
Abstract
Japanese encephalitis virus (JEV) is a major public health threat in the Asia-Pacific region. The pre-membrane (PrM) protein of Japanese encephalitis virus is cleaved during maturation by the cellular protease into the structural protein M and a pr-segment. Here, we describe a procedure to generate monoclonal antibody (MAb) against JEV PrM/M protein and investigate its characteristics. Western blot analysis showed that the MAbs produced in this study were against JEV PrM/M specifically. Indirect immunofluorescence assay demonstrated that they could recognize native PrM/M protein in JEV-infected BHK-21 cells. Preliminary studies identified the epitope of the MAb with a set of synthesized overlapping peptides covering the whole length of PrM protein of JEV. The MAbs reported here may provide valuable tools for the further exploration of biological properties and functions of PrM/M protein and may also be developed for potential clinical applications.
Introduction
JEV belongs to the family Flaviviridae, genus Flavivirus. It possesses a single-strand, positive sense, approximate 11 Kb RNA genome that contains a long, open reading frame. A polyprotein, encoded by the ORF, is cleaved co- and post-translationally into three structural proteins (C, prM/M, and E) and seven non-structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5).(7)
The PrM protein has a length of 167 amino acids, with a molecular weight of about 20 to 26 kDa.(8) PrM is the precursor of the membrane-anchored and virion-associated M protein (about 8 kDa). After synthesis of the nascent polyprotein of PrM and E, the signal cleavages generate the N and C termini of PrM. The C terminus containing the adjacent transmembrane domains allows PrM to anchor itself to the membrane and serves as the signal sequence for the translocation of E.(7,9) PrM is also associated with the E protein folding into the native conformation.(10) Additionally, PrM interacts with E to form PrM-E heterodimers and formed immature virions.(11) When the virus is released from the cell, the PrM protein is cleaved by furin or related protease in the trans-Golgi apparatus and the proteolytic cleavage of PrM to M protein generates mature virions.(12) However, the functional role of PrM in replication, virus infectivity, and induction of neutralizing activity is not fully understood.
In order to develop a useful tool for studying PrM/M protein of flavivirus, in the present study we have generated a hybridoma cell line secreting monoclonal antibodies (MAb) against JEV PrM/M protein and have investigated some of its characteristics.
Materials and Methods
Cell lines, virus, and other reagents
Baby hamster kidney (BHK-21) cells and SP2/0 myeloma cells were cultured in RPMI-1640 medium (Hyclone, Beijing, China) supplemented with 10% fetal calf serum (PAA, Somerset, United Kingdom) and antibiotics (100 IU/mL penicillin and 0.1 mg/mL streptomycin). All cells were maintained in humidified 5% CO2 atmosphere at 37°C. The JEV strain SA14-14-2 was propagated in BHK-21 cells to prepare the antigen for WB and immunofluorescence assays. The JEV-positive sera were obtained from pigs reared in pig farms. The sera were first tested by indirect ELISA and latex agglutination test (LAT) for detection of antibodies to JEV.
Animals
Eight-week-old female BALB/c mice were purchased from the Experimental Animal Center of Harbin Veterinary Research Institute (Chinese Academy of Agricultural Sciences, Harbin, China). The experiments using animals were licensed by the Animal Ethics Committee of Harbin Veterinary Research Institute.
Construction of recombinant plasmid bearing PrM/M gene of JEV
Total RNA was extracted from JEV-infected BHK-21 cells via TRIzol (Invitrogen, Carlsbad, CA), according to standard protocol, and the cDNA was produced with SuperscriptII RNAse H(-) reverse transcriptase (Invitrogen). In order to amplify the full-length gene of PrM from the cDNA template, a pair of primers, containing BamH I and Xho I sites, were designed according to the sequence of JEV SA14-14-2 strain (GenBank accession no. AF315119). The upstream primer was 5′-GCA
Expression and purification of PrM/M protein
The confirmed recombinant plasmids were transformed into Escherichia coli BL21(DE3) (Novagen) cells for expression. Overnight cultures of the transformed cells were diluted 1:100 in 50 mL Luria-Bertani (LB) broth containing 100 μg/mL kanamycin at 37°C. When OD600 reached 0.6, 1 mM isopropyl-D-thiogalactopyranoside (IPTG; Sigma, St. Louis, MO) was added to the broth to induce expression and the cells were incubated for a further 3 h with agitation. Thereafter, bacterial cells were removed from the growth medium by centrifugation at 5000 g for 15 min and lysed by sonication in cold phosphate-buffered saline (PBS; pH 7.4). To investigate whether the recombinant protein was expressed as soluble or inclusion body, sodium dodecyl sulfate-polyarcylamide gel electrophoresis (SDS-PAGE, 12%) was performed. Results showed that the His-tagged protein expressed predominantly as an inclusion body. To prepare antigen for immunization, we purified fusion protein by SDS-PAGE. Briefly, the inclusion bodies were resuspended in 2 mL PBS; the samples were then mixed with an equal volume of sample loading buffer (50 mM Tris-HCl [pH 6.8], 100 mM DTT, 2% SDS, 0.1% bromophenol blue, and 10% glycerol) and subjected to SDS-PAGE. By soaking in 0.25 M potassium chloride for a few minutes, a clear, white band representing NS1 protein can be seen on the gel. The band was then excised, triturated, and added to a proper volume of sterilized PBS. After freeze-thawing three times, supernatant was then centrifuged at 10000 r/min for 15 min collected and used for immunization and ELISA coating antigen.
