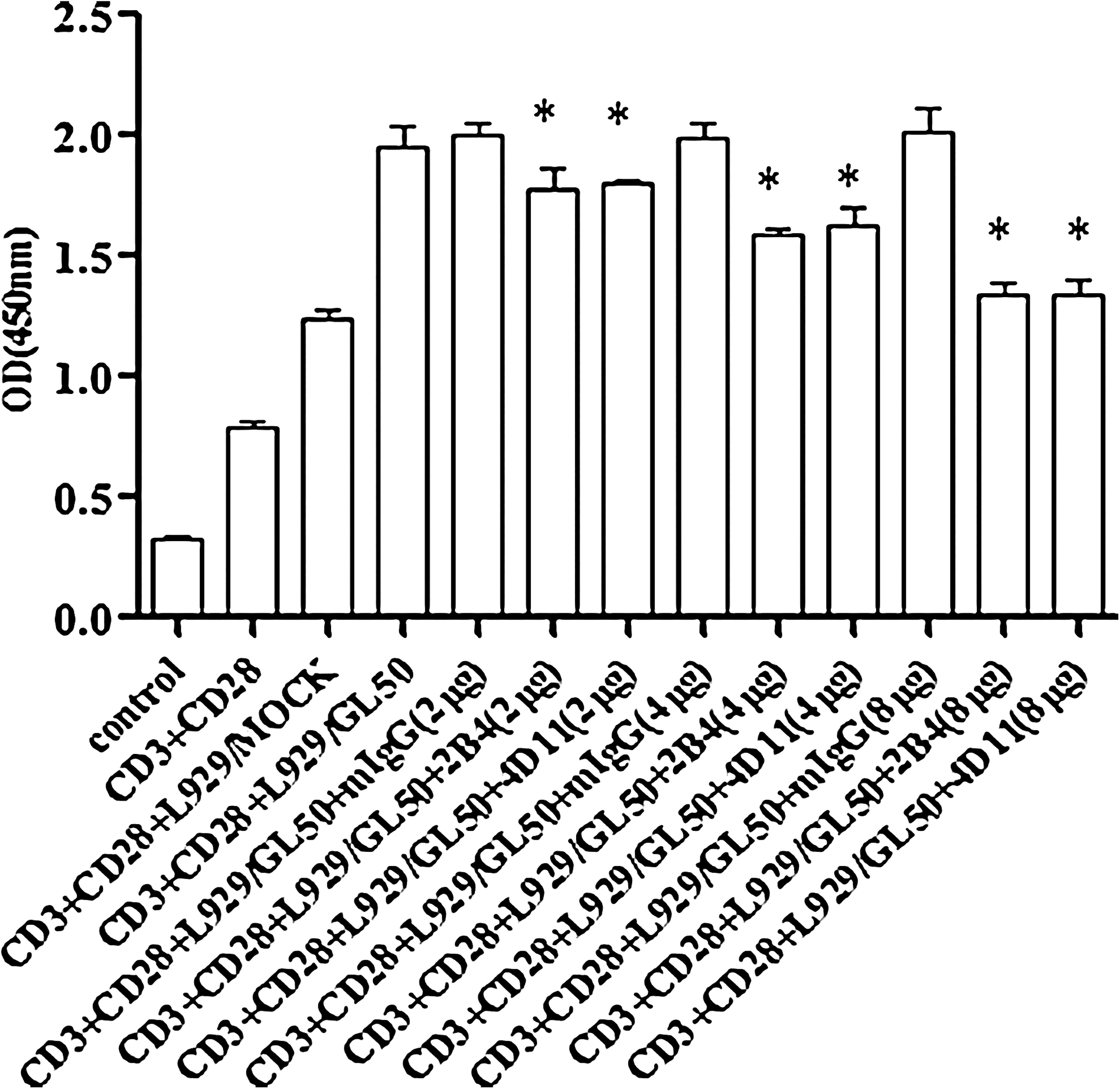
T cell proliferation upon engagement of GL50 with MAbs 2B4 and 4D11. Blockade of ICOS/GL50 positive pathway by 2B4 and 4D11. MAbs inhibit T cell proliferation. T cells were seeded to the 96-well plates pre-coated with anti-CD3 MAb (0.5 μg/mL). Meanwhile, different concentrations (2, 4, or 8 μg/mL) of 2B4 and 4D11 MAbs, anti-CD28 MAb, or mIgG1 isotype control were added to the culture systems for 72 h. T cell proliferation was determined by the CCK-8 reagent according to the protocol. The results are shown as the mean ± SD of triplicate wells (*p < 0.05 compared to the control groups). All these experiments used isotype-matched mouse IgG1 as control. The figure is representative of three experiments.
