
Abstract
Chitinase 3-like 2 (CHI3L2) is one of the most overexpressed genes in glioblastoma. Despite this, both the CHI3L2 gene and its protein product CHI3L2 are poorly characterized. Here we report the generation and characterization of monoclonal antibodies to CHI3L2 protein (CHI3L2 MAbs). Bacterially expressed 6 His-tagged full-length CHI3L2 was used as antigen. Spleen cells from immunized mice were collected and fused with SP2/0 myeloma cells. Hybridoma clones 2D3 and 4D2 producing high titer CHI3L2 MAbs were identified by enzyme-linked immunosorbent assay (ELISA) and further examined for their activity with the CHI3L2 protein by Western blot analysis and immunoprecipitation. The 2D3 clone was chosen for mouse inoculation and ascites formation. Antibodies derived from the ascitic fluid specifically recognized the recombinant CHI3L2 protein and strongly interacted with CHI3L2 in glioblastoma tissue lysate, as determined by Western blot analysis. The antibodies generated may be useful as a tool in various aspects of CHI3L2 investigation.
Introduction
The protein most thoroughly investigated in the chitinase-like family is CHI3L1, which has been studied in detail. Expression of CHI3L1, as well as CHI3L2, was found in synovial cells and chondrocytes, but was not detected in the brain, spleen, pancreas, or liver.(1) In contrast to CHI3L2, no CHI3L1 mRNA could be detected in the lung and heart. CHI3L1 expression was increased significantly in various tumors and cell lines derived from such tumors, including tumors of the brain, bone, breast, ovary, and lung.(8) In the same manner as CHI3L1,(9) increasing CHI3L2 gene expression was shown in glioblastomas using serial analysis of gene expression (SAGE), Northern blotting, and polymerase chain reaction (PCR). Our preliminary studies indicated that elevated expression of CHI3L2 in glial tumors may serve as a prognostic marker.(10) A clue to CHI3L2 function could be revealed from its structural similarity to CHI3L1 in size and nucleotide and amino acid sequences (52% and 56% homology, correspondingly). They have the same NH2-terminal amino acid residues as is shown in their alternative names (YKL-40 and YKL-39).(1) Recently, it has been reported that their 3D structures are also similar.(11) CHI3L2, similarly to CHI3L1, induced the phosphorylation of extracellular signal regulated kinases 1 and 2 (Erk1/2)(12); however, it appears to have an opposite effect on cell proliferation.(13)
Production and usage of polyclonal anti-serum against CHI3L2 protein was described by our lab previously,(10) and recently such antibodies have begun production by several companies, such as Santa Cruz Biotechnology (Santa Cruz, CA), R&D Systems (Minneapolis, MN), Novus Biologicals (Littleton, CO), and Gene Tex (Irvine, CA). However, use of monoclonal antibodies are often preferred due to their high specificity and significantly less signal-to-noise ratio. Therefore, the focus of this work was to develop and characterize monoclonal antibodies, which could be used for specific and sensitive detection of CHI3L2 protein.
Materials and Methods
Cell culture and cell culture media
Escherichia coli cells BL21 (Lys E) (Novagen, Madison, WI) were grown in lysogeny broth (LB) media (Fluka, Buchs, Switzerland). SP2/0 myeloma cells were obtained from Russian cell culture collection (Institute of Cytology of the Russian Academy of Sciences, St. Petersburg, Russia). A total of 293 cells were a kind gift of Dr. V. Filonenko (NAS of Ukraine, Kyiv). Roswell Park Memorial Institute medium (RPMI) 1640 and fetal calf serum (FCS) were purchased from Gibco (Grand Island, NY). Polyethylene glycol (PEG)-2000 was from Merck (Darmstadt, Germany), and Protein G-Sepharose CL-4B was from GE Healthcare (Piscataway, NJ).
Production of recombinant CHI3L2 protein
Generation of human recombinant CHI3L2 was performed according to our previous description.(10) Briefly, E. coli cells transformed by recombinant plasmid pET-24a(+)-CHI3L2 were grown at 37°C to optical density 0.5–0.7 (600 nm) and synthesis of recombinant protein was induced by the addition of isopropyl β-D-thiogalactopyranoside (IPTG, Sigma, St. Louis, MO) for 3 h at a concentration of 100 mM. Affinity purification using nickel-nitriloacetic acid (Ni-NTA)-agarose (Qiagen, Valencia, CA) was carried out under denaturing conditions in the presence of 8 M urea.
Recombinant and native human CHI3L1 were kindly provided by Dr. A. Iershov (Dept. of Biosynthesis of Nucleic Acids, Institute of Molecular Biology and Genetics, NAS of Ukraine, Kyiv).
Electroelution of proteins from SDS-PAGE
Purified recombinant CHI3L2 protein was electrophoresed on 10% sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) with subsequent Coomassie Brilliant Blue R250 (Fluka) staining. The protein bands were cut from the gel with a razor blade, unstained, sliced into small pieces about 1 mm×1 mm, and then placed into dialysis bags. The electroelution of protein was performed in gel running buffer (25 mM Tris, 192 mM glycine, 0.1% SDS [pH 8.3]) under the following conditions: 100 V, 1 h. Electroeluted proteins were dialyzed twice against phosphate buffered saline (PBS).