Generation, selection, and purification of PrM/M specific MAb
Eight-week-old, female BALB/c mice were immunized subcutaneously with recombinant protein emulsified with Freund's complete adjuvant (Sigma). The mice were then given two booster injections of purified protein emulsified with Freund's incomplete adjuvant. A final intraperitoneal booster immunization of soluble PrM protein alone was given 3 days prior to the harvest of spleen cells for hybridoma fusion. General MAb preparation procedures were performed as described previously with minor modifications. Splenocytes from immunized animals were harvested aseptically and fused to the myeloma cell line SP2/0 with polyethylene glycol 50% (w/v) PEG 4000 (Sigma) at a splenocyte-myeloma cell ratio of 5:1. The fused cells were cultured and selected in HAT medium (RPMI medium with 10% fetal bovine serum, 100 μg/mL streptomycin, 100 IU/mL penicillin, 100 μM hypoxanthine, 16 μM thymidine, and 400 μM aminopterin); aminopterin was omitted from the medium on the twelfth day. Cell culture supernatants from the surviving clones were screened by indirect enzyme-linked immunosorbent assay (ELISA), and positive cell lines were subcloned thrice by limiting dilution method. To produce large quantities of the MAbs, selected hybridoma cells were injected intraperitoneally into pristine-primed mice, and ascitic fluids were collected 10 days after injections. Isotype of the MAbs was determined with a mouse MAb isotyping kit according to the manufacturer's instructions (Hycult Biotechnology, Uden, The Netherlands).
Western blot analysis
MAbs were analyzed by Western blot analysis with infected cells and recombinant proteins expressed in E. coli to determine their specificity and reactivity. Samples were mixed with an equal volume of 2×SDS sample loading buffer, boiled for 5 min, separated on 5% stacking/12% separating SDS polyacrylamide gels in a Tris-glycine buffer (0.025 M Tris base, 0.25 M glycine, 0.1% SDS), and transferred onto a nitrocellulose membrane with a Transblot apparatus (Bio-Rad, Hercules, CA). The membrane was washed three times in PBST with shaking, blocked with 5% skim milk at 4°C overnight, and probed with MAbs at 37°C for 1 h. Membranes were washed three times with PBST (0.1% Tween-20 in PBS) and incubated with anti-mouse Alexa Fluor 680-conjugated secondary antibodies (Invitrogen, Carlsbad, CA) for 1 h. Membranes were washed three times with PBST, then protein bands were detected with the Li-Cor Odyssey system (Li-Cor Biosciences, Lincoln, NE) and quantified using the Odyssey infrared imaging software (Li-Cor Biosciences).
Indirect immunofluorescence assays
BHK-21 cells were cultured as monolayers and infected with JEV. When 60−70% CPE was observed, cells were harvested and washed in PBS. Smears were coated with infect and mock-infect cells, air dried, and fixed with acetone. The MAbs were added to glass slides as primary antibody. The slides were incubated for 1 h at 37°C in a moist chamber. After washing three times with gentle shaking in PBS, 100 μL FITC-labeled goat anti-mouse IgG (Sigma), diluted 1:200 in PBS, was added and incubated at 37°C for 30 min. The smears were washed three times as described above, air dried, and mounted in glycerol. Finally, the cells were observed under the fluorescence microscopy (IMT2 Olympus, Tokyo, Japan).
Epitope mapping of MAb
To map the epitope of MAb against PrM/M protein of JEV, a set of 20 partially overlapping in length of 16 amino acids (with the exception of the last one, which was 15 in amino acid length) short peptides covering the whole PrM/M protein were synthesized (Table 1). The epitope peptide was screened by indirect ELISA using synthesized peptides as the coating antigen.
