
Abstract
The heat shock protein 70 (Hsp70) family members function as ATP-dependent molecular chaperones that assist in the folding of newly synthesized polypeptides and in the refolding of misfolded/aggregated proteins. These heat shock proteins comprise at least eight sets of molecular groups that share high homology, but differ from each other in their expression level and subcellular localization. Hsp72, which is also known as Hsp70 and Hsp70-1, is localized mainly in the cytoplasm but is also found in the nucleus. Stress-induced Hsp72 functions as a chaperone enabling the cells to cope with harmful aggregations of denatured proteins during and following stress. The difference in the function of Hsp72 from that of other Hsp70 members, however, remains unclear. We report the establishment of a monoclonal antibody specific for Hsp72 using the rat medial iliac lymph node method. Immunoblot analysis revealed that our monoclonal antibody against Hsp72 specifically identified the 65 kDa protein. Immunocytochemical staining also revealed that Hsp72 localized in the cytoplasm and nucleus, and aggregated in the nucleus in response to heat stress. This MAb against Hsp72 will allow for further studies to elucidate the mechanism by which Hsp72 is localized in the cell in response to stress stimuli, and aid in the identification of specific interacting molecules.
Introduction
Hsp72 and heat shock cognate protein 70 (Hsc70) are two major cytosolic/nuclear Hsp70s with extensive sequence homology (86%). Hsc70 is abundantly and ubiquitously expressed in all cells, whereas Hsp72 is stress-induced. Hsp72 is expressed only at low or undetectable levels in most unstressed normal cells and tissues, but its expression is rapidly induced by a variety of physical and chemical stressors, and it is therefore often called stress-inducible Hsp70. Stress-induced Hsp72 functions as a chaperone enabling the cell to cope with harmful aggregations of denatured proteins during and following stress.(3–5) In response to heat stress, the chaperone function of Hsp72 is required for its cytoprotective effects, and Hsp72 is suggested to inhibit the accumulation of protein aggregates and thereby remove the stimulus that triggers cell death.(6,7) Hsc70 also has chaperone functions, including the folding of nascent polypeptides and the prevention of protein aggregation under stress conditions.(5,8) Furthermore, the intracellular localization of Hsp72 and Hsc70 is very similar.(9)
Previous studies in HeLa cells using indirect immunofluorescence demonstrated that Hsp72 and Hsp73 are present in both the cytosol and nucleus in resting cells and become predominantly located in the nucleus during and soon after heat stress.(10,11) With time after stress, these proteins return to the cytosol. It is unclear why two isoforms are expressed and whether these two proteins have different functions in the cell. The functional specificity of the two cytosolic isoforms, Hsp72 and Hsc70, has not yet been resolved. Elucidation of the difference in Hsp70 and Hsc70 would be useful for functional analysis. One effective method of elucidating the function of Hsp72 would be to determine the changes in stimulus-dependent Hsp72 binding proteins and the localization of Hsp72 in detail. We previously reported an Hsc70-specific antibody, 6E1.(12)
In this report, we established a monoclonal antibody specific for Hsp72 to elucidate the mechanism by which Hsp72 localizes in the cell against stress stimuli and to identify specific interacting molecules.
Materials and Methods
Cell culture
HeLa cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, penicillin (100 U/mL), and streptomycin (100 mg/mL; Nacalai Tesque, Kyoto, Japan) under a humidified atmosphere with 5% CO2 at 37°C.
