
Abstract
To investigate cell surface antigens on human embryonic stem cells (hESCs), we generated a panel of monoclonal antibodies (MAbs) against undifferentiated hESCs by a decoy immunization strategy. One of the MAbs, MAb 2-E2, specifically bound to human pluripotent stem cells but not to mouse pluripotent stem cells and mouse embryonic fibroblasts. 2-E2 also bound to human differentiated cells, peripheral blood monocytes, and dermal fibroblasts. 2-E2 antigen expression drastically decreased in retinoic acid-induced differentiated hESCs. However, it gradually increased after initial decrease during embryoid body formation of hESCs. 2-E2 recognized approximately 68 and 27 kDa proteins present on the surface of human pluripotent stem cells. The results suggest that 2-E2-specific surface protein is a novel cell surface protein that plays a role in the early differentiation of human pluripotent stem cells.
Introduction
In order to study the effect of the external signal on hESC self-renewal and pluripotency and to prospectively isolate hESCs from the mixed cell population, it is necessary to find the specific surface markers for hESCs. Previously, we generated a panel of monoclonal antibodies (MAbs) specific to surface molecules of undifferentiated hESCs by using a decoy immunization strategy.(7) During further study of the isolated MAbs, 2-E2 was specifically shown to bind to human pluripotent stem cells, but not to mouse pluripotent stem cells. In addition, 2-E2 unexpectedly bound to human differentiated peripheral blood monocytes (PBMs). Therefore, the level of 2-E2 antigen was dissected during differentiation. The surface protein recognized by 2-E2 was also investigated.
Materials and Methods
Cell culture and antibody purification
hESC lines H1, H9, and CHA-hES4 were cultured on mouse embryonic fibroblast (MEF) feeder cells as described previously.(7) Differentiation of H9 cells was induced by incorporating all trans-retinoic acid (RA, Sigma-Aldrich, Seoul, Korea) at 10−5 M into the medium and culturing the cells for at least 21 days. Human embryoid body (EB) was prepared from H9 cell colonies as described previously.(8) Mouse embryonic stem cell (mESC) lines J1, R1, and E14Tg2a.4 were cultured as described previously.(7,9) Human peripheral blood mononuclear cells (PBMCs) were isolated by the Ficoll-Paque Plus method (GE Healthcare, Seoul, Korea). The human embryonal carcinoma cell lines NT-2/D1 and NCCIT were obtained from the ATCC (Manassas, VA) and maintained according to the protocol provided by the supplier. Human dermal fibroblast cells (DFCs) were purchased from MCTT (Seoul, Korea). Hybridoma 2-E2 has been described previously.(7) For antibody purification, female BALB/c mice were injected intraperitoneally with 5 × 106 2-E2 cells. Ascites was collected and clarified by centrifugation at 500 xg for 10 min. The ascitic fluid was then subjected to affinity chromatography on protein G-Sepharose column, and bound antibody was purified as described previously.(7)
Flow cytometry analysis
hESCs and mESCs were treated with collagenase IV (2 mg/mL) for 5 − 10 min in normal growth medium, treated with cell dissociation buffer (Invitrogen, Seoul, Korea) for 10 min in a 37°C incubator, and filtered through a 40 μm cell strainer as described previously.(7) The other cells were also treated with cell dissociation buffer as described above. The dissociated cells were incubated with antibodies in PBA (1% bovine serum albumin, 0.02% NaN3 in PBS) for 30 min at 4°C. Then, the cells were further incubated with fluorescein isothiocyanate (FITC)-conjugated anti-mouse immunoglobulin (Ig) G (BD Biosciences, Seoul, Korea). Propidium iodide (PI)-negative cells were analyzed for the antibody binding using FACSCalibur (BD Biosciences).