Results
Expression of PrM/M protein
To detect the expression of JEV PrM in E. coli, the bacteria lysate were analyzed by SDS-PAGE. Results indicated that the recombinant protein was successfully expressed after IPTG induction, which mainly expressed in the form of an inclusion body. Western blot analysis results showed that the recombinant PrM protein could be recognized by sera against JEV (Fig. 1).

Expression and Western blot analysis of JEV PrM protein. The bacteria harboring the recombinant plasmid were induced with IPTG and the bacterial protein was analyzed by SDS-PAGE. After transferring the protein to the NC membrane, it was probed with JEV-positive swine sera. M, protein marker; lane 1, bacterial protein harboring empty plasmid; lane 2, bacterial protein harboring PrM protein expressing plasmid; lane 3, bacterial protein harboring empty plasmid probed with JEV-positive swine sera; lane 4, bacterial protein harboring PrM protein expressing plasmid probed with JEV-positive swine sera.
Preparation of anti-PrM/M protein MAb
Five female BALB/c mice were immunized with the fusion protein. After thrice immunization, the mouse that presented the highest antibody titer to the immunogen was selected and given a booster injection so that its spleen cells could be used in cell fusion. For screening with ELISA, three positive clones were obtained. In order to avoid cross-reaction with His tag, His-E, a recombinant protein of His tag and JEV E protein prepared by our laboratory (data not published), was used as control. After three rounds of subcloning, one strain showed strong and specific reactivity to PrM protein and was designated as 3C8. After isotyping, it was found that 3C8 was of IgG1 type, and the light chains were κ type (Fig. 2).

Isotyping of monoclonal antibody. The isotype of the MAb 3C8 was identified with mouse MAb isotyping kit. The isotype is shown on the left. The developed dark point was the positive signal.
Biological activity of MAb 3C8