MALDI-TOF mass spectrometry analysis
The protein spots (1–2 mm in length) were excised from Coomassie Brilliant Blue R250 stained SDS-PAGE and transferred to a microcentrifuge tube. The excised gel was washed with 200 μL of deionized water, followed by washing with 25 mM Tris-HCl (pH 8.5) and 50% acetonitrile (Fluka) until becoming colorless. The gel was dried in a vacuum centrifuge for 30 min, and incubated with 15 μL of sequencing-grade modified trypsin (12.5 ng/μL; Promega, Madison WI) in 50 mM ammonium carbonate at 37°C for 8 h. The digested polypeptides were recovered in 10 μL solution containing 0.1% (v/v) trifluoroacetic acid (TFA, Fluka) and 50% (v/v) acetonitrile. MALDI-TOF analysis was carried out on an Ultraflex II MALDI-TOF/TOF instrument (Bruker-Daltonics, Billerica, MA). The mass spectrometer was operated with positive polarity in reflectron mode and spectra acquired in the range of m/z 500–6000 at 20 kV accelerating voltage and laser frequency of 200 Hz. The mass values for monoisotopic peaks of obtained spectra were used for searches against Mascot database (
Production of polyclonal anti-serum
Production of rabbit polyclonal anti-serum against human CHI3L2 protein and Western blot analysis were performed according to our previous description.(10) RPMI medium with 20% FCS and serum of non-immunized mice were used for control of non-specific binding.
For Western blot of cross-reactivity, 30 ng of recombinant CHI3L2 or recombinant and native CHI3L1, as well as 40 μg of lysed 293 cells expressing CHI3L2 or CHI3L1, were loaded onto gel and immunoblotted with either CHI3L2 MAbs or polyclonal anti-CHI3L1 antibodies (sc-30465, Santa Cruz Biotechnology, Santa Cruz, CA).
Production of hybridomas and CHI3L2 MAbs
Full-length CHI3L2 protein was used for hybridoma generation according to standard protocol(16) with some modifications. Female BALB/c mice (8 weeks old) were immunized with 45 μg of recombinant 6 His-tagged CHI3L2 protein in complete Freund's adjuvant by intraperitoneal (i.p.) injection. Subsequently, 45 μg of recombinant protein mixed with incomplete Freund's adjuvant (Sigma) were used four times for immunization by i.p. injection. After immunization with boosting every 2 weeks, the total titer of anti-CHI3L2 antibodies in blood serum was monitored by ELISA. The titer of antibodies to CHI3L2 protein in the serum of immunized mice was 10−6 after 45 days. At the next stage, the mouse that showed the best response was inoculated with 30 μg of antigen by i.p. injection in PBS without adjuvant. Three days later, splenocytes from this mouse and SP2/0 myeloma cells cultured in RPMI 1640 with 20% FCS were fused in the presence of PEG-2000. The positive hybrids were selected by ELISA.
For production of ascitic fluid, BALB/c mice were injected with 0.5 mL of pristane (Sigma) and 7–10 days later inoculated with 5×106 hybridoma cells. The ascitic fluid was collected after 10–14 days. The fraction of immunoglobulins was precipitated from ascitic fluid with 50% ammonium sulphate and used for affinity purification by Protein G-Sepharose CL-4B chromatography. The IgG fractions were pulled together and dialyzed against PBS. The purified antibodies were aliquoted and stored at −70°C.
ELISA assay
CHI3L2 (0.5 μg/well) in PBS was incubated in 96-well polystyrene plates for 1 h at 37°C. The plates were then washed with PBS-0.1% Tween-20 (PBS-T), and incubated with 200 μL of 2% bovine serum albumin (BSA) in PBS added to each well for 1 h at 37°C in order to block non-specific binding. Subsequently, the wells were loaded with 100 μL aliquots of hybridoma media and incubated for 1 h at 37°C followed by four washes with PBS-T. After washing, 100 μL of horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG antibodies (1:5000 v/v, Promega) were added and incubated for 1 h at 37°C. Plates were washed again four times with PBS-T, then substrate solution, containing 0.02% H2O2, 0.1 M citrate-phosphate buffer (pH 5.8), and 0.5 mg/mL 2,2'-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) sodium salt (Sigma), was added to each well. After 15 min incubation at 37°C, the absorbance was measured at 490 nm using Sunrise plate reader (Tecan, Männedorf, Switzerland). RPMI medium supplemented with 20% FCS was used as a negative control.
Immunoprecipitation
CHI3L2 MAbs in amounts of 4, 10, and 20 μg, or rabbit polyclonal anti-serum at dilution 1:100, 1:50, and 1:10 were pre-incubated with 20 μL of 30% Protein A-Agarose slurry (Sigma), centrifuged, and washed twice in PBS. Immunoprecipitation was performed with 1 μg of CHI3L2 recombinant protein, 600 μg of glioblastoma tissue lysate, or 300 μg of lysed 293 cells transfected by pcDNA3.1_CHI3L2. Reaction mixture was rotated overnight at 4°C in a total volume of 500 μL of lysis buffer (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1% Triton X-100, 50 mM EDTA, and a mixture of protease inhibitors [Roche Molecular Diagnostics, Meylan, France]). Immunoprecipitates were recovered by brief centrifugation and washed three times with 1 mL of lysis buffer before boiling for 5 min in 4x SDS-PAGE sample buffer. The samples were subjected to Western blot analysis using rabbit polyclonal anti-serum to CHI3L2 or CHI3L2 MAbs. Protein bands on nitrocellulose membrane were visualized by Ponceau S (Sigma) staining (0.1% w/v in 5% v/v acetic acid). Negative controls were performed with 600 μg of normal brain tissue lysate or 300 μg of lysed non-transfected 293 cells. Control for non-specific binding of recombinant CHI3L2 to Protein A-Agarose without antibodies was performed with 1 μg of recombinant CHI3L2 and 20 μL of 30% Protein A-Agarose slurry.