Cell-surface biotinylation and immunoprecipitation
Cell surface biotinylation of H9 or NT-2/D1 cells was performed as described previously.(7) Briefly, biotin-labeled H9 or NT-2/D1 cells were treated with lysis buffer (25 mM Tris-HCl [pH 7.5], 250 mM NaCl, 5 mM EDTA, 0.1% Nonidet P-40, 2 μg/mL aprotinin, 100 μg/mL PMSF, 5 μg/mL leupeptin) at 4°C for 30 min. Cell lysates were incubated with 2-E2 antibody and protein G-Sepharose beads as described previously.(7) Immunoprecipitated complexes were separated on a SDS-PAGE gel and transferred to PVDF membrane. The Western membrane was then incubated with streptavidin-horseradish peroxidase (SA-HRP, 1:4000; GE Healthcare) at room temperature for 1 h, and visualized by ECL detection reagent (GE Healthcare).
Results and Discussion
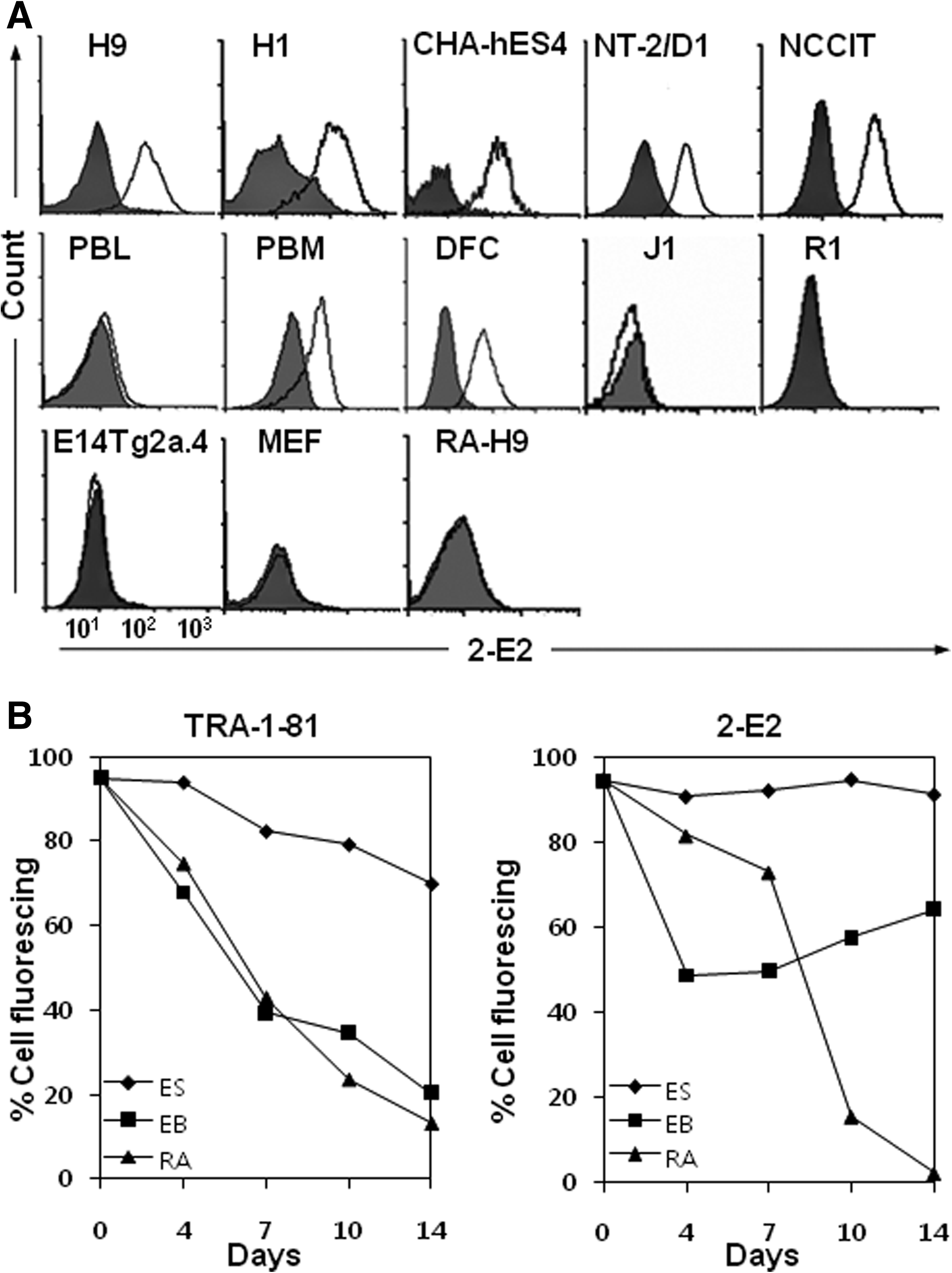
To identify and characterize cell surface molecules on pluripotent and undifferentiated hESCs, we previously generated 70 MAbs against hESCs using a modified decoy immunization strategy.(7) Most MAbs bound to undifferentiated hESCs, but only weakly or not at all to differentiated hESCs. As an attractive result, MAb 2-E2 only was shown to bind PBMCs, which was used as a differentiated primary human cell. Then 2-E2 was purified from ascitic fluid and used for further characterization in this study. In flow cytometric analysis, 2-E2 bound to human pluripotent stem cells H1, H9, CHA-hES4, NT-2/D1, and NCCIT, but not to mESC lines J1, R1, and E14Tg2a.4, and mouse embryonic fibroblasts (MEFs) (Fig. 1A). When PBMCs were gated on peripheral blood lymphocytes (PBLs) or peripheral blood monocytes (PBMs), 2-E2 did not bind to PBLs, but bound to PBMs. 2-E2 binding capability was also observed in DFCs (Fig. 1A), whereas it did not bind to RA-induced differentiated H9 cells (Fig. 1A). Therefore, we hypothesized that 2-E2 antigen expression may fluctuate during differentiation of hESCs.