Indirect ELISA was performed to measure the MAb titers. Results showed that the antibody titers of ascites were over 204,800 (Fig. 3). The specificity and reactivity were further determined by Western blot analysis using JEV-infected BHK-21 cells. The results of Western blotting suggested that MAb 3C8 reacted with virus-infected cells (Fig. 4). These data revealed that the MAbs have good specificity and reactivity. To identify the ability of specific MAbs to recognize the native NS1 protein in JEV-infected cells, indirect immunofluorescence assay was carried out. A bright green signal could be detected, showing that the MAb recognized the native PrM/M protein (Fig. 5).
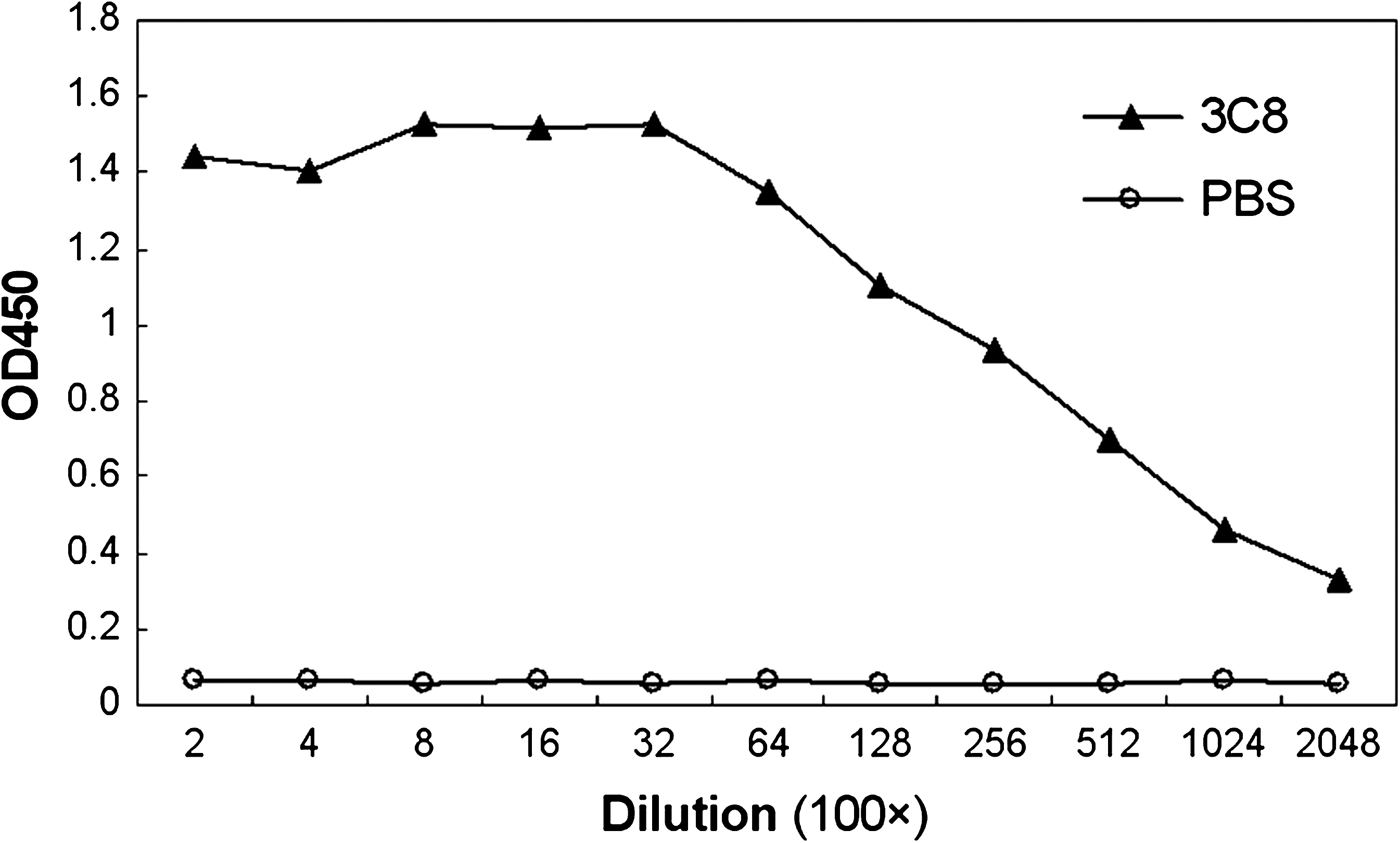
Titration of MAb 3C8 by ELISA. The ascites of MAb 3C8 was titered with indirect ELISA. PBS was used as the negative control. The dilution titer of MAb 3C8 ascites was over 204,800.
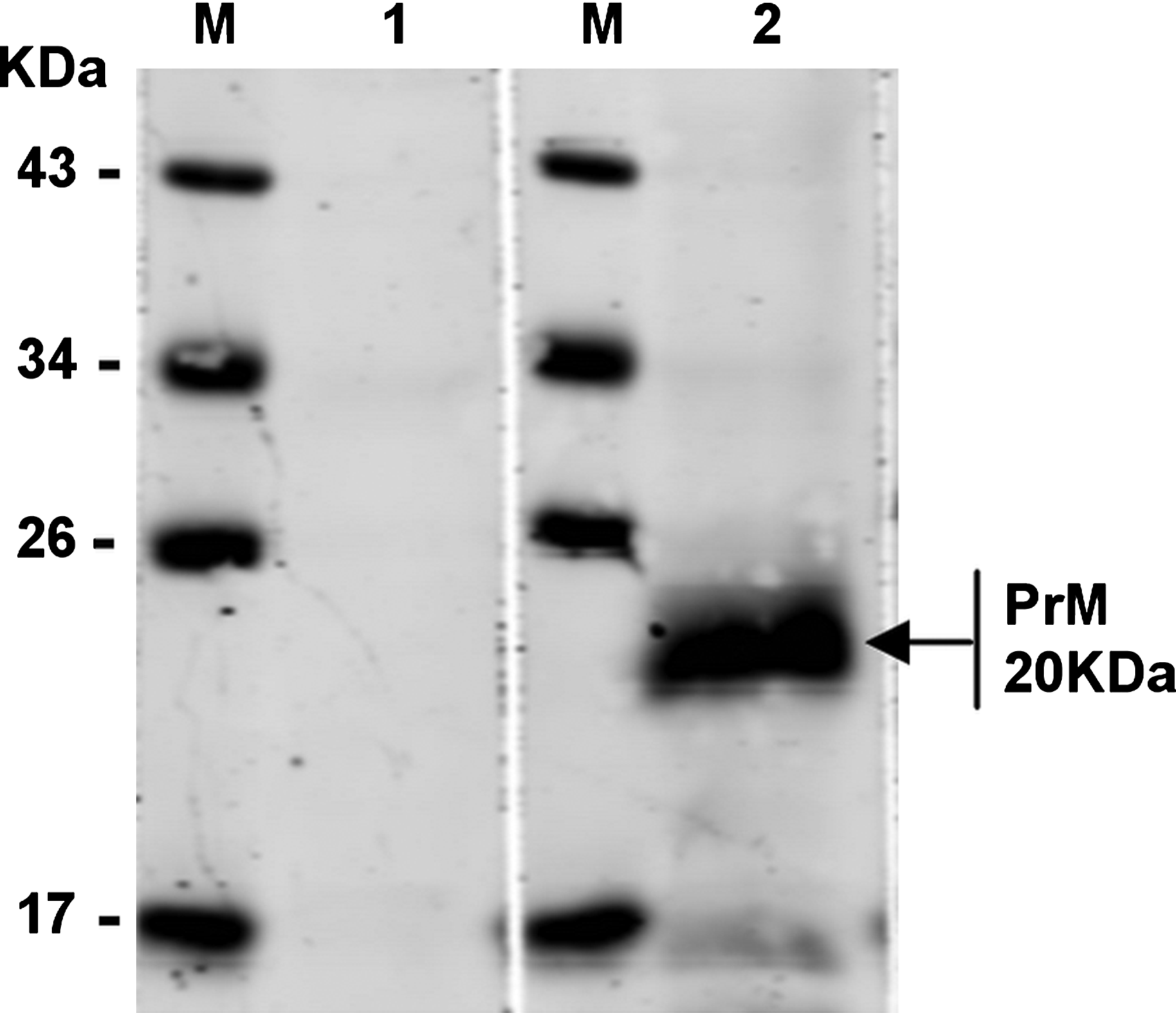
Western blot analysis shows the reaction to MAb 3C8 with JEV-infected BHK-21 cell. BHK-21 cells infected with JEV or mock infected were analyzed by SDS-PAGE and transferred to NC membrane, probed with the MAb 3C8, and developed with goat anti-mouse IgG. M, protein marker; lane 1, mock-infected cell lysate; lane 2, BHK-21-infected cel lysate.

Indirect immunofluorescence assay analysis shows the reaction of MAb 3C8 with native PrM/M on JEV-infected cells. (
Epitope mapping of MAb 3C8
With a set of overlapping synthesized peptides as coating antigen, the linear epitope of MAb 3C8 was mapped. Among these 20 peptides, MAb 3C8 was only strongly reactive with VNKKEAWLDSTKATRY. Other peptides, especially those containing eight amino acids overlapping with the epitope in the N and C termini, were not at all reactive with MAb 3C8 (Table 1).
Discussion
Although JEV is mainly prevalent in tropical and subtropical regions of Asia, the epidemiological trend of the JEV outbreak has changed since the virus was isolated in Australia.(1,2) This suggests that JEV has spread to regions beyond its ecological boundaries and there is concern that JEV might become a global threat. In fact, it is not unusual to find two or more flaviviruses co-circulating in one area. In Southeast Asia, the most important flaviviruses are JEV and dengue virus (DENV).