Generation of 293 cells, transiently expressing CHI3L2
The 1.2-kb DNA fragment encoding CHI3L2 was amplified by a PCR using clone IMAGp998E049409Q3, entry # 4148715 (IMAGE collection), which contains full length CHI3L2 cDNA. The following primers were used: CHI3L2pcDNA3for (5′-CTAAGCTTGGCCAAGGATATGGGAGCAACCAC-3′) and CHI3L2pcDNA3FLAGrev (5′ATGGATCCTCATTTGTCATCGTCATCCTTGTAGTCCAGGGAGCCAAGGCTTCT-3′), with additional nucleotides on the 5′-end for generation of restriction sites AAGCTT (Hind III) and CTCGAG (BamH1). CHI3L2pcDNA3FLAGrev also contained a sequence for FLAG tag (GACTACAAGGATGACGATGACAAA). PCR was performed under the following conditions: 30 cycles at 94°C for 1 min, 56°C for 1 min, and 72°C for 1.5 min. Hind III/Bam HI fragment of CHI3L2 cDNA was ligated to the Bam HI/Hind III site of the pcDNA3.1(+)/neomycin mammalian expression vector (Invitrogen, Carlsbad, CA). 293 cells were transfected by 6 μg of pcDNA3.1_CHI3L2 using 24 μL of polyethylenimine (Sigma) per 60-mm dish. CHI3L2 protein expression was verified by Western blot analysis.
Lysates of 293 cells, stably producing CHI3L1 (293_CHI3L1 cells), were kindly provided by Dr. O. Balynska (Department of Biosynthesis of Nucleic Acids, Institute of Molecular Biology and Genetics, NAS of Ukraine, Kyiv).
Results and Discussion