Discussion
GL50 is an identified member of the B7 superfamily.(7) The interaction of GL50 with ICOS, the specific receptor for GL50, is critically involved in the activation, proliferation, differentiation, and cytokine production of T cells as well as in the antibody secretion from B cells during secondary immune responses.(2,6,9,10,22,27) It is well-accepted that the ICOS/GL50 signal pathway plays a pivotal role in the regulation of inflammatory responses, in the prevention of transplantation rejection and tumor development, as well as in the modulation of autoimmune diseases.(9,10,14,15,20,28)
The fluctuant balance between co-stimulatory and co-inhibitory signals that a T cell receives participates in the initiation, effection, and termination of an immune response.(8,21) Excessive activation and immune reaction of T cells may result in autoimmune diseases and host immune injury. GL50 delivers a potent co-stimulatory signal to T cells when engaged by ICOS, resulting in T cell activation and proliferation.(2,9,10) The existence of ICOS/GL50 signal in vivo is closely associated with many mouse autoimmune disease models. Conversely, the absence of ICOS/GL50 signal may be a good way to relieve autoimmune disease.(19,24,25) In view of its critical function in regulating immunohomeostasis, ICOS signaling has aroused great attention in immunodiagnosis and therapy.
Undoubtedly, the development of more efficient and specific monoclonal antibodies may be important for further disclosure of GL50 function. The choice of appropriate immunization strategy and screening assay is the most important part of hybridoma production. Here, in order to avoid an immune response against heterogeneous antigen, mouse original fibrous cell line L929 was selected to carry the target antigen. Human GL50 antigen presented by the L929 cells can produce a more specific immune response and decrease the probability of non-specific MAb generation. All the hybridomas were screened by flow cytometry, whose constraints on screening of assays are reliable, fast, and simple. According to these considerations, we successfully obtained two desired hybridomas secreting mouse anti-human GL50 MAbs by one fusion operation.
After purification, the two novel mouse MAbs generated against human GL50 were identified and subjected to strict verification by dot-blot and Western blot analysis. Both 2B4 and 4D11 specifically bound to natural GL50-Ig in dot-blot while in conditioned Western blotting the denatured GL50 protein could not be immunoblotted. This indicated that the two MAbs could not bind to a linear epitope. However, they seemed to recognize the same epitope of GL50, which is different from commercially available 9F-8A4.
Additional experimental evidence showed that the reactivity of the two MAbs in a capture ELISA with the pre-coated GL50-Ig fusion protein was markedly increased in a dose-dependent manner. Moreover, the MAbs have the ability to prevent the binding of the GL50 to ICOS. Engagement of human GL50 by the two MAbs could inhibit T cell proliferation, suggesting that these two MAbs may be useful for blocking the ICOS/GL50 positive signal.
With their high specificity and few side effects, monoclonal antibodies have recently been unanimously thought to be ideal medicaments in targeting therapy for some refractory diseases such as malignant tumor.(28) With the expression of GL50 on the tumor cell lines above, detected by two new MAbs 2B4 and 4D11, these MAbs may have great value for the further study of tumor stem cells. Therefore, 2B4 and 4D11 have potential value in the further study of tumor escape and autoimmune diseases.
In conclusion, we have prepared two specific mouse MAbs against human GL50 that recognize the same epitope, which is different from the commercially available 9F-8A4. They act as an adjuvant to inhibit the proliferation on purified human T cells. This makes MAb 2B4 and 4D11 valuable tools for further exploration of GL50 function. Furthermore, another anti-GL50 MAb will be generated so that a sandwich ELISA technology can be developed based on these MAbs to analyze GL50 in view of its important regulatory functions.
Footnotes
Acknowledgments
This work was supported by a grant from the National Natural Science Foundation of China (grant no. 30801023).
Author Disclosure Statement
The authors have no financial conflict of interest to disclose. The generated antibodies were distributed solely to non-profit research organizations for research purposes only.