Production and purification of recombinant proteins
Expression vectors of glutathione S-transferase (GST) fusion proteins to full-length human Hsp72 were transformed into Escherichia coli BL21(DE3) (Novagen, Madison, WI).(13) The E. coli cells were grown in MMI medium containing 50 mg/mL carbenicillin (Nacalai Tesque) at 37°C. Expression was induced by the addition of 0.5 mM isopropyl-1-thio-b-D-galactopyranoside (Nacalai Tesque), followed by incubation at 37°C for 12 h. Bacteria were lysed and fusion proteins purified according to previously described methods.(12,14,15)
Immunization of rat and production of monoclonal antibodies
Anti-Hsp72 rat monoclonal antibodies (MAbs) were generated using the rat lymph node method established by Sado and colleagues.(14,16,17) A 10-week-old female lzm rat (Japan SLC, Shizuoka, Japan) was injected via the hind footpads with 500 mL of an emulsion containing 1000 mg of recombinant mouse Hsp72 protein and Freund's complete adjuvant. After 2 weeks, the cells from the lymph nodes of a rat immunized with the antigen were fused with mouse myeloma Sp2/0-Ag14 cells at a ratio of 5:1 in a 50% polyethylene glycol (PEG1500, Merck, Darmstadt, Germany) solution. The resulting hybridoma cells were plated onto 96-well plates and cultured in HAT selection medium (hybridoma SFM medium [Invitrogen, Carlsbad, CA]; 10% fetal bovine serum; 10% BM-condimed H1 [Roche, Basel, Switzerland]; 100 mM hypoxanthine; 0.4 mM aminopterin; 1.6 mM thymidine). At 7 day post-fusion, the hybridoma supernatants were screened using an enzyme-linked immunosorbent assay (ELISA) against GST-fused Hsp72. Positive clones were subcloned and rescreened by ELISA and immunoblotting. To prepare hybridoma supernatants containing highly concentrated antibodies, the resulting positive clone 3G7 was then cultured at a high density using a miniPERM bioreactor (Vivascience, Hannover, Germany).
ELISA
Hsp72 antigen (5 mg/mL) in TBS-T (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, and 0.05% Tween-20) was adsorbed on the surface of 96-well flexible microplates (Thermo Scientific, Rockford, IL) by overnight incubation at 4°C. To avoid non-specific binding, the plates were blocked with 1% bovine serum albumin (BSA, Nacalai Tesque) in TBS-T. Hybridoma supernatants were incubated for 1 h at room temperature, and then washed with TBS-T three times. The plates were incubated for 30 min at room temperature with alkaline phosphatase-conjugated anti-rat IgG antibody (Sigma, St. Louis, MO) diluted 1:10,000. After washing with TBS-T three times, immunoreactivity was visualized using a pNPP phosphatase substrate system (KPL, Gaithersburg, MD).(18)
Immunoblotting
HeLa cells were washed with phosphate-buffered saline (PBS) and lysed in TNE lysis buffer comprising 10 mM Tris-HCl [pH 7.4], 150 mM NaCl, 5 mM EDTA, 1% Triton X-100, 10% glycerol, 100 mM NaF, 0.2 mM dithiothreitol, 1 mM phenylmethylsulfonyl fluoride, and protease inhibitor cocktail (Nacalai Tesque). Lysates for immunoblotting (25 mg protein) were separated on 7.5% sodium dodecyl sulfate-polyacrylamide gels under reducing conditions, followed by electrophoretic transfer to polyvinylidine difluoride membranes (Immobilon-P, Millipore, Billerica, MA). The membranes were blocked for 1 h at room temperature with a blocking solution containing 5% skim milk in TBS-T (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, and 0.1% Tween-20), and then incubated for 1 h at room temperature with anti-Hsp72 rat MAb diluted in a 5% skim milk/TBS-T. After washing with TBS-T, the membranes were incubated for 1 h at room temperature with anti-rat IgG conjugated horseradish peroxidase (GE Healthcare, Buckinghamshire, United Kingdom). After washing with TBS-T, the membranes were exposed to LAS-3000 (Fujifilm) using the enhanced chemiluminescence technique (Immobilon Western HRP Substrate, Millipore).(19) Other antibodies were obtained from the following sources: anti-Hsc70 antibodies from StressMarq Bioscience (Victoria, Canada); anti-V5 antibody from Bethyl Laboratories (Montgomery, TX); and anti-β-actin antibody from Sigma.