The decision regarding whether to use polyclonal or monoclonal antibodies depends on many factors.(14) Polyclonal antibodies can help to amplify a signal from the target protein as they bind multiple epitopes. However, affinity purification yields of polyclonal antibodies are small; moreover the quality of the anti-serum may vary from bleed to bleed. The principal advantages of monoclonal antibodies are high homogeneity, purity, and production in large amounts with less inter-batch variation. Some highly sensitive, specific, and rapid techniques include general advantages of both antibodies (e.g., sandwich-type ELISA).(15) Such detection systems are already developed for antigens, which are considered to be potential tumor markers, including prostate-specific antigen (PSA), prostatic acid phosphatase (PAP), squamous cell carcinoma antigen (SCC),(15) and CHI3L1.(8)
Here we describe the production of monoclonal antibodies, which may be useful for the detection of CHI3L2 and provide the opportunity to create a highly specific and sensitive ELISA method for this protein.
Antigen purification
To generate large quantities of recombinant CHI3L2 protein, we used a bacterial expression system. Electrophoresis of CHI3L2 after purification produced two major bands corresponding to proteins with a molecular mass of 43 and 35 kDa, despite the use of protease inhibitors (Fig. 1). Western blot showed that rabbit polyclonal anti-serum against a recombinant CHI3L2 recognized both of these proteins. The first band corresponded by size to the previously described native CHI3L2 (about 40 kDa).