(
In order to dissect the surface expression of 2-E2 antigen during differentiation, H9 cells were differentiated for 14 days by two different ways––RA treatment and EB formation––and the levels of 2-E2 antigen were measured by using mean fluorescence intensities from flow cytometric analyses (Fig. 1B). The levels of the hESC surface marker TRA-1-81 and 2-E2 antigens were generally maintained in undifferentiated H9 cells, although slight decreases were observed due to spontaneous differentiation over time. When H9 cells were treated with RA, the levels of TRA-1-81 and 2-E2 antigens was rapidly decreased, although the level of 2-E2 antigen was slightly decreased until day 7. The result was expected from our previous study.(7) When EB cells were examined for the level of TRA-1-81, the level of TRA-1-81 was rapidly decreased over time as expected. The level of 2-E2 antigen was also decreased in EB cells, and approximately 50% of EB cells remained positive for 2-E2 antigen at day 4 (Fig. 1B). However, the level of 2-E2 antigen was gradually increased in subsequent differentiation. The results suggest that 2-E2 antigen may regulate the early stage of hESC differentiation.
In order to identify the cell surface antigen recognized by 2-E2, the surface proteins of H9 cells were biotinylated, immunoprecipitated with 2-E2, and visualized with SA-HRP. MAb 2-E2 immunoprecipitated ∼68 and 27 kDa proteins from H9 cells (Fig. 2A). The same result was obtained from NT-2/D1 cells (Fig. 2B). In Western blot analysis, however, 2-E2 was not able to detect the 68 and 27 kDa proteins (data not shown), suggesting that 2-E2 may recognize the 2-E2 antigen in a conformation-dependent manner.