(13) In northern Australia, Kunjin virus is found to co-circulate with JEV.(14) In Vladivostok, Russia, studies have reported the detection of WNV in birds.(15) In addition, there is evidence of WNV infection in India from Japanese encephalitis endemic and nonendemic areas.(16) Flaviviruses, such as WNV, DENV, and JEV, share some common features, such as transmission via mosquitoes, and cross-react with each other in serological tests. These cross-reactive responses could confound the interpretation during serological testing, including neutralization tests and ELISA.(17)
Previous reports,(18,19) however, show that in Western blot (WB) prM protein may be used to serologically differentiate individuals infected with JEV from those infected with DENV, WNV and SLEV. Hence, prM and antibodies against prM would be useful for conducting seroepidemiological studies of flavivirus infections in the regions that have prevalence of more than one flavivirus.
One of the most powerful tools in the revelation of protein properties represents the use of MAbs, which have been successfully applied for antigenic analysis, clinical diagnosis, and treatments. There are few reports about the production of JEV PrM specific MAbs. In this study, using a prokaryotic expressed recombinant protein as antigen to immunize the mice, we developed an MAb against JEV PrM protein using the hybridoma technique. Our results indicated that the resulting MAb 3C8 recognized the PrM/M protein specifically. Indirect immunofluorescence assay showed that the MAb could react with native PrM/M protein in JEV-infected BHK-21 cells. We believe that the MAb generated in this study would be important for delineating undiscovered characteristics and functions of PrM/M protein, and may also be useful for clinical application in the diagnosis and therapeutics for controlling JE.
Footnotes
Acknowledgments
This work was supported by a grant from the National Natural Science Foundation of China (30700027) and a special fund for agro-scientific research in the public interest of China (200803015).
Author Disclosure Statement
The authors have no financial interests to disclose.