(1) The 3 kDa difference may be explained by the fact that recombinant CHI3L2 contains a 26-residue transmembrane signal peptide, which is absent in mature protein, and 6 His at C-terminus.
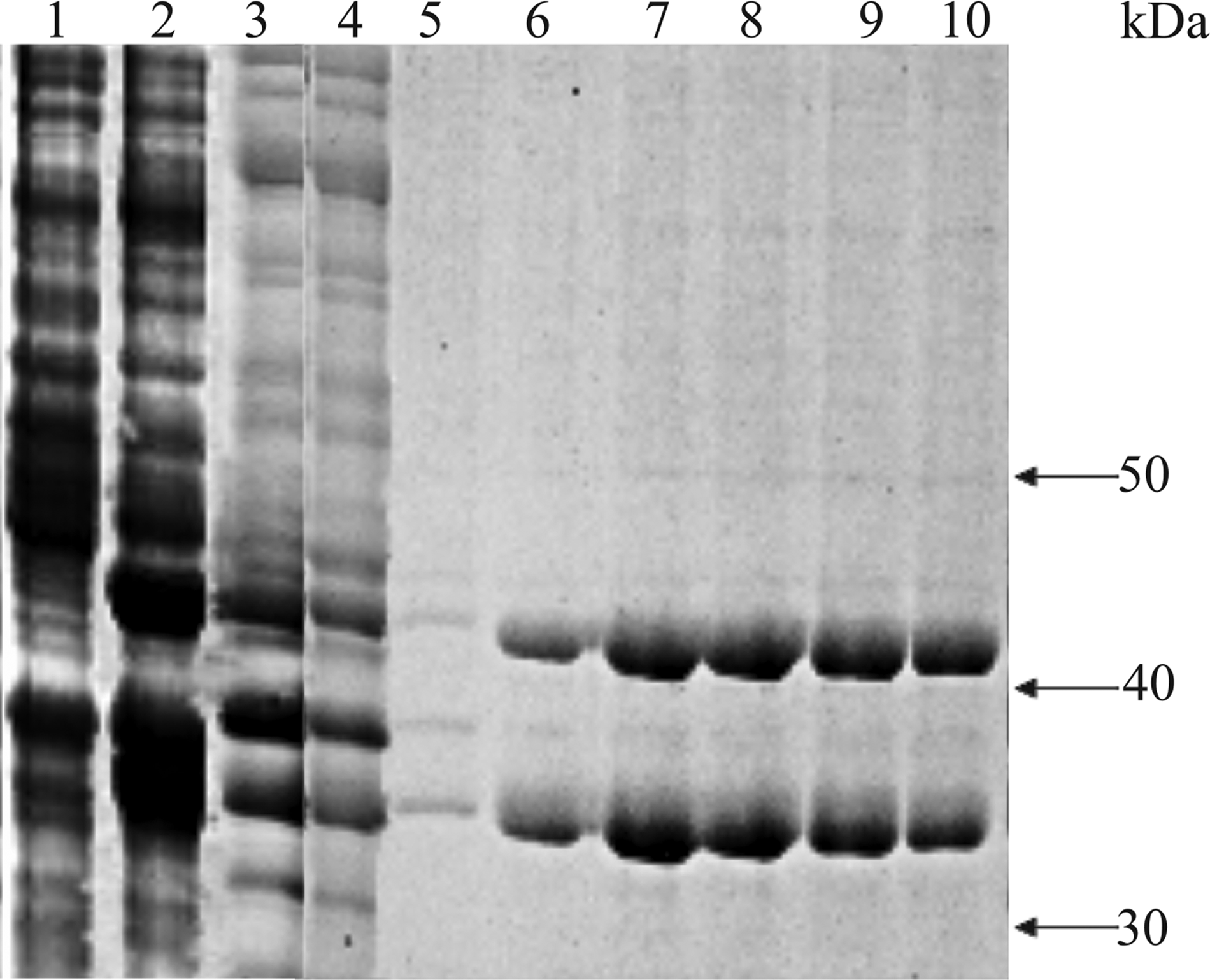
Isolation of recombinant CHI3L2 protein. SDS-PAGE electrophoresis: lysate of E. coli cells transformed with pET-24a-CHI3L2, without addition of 1
To identify the protein in the second band, it was electroeluted and checked for purity by SDS-PAGE electrophoresis and immunoblotting with rabbit polyclonal anti-serum (as shown in Fig. 2, it was more than 90% pure). The results of mass spectrometry analysis show that the 35 kDa protein in the second band is a short fragment of the CHI3L2 protein without about 80 NH2-terminal amino acid residues (Fig. 3).
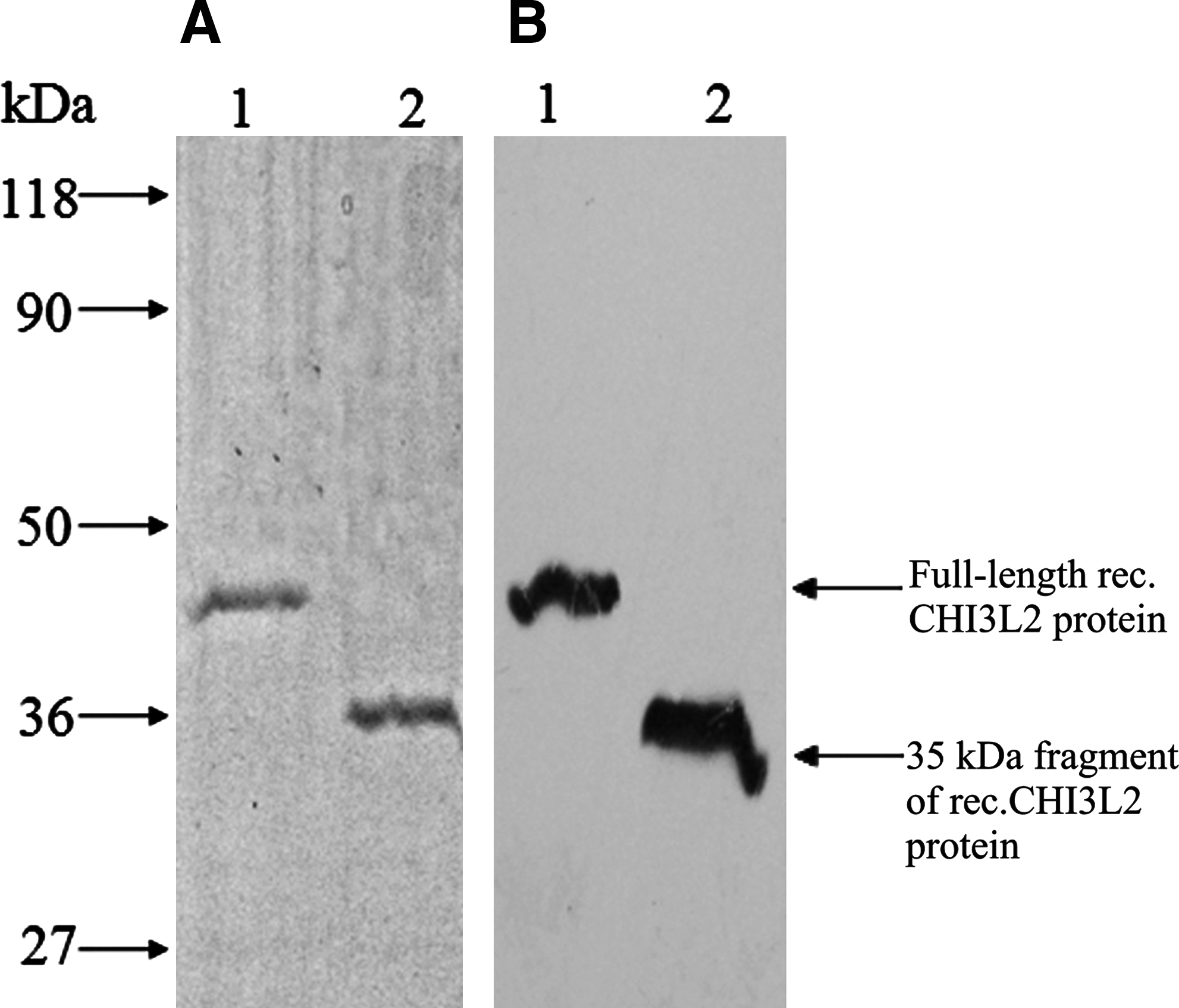
SDS-PAGE electrophoresis (
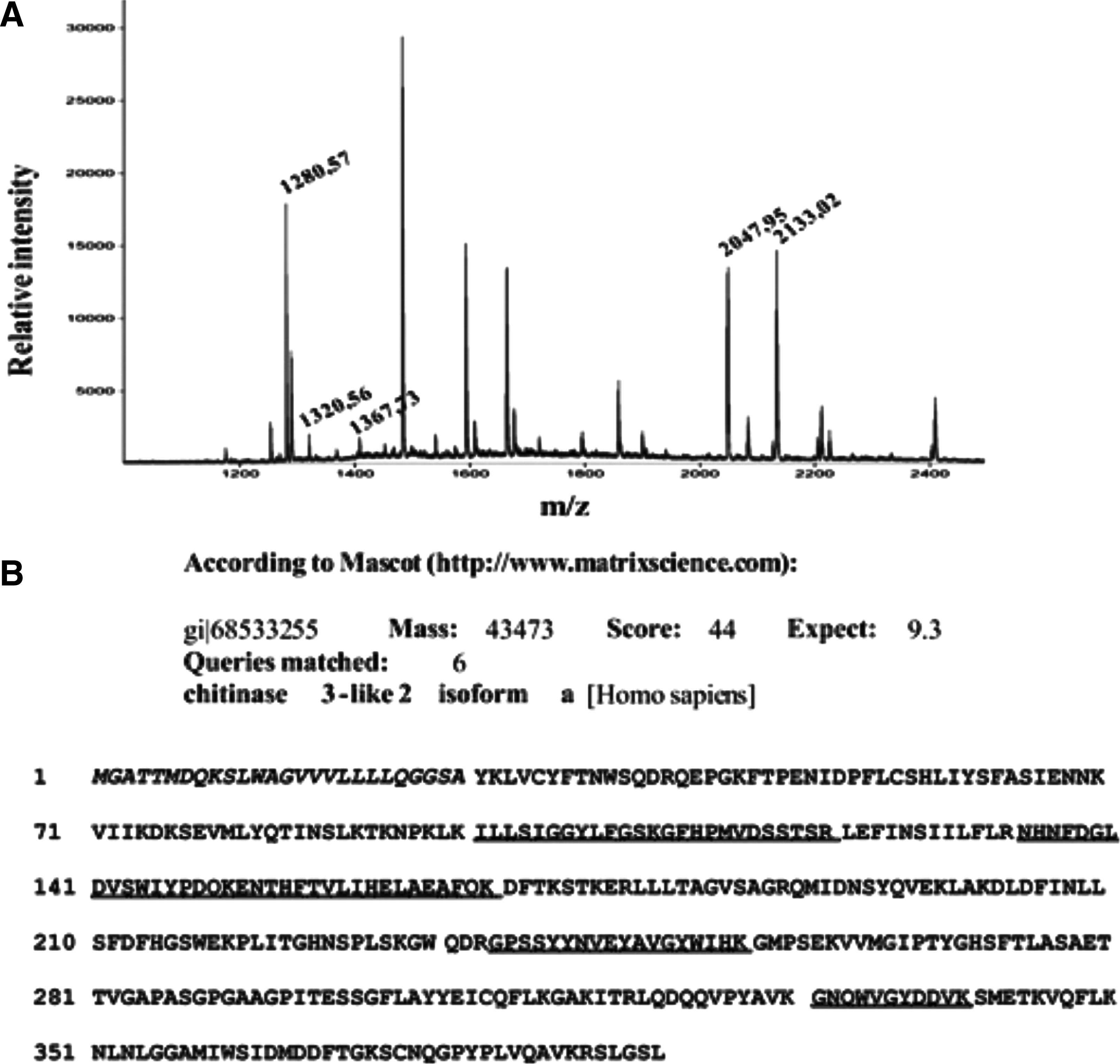
Mass spectrometry analysis of 35 kDa protein. (
Selection of positive clones
For selection of specific clones, obtained hybridoma cells were tested for the production of antibodies to CHI3L2 in ELISA assay. Three positive clones were identified by screening of 300 wells in five 96-well plates with growing hybridomas. In the second round of ELISA screening, two clones were selected, termed 2D3 and 4D2, that gave the strongest response to CHI3L2 protein.