Immunocytochemistry
HeLa cells were grown on coverslips and washed twice in PBS at 37°C and fixed with 1% paraformaldehyde for 5 min at room temperature. After rapid washing in PBS, cells were permeabilized at 37°C for 5 min in 0.5% Triton X-100 in PBS, then incubated in blocking buffer as described previously.(20) Hsp72 was detected with anti-Hsp72 rat MAb 3G7, followed by an Alexa 488-conjugated goat anti-rat IgG (Invitrogen) for 1 h at room temperature. Coverslips were mounted with Prolong (Molecular Probes, Carlsbad, CA). Hsp72-expressing cells were visualized under a fluorescence microscope (Olympus, Tokyo, Japan).
Result and Discussion
The human Hsp72 possesses a full-length 641-amino acid sequence. To establish a specific antibody against Hsp72, we utilized a full-length amino acid of human Hsp72 as the antigen. We then generated a monoclonal antibody specific for Hsp72 with GST-hHsp72 using the rat lymph node method. ELISA screening was performed and 34 positive identical clones, 3G7, were obtained from the hybridoma. The isotype of this clone was IgG2a/k.
Stress-protective heat-shock proteins are often overexpressed in cells of various cancers and are suggested to be contributing factors in tumorigenesis.(3,21,22) Therefore, we evaluated the binding ability of this antibody, 3G7, using HeLa cells. To evaluate the reactivity against Hsp72, immunoblot analysis was performed using 3G7. 