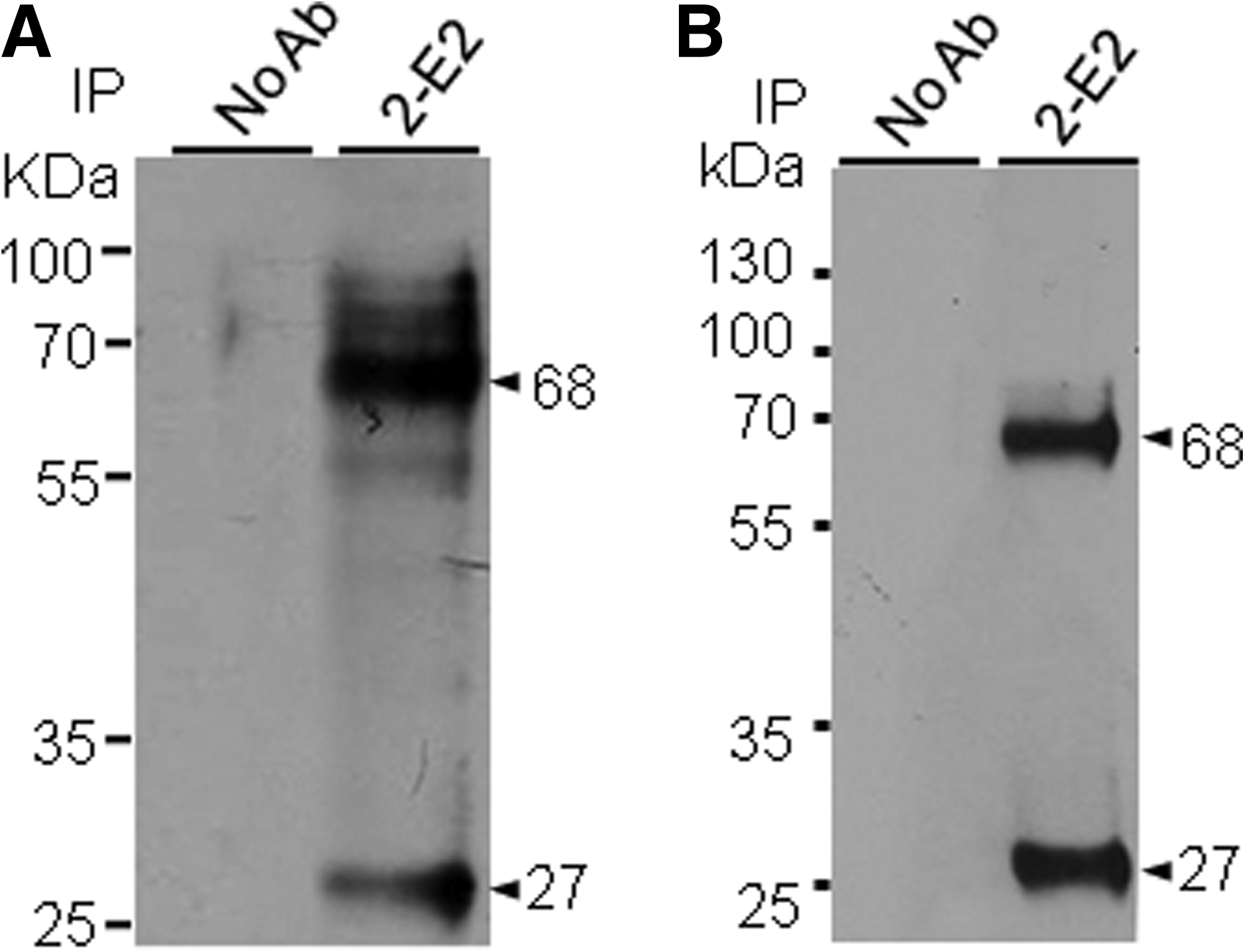
Detection of 2-E2 antigen. H9 cells (
As expected from our previous report,(7) 2-E2 binding capacity to hESCs was downregulated during RA-induced differentiation (Fig. 1). Interestingly, when the level of 2-E2 antigen was examined during EB formation, it gradually increased after initial downregulation (Fig. 1B), indicating that 2-E2 antigen may be re-expressed in adult differentiated primary cells. Consistent with this notion, we could detect 2-E2 antigen expression in DFCs and PBMs. Therefore, 2-E2 antigen would be a surface protein that plays a role in the transition period from self-renewal phase to differentiation phase of human pluripotent stem cells. 2-E2 antigen turned out to be 68 and 27 kDa cell surface proteins (Fig. 2). However, the 27 kDa protein was not always detected in cell surface biotinylation and immunoprecipitation assay, suggesting that it may be the truncated form of 68 kDa protein (data not shown). Although we do not know the identity of 2-E2-specific 68 kDa protein at the moment, the molecular characteristics of the 2-E2-specific 68 kDa protein make the protein worthy of future study for use as an indicator of early differentiation of human pluripotent stem cells.
Footnotes
Acknowledgment
This study was supported by the National Research Foundation of Korea (nos. 2009-0084052, 2009-0053111, and 2010-0015752).
Author Disclosure Statement
The authors have no financial interests to disclose.