The specificity of antibodies from culture media of both clones was confirmed by Western blot using full size CHI3L2 protein and glioblastoma tissue lysate (Fig. 4A and B). RPMI medium supplemented with 20% FCS was used as a negative control and showed no reactivity, both to recombinant CHI3L2 and glioblastoma tissue lysate (Fig. 4F). 2D3 cells were subcloned twice using limiting dilution method to guarantee monoclonal behavior of the produced immunoglobulins, while 4D2 was frozen and stored.
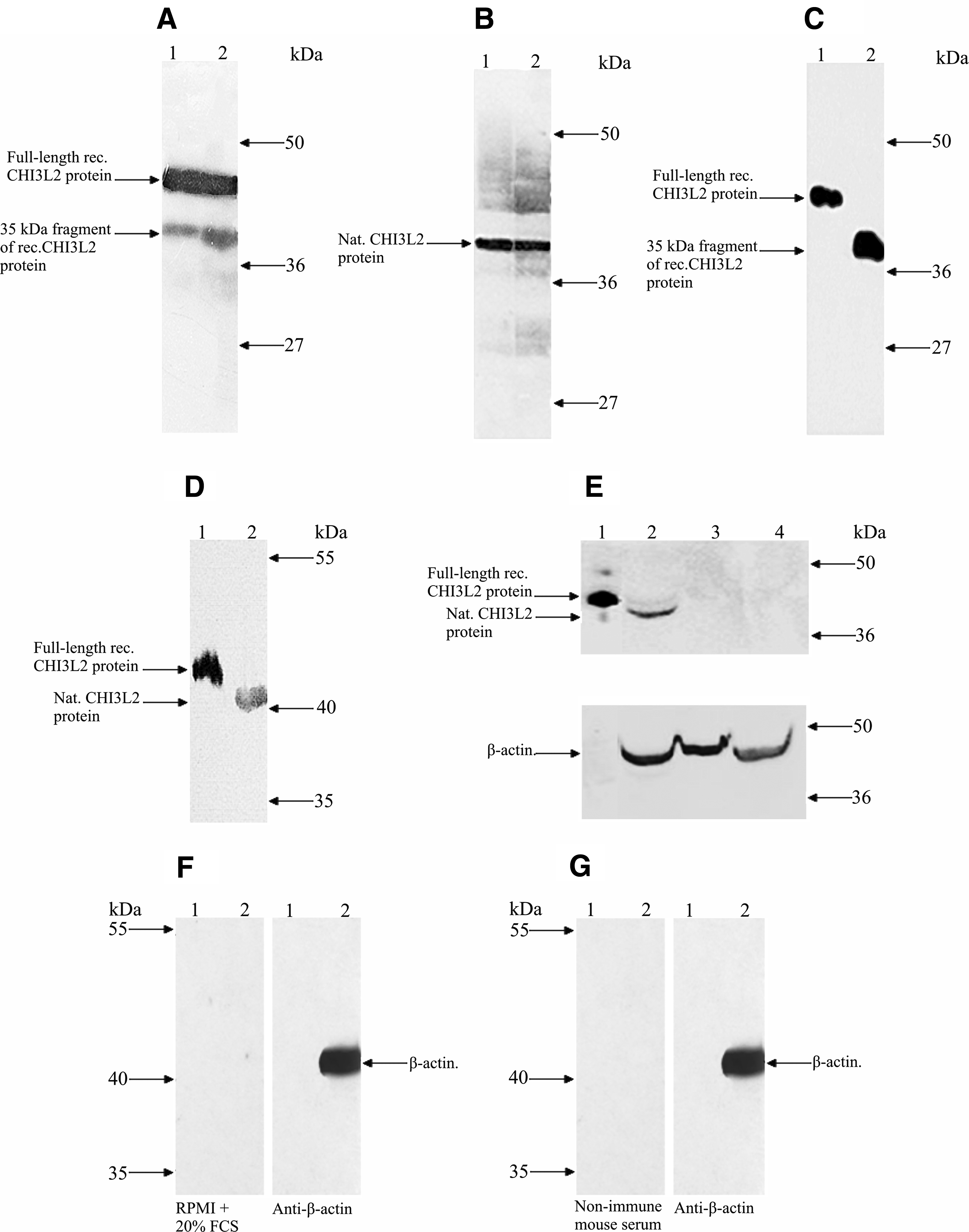
Application of CHI3L2 MAbs in Western blot. Detection of recombinant CHI3L2 (
CHI3L2 MAbs application in Western blot analysis
Ascitic fluid, obtained after inoculation of 2D3 clone, was used for purification of CHI3L2 MABs.
Purified antibodies were shown to be highly specific in Western blot. As seen in Figure 4C, CHI3L2 MAbs detect about 10 ng of full length and short fragment recombinant CHI3L2 protein at a concentration of 0.1–0.2 μg/mL, suggesting that the CHI3L2 MAb epitope is not localized within the NH2-terminus of the protein sequence. The same working concentration was also efficient for selective recognition of CHI3L2 protein in glioblastoma lysate and 293 cell lysates after transfection by pcDNA3.1_CHI3L2 (Fig. 4D and E). Pre-immune mouse serum did not react with the recombinant CHI3L2 or glioblastoma tissue lysate (Fig. 4G). CHI3L2 MAbs showed no cross-reactivity with recombinant and native human CHI3L1 protein, which is quite closely related to CHI3L2 (about 50% homology) (Fig. 5). The difference in molecular mass between recombinant and native CHI3L1 may be explained by the facts that recombinant CHI3L1 was obtained from cDNA without signal sequence and that native CHI3L1 is a glycoprotein.