3G7 reacted strongly with a 65 kDa band in the HeLa cell protein extract (Fig. 1A). Additional bands were observed at 110 and 45 kDa. To evaluate the affinity of 3G7 against Hsp72, we examined whether 3G7 could be used to observe Hsp72 expression induced by heat shock. HeLa cells were exposed to heat shock (42°C) for 1 h, and then the cells were cultured at 37°C for 3 or 12 h. 3G7 recognized heat-induced Hsp72 expression at 3 h. Expression of Hsp72 continued for 12 h (Fig. 1A). We also performed immunoprecipitation using 3G7. Immunoblot analysis was performed after immunoprecipitation and 3G7 recognized Hsp72 as a 65 kDa band (Fig. 1B). These findings suggest that 3G7 specifically identified Hsp72 and allows for effective affinity purification of proteins that bind Hsp72.
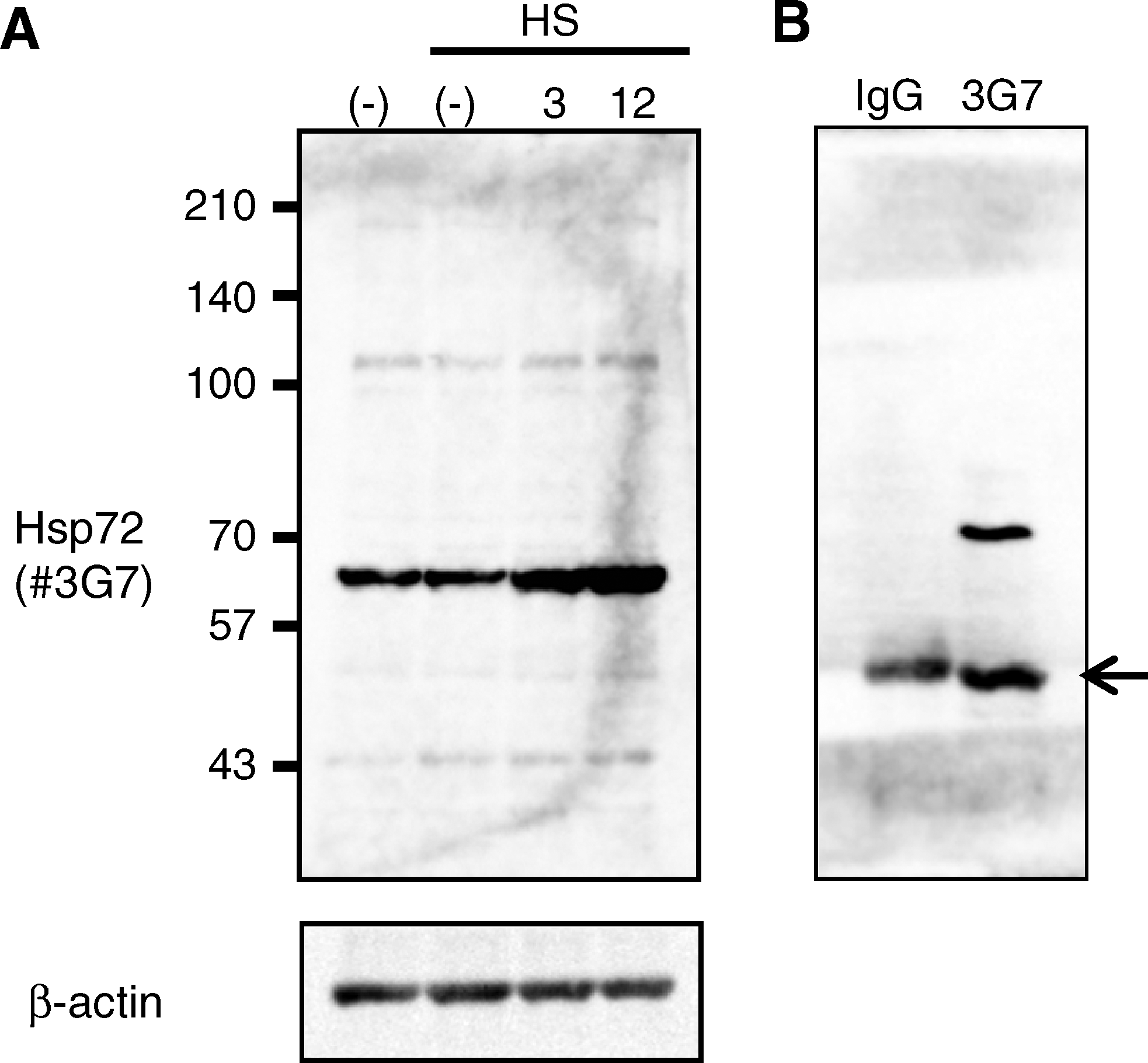
Expression of Hsp72 detected by MAb 3G7. The specificity of the antibody was examined. (
Hsp72 protein localizes in the cytosol and in the nucleus. Recent evidence indicates that Hsp72 protein is concentrated in the nuclei when cells are exposed to heat stress.(11) Therefore, we determined the subcellular localization of endogenous Hsp72 using 3G7 in heat-shocked HeLa cells (Fig. 2). HeLa cells were exposed to 1 h of heat shock, after which the cells were allowed to recover at a normal temperature for 3 or 12 h. Hsp72 protein was identified in the cytoplasm and in the nucleus under the unstressed condition, but Hsp72 rapidly translocated from the cytosol to the nucleus, particularly the nucleoli, following 1 h of heat shock. Three hours after heat stress, Hsc70 gradually re-translocated to the cytoplasm and returned to the basal distribution 12 h later. These data are consistent with previous reports.(10,23,24) Thus, immunostaining of Hsp72 with 3G7 showed a profound change in the distribution of Hsp72 depending on heat stress.