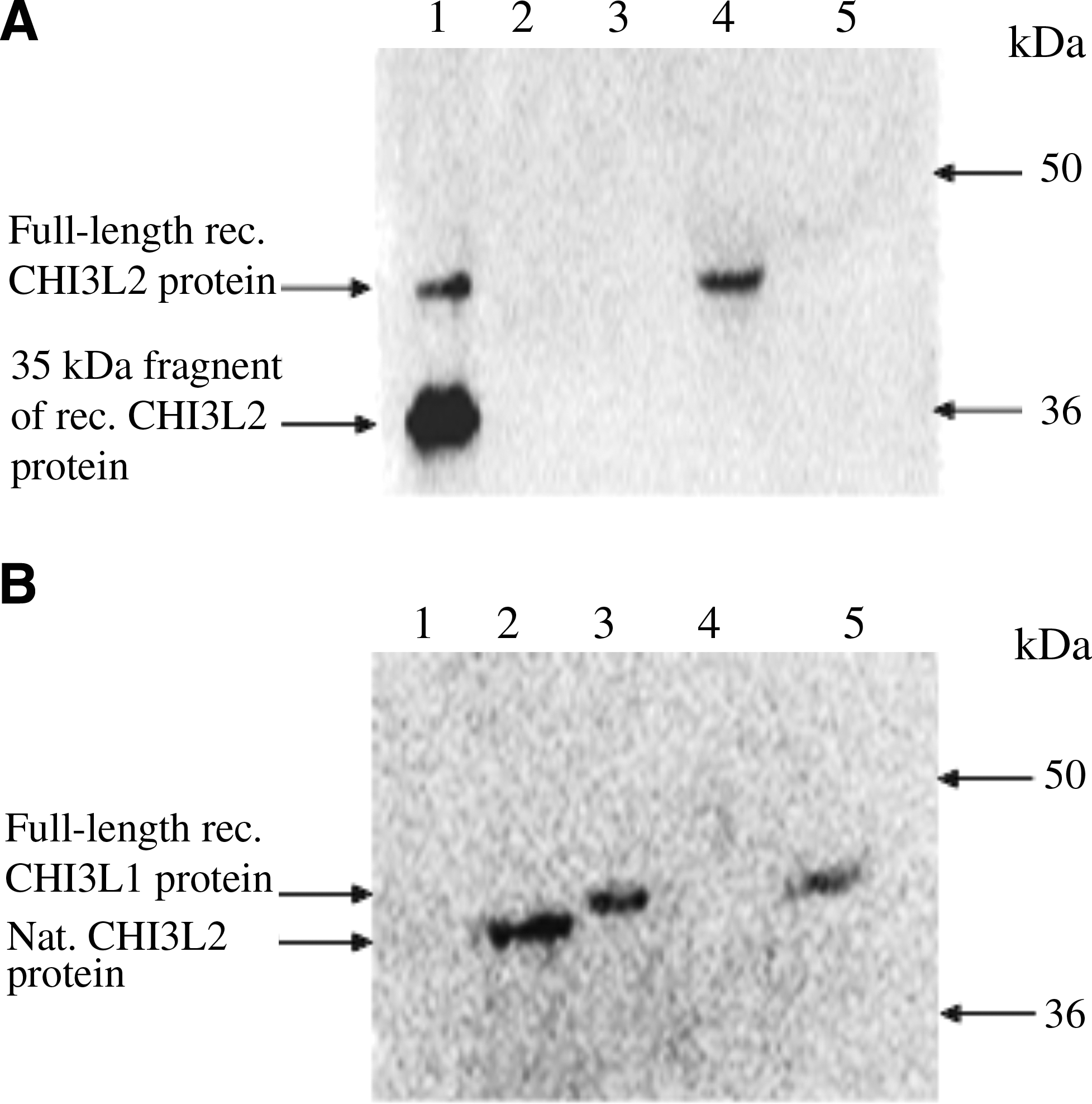
Cross-reactivity of CHI3L2 MAbs from mouse ascitic fluid with CHI3L1 protein. (
Application of CHI3L2 MAbs in immunoprecipitation
To determine whether CHI3L2 MAbs are suitable for immunoprecipitation, we performed this experiment in duplicate using protein eluate from Ni-NTA-agarose consisting of full length CHI3L2 protein with a high content of the 35 kDa fragment, glioblastoma tissue lysate, and 293 cells transiently transfected with CHI3L2. To determine a working amount of antibodies, 4 μg, 10 μg, and 20 μg of CHI3L2 MAbs and 1:100, 1:50, and 1:10 dilutions of rabbit polyclonal anti-serum were tested. The most effective were found with 20 μg of CHI3L2 MAbs and 1:10 dilution of rabbit polyclonal anti-serum per 1 μg of recombinant CHI3L2 protein (Fig. 6). CHI3L2 MAbs precipitated recombinant protein, but did not recognize CHI3L2 protein in glioblastoma tissue lysate or lysate of 293 cells transfected by pcDNA3.1_CHI3L2, where CHI3L2 was present in native form (Fig. 6A). The same results were obtained with rabbit polyclonal anti-serum (Fig. 6B). The bands of about 50 kDa in Figure 6A are the immunoglobulins from glioblastoma tissue lysate, which cross-react with anti-rabbit antibodies.
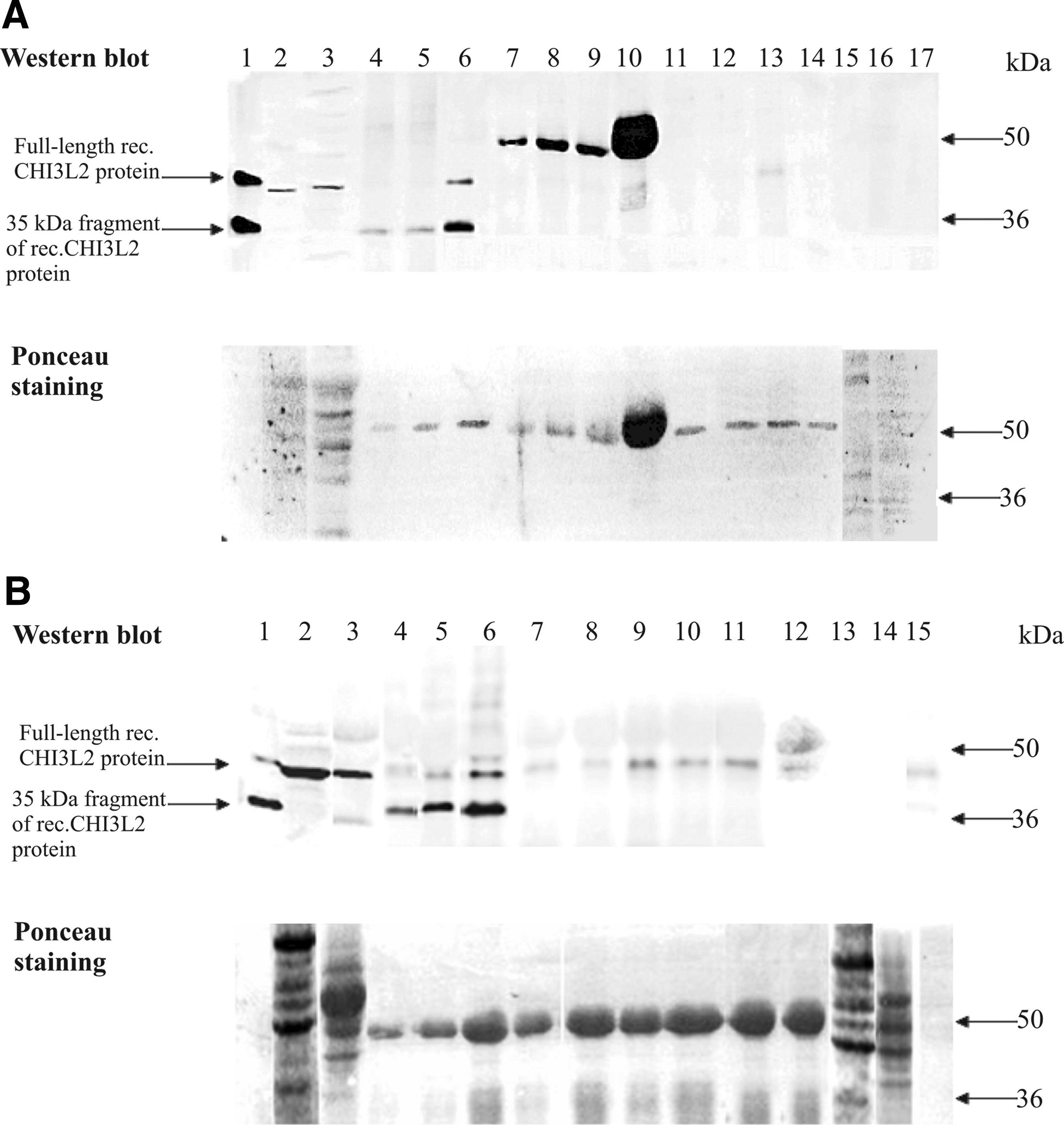
Western blot of immunoprecipitates obtained using CHI3L2 MAbs from mouse ascitic fluid (
Previously, it was shown that monoclonal antibodies, which worked efficiently in Western blot, as well as polyclonal anti-serum, were insufficient for immunoprecipitation of native protein. Thus, Evan and colleagues(17) reported on the inability to immunoprecipitate native p62 c-myc protein from cell lysates using one of six generated MAbs, and Richards and colleagues(18) demonstrated that rabbit polyclonal anti-sera directed against FATP1 and ACSL1 proteins do not immunoprecipitate them efficiently. Okuma and colleagues(19) showed the reduced ability of the C-terminal-directed anti-Gi anti-serum to immunoprecipitate the agonist bound α2a receptor and Gannon and colleagues(20) reported that monoclonal antibody against mutant p53 did not immunoprecipitate wild-type p53 but detected it well in Western blot. This phenomenon may be explained by several factors, including conformation-dependent epitopes, association with other proteins, post-translational modification, and specific folding, which may abrogate antibody-antigen reaction and markedly affect the function of antibodies.(14)
The research on CHI3L2 protein was started not long ago and many questions still remain. This study describes the production of monoclonal antibodies against CHI3L2 protein, which works efficiently in Western blot and is useful for investigation of CHI3L2 in health and disease.
Footnotes
Acknowledgments
We acknowledge Prof. Y. Zozulya for providing glioblastoma samples and Dr. I. Toropygin for his help in performing mass spectrometry analysis. This work was supported in part by the Science & Technology Center in Ukraine (Project 4688) and by the National Academy of Sciences of Ukraine within the program Fundamentals of Molecular and Cellular Biotechnologies, and SFFR of Ukraine (Project F4B/457-2011).
Author Disclosure Statement
The authors have no financial interests to disclose.