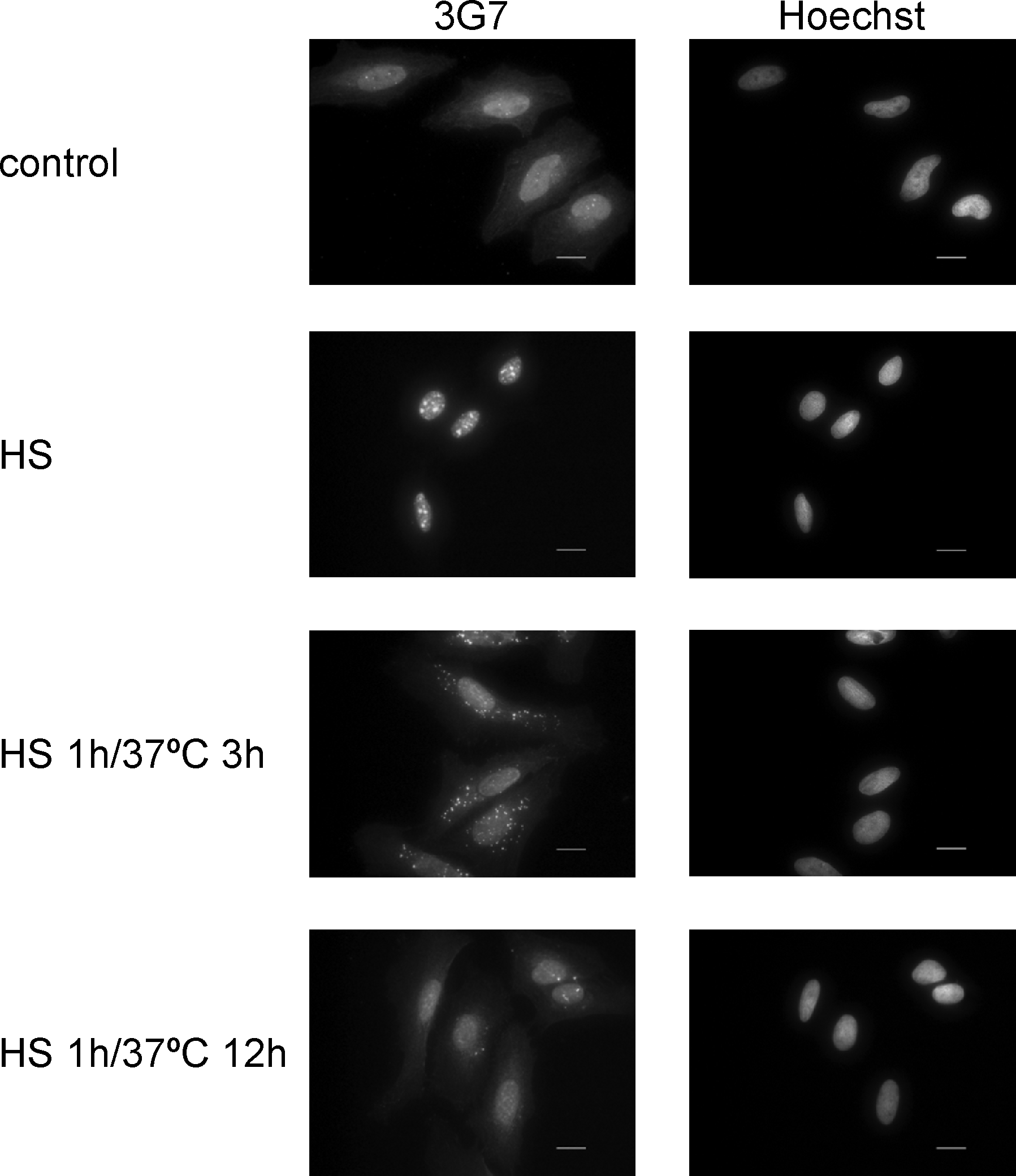
Subcellular localization of endogenous Hsp72 detected using 3G7 by immunocytochemistry. HeLa cells were untreated (control) or heat shocked at 42°C for 1 h (HS). After heat shock, cells were cultured at 37°C for 3 or 12 h (HS 1 h/37°C 3 h; HS 1 h/37°C 12 h). The cells were immunostained with the 3G7 clone. The antibody was detected with Alexa 488-conjugated anti-rat IgG. The sample was counterstained with Hoechst. Scale bar, 20 μm. HS, heat shock.
This monoclonal antibody 3G7 is specific for Hsp72 and will facilitate elucidation of the function of Hsp72 under stress.
Footnotes
Author Disclosure Statement
The authors have no financial interests to disclose.