
Abstract
The baculovirus-insect cell system is considered a feasible expression system for recombinant glycoprotein production due to its several advantages, including high capacity, flexibility, and glycosylation capability. However, accurate titering of the recombinant baculovirus is required to ensure high expression in insect cells using a commercial and expensive immunoassay titer kit in which the envelope glycoprotein of the Autographa californica multiple nucleopolyhedrovirus (AcMNPV)-type baculovirus is detected by anti-envelope glycoprotein antibody and a secondary antibody conjugated to horseradish peroxidase (HRP). In this study, conditions for the expression of the CO17-1A immunotherapeutic monoclonal antibody (MAb) against colorectal cancer cells in a baculovirus system were optimized without using a commercial titering kit. Several variables were investigated to optimize antibody expression in a baculovirus-insect cell system, including baculovirus passage, volume of the infecting baculovirus inoculum (100, 200, 400, and 800 μL), and the harvest time of insect cells or cell supernatants after virus infection (24, 48, and 72 h). Two different pFastBac vectors carrying the CO17-1A MAb genes with or without the KDEL endoplasmic reticulum (ER) retention motif (Lys-Asp-Glu-Leu) fused to the HC (MAb CO17-1A K and MAb CO17-1A, respectively) were constructed and used to generate baculoviruses. Immunoblot analysis was conducted to confirm expression of MAb CO17-1A K and MAb CO17-1A in baculovirus-infected insect cells. Densitometry analysis of the protein bands was used to quantify the relative expression under different conditions. The highest expression was observed in lysed cells infected with 400 μL of passage 3 baculovirus (P3 BV) carrying the gene encoding the CO17-1A MAb without KDEL at 72 h after virus infection. These results suggest that the infection conditions, the number of virus passages, baculovirus inoculum volume, and the harvest time can be modified to optimize MAb expression without using a BaculoELISA titer kit in a baculovirus-insect cell system.
Introduction
Recently, a new method has been developed using the BaculoELISA titer kit, which represents an evolution of existing titration methods, and makes use of an AcNPV-type baculovirus.(9) This method is a standard ELISA, which enables us to determine baculovirus titers based on p35, an early expressed viral protein that prevents cellular apoptosis and is essential for infectivity within 24 h.(10,11) The BaculoELISA titer kit is relatively rapid and simple to use, and is more efficient and accurate. However, the control virus in this kit is still too costly to be used for virus titration.
The monoclonal antibody (MAb) CO17-1A (IgG2a) is a glycoprotein that plays an important role in the treatment of micrometastases and the recurrence of colorectal cancer in the colon mucosa.(12–14) The MAb CO17-1A detects the tumor-associated antigen GA733-2E, which is a major glycoprotein with a molecular mass of 40 kDa that is expressed in human colorectal carcinomas. The functional MAb CO17-1A has been produced in the baculovirus expression system.(15)
Glycoproteins are secreted through subcellular compartments directed by secretion-related signal peptides. The secretion pathway includes protein folding and assembly processing in the ER, transfer to the Golgi complex, and secretion through the outer membrane. When the ER retention-signal peptide KDEL is fused at the C terminus of a protein, it causes the protein to localize to and accumulate in the endoplasmic reticulum (ER), resulting in enhanced production in heterologous expression systems.(16)
Therefore, in this study, we investigated the effect of the ER retention signal KDEL on the expression of the anti-colorectal cancer MAb CO17-1A in the BES. In addition, in order to enhance antibody expression in the baculovirus-insect cell system without virus titration, various factors of the infection conditions were optimized, including the volume of the baculovirus inoculum, baculovirus passage (P2 and P3 of the baculovirus stock), and the harvest time (24, 48, and 72 h) of the insect cells.
Materials and Methods
Bacmid construction for MAb CO17-1A gene expression
A pFastBac dual vector (pFB, Invitrogen, Carlsbad, CA) carrying the anti-colorectal MAb CO17-1A HC and LC genes (pFB CO17-1A) was obtained as previously described(17) (Figs. 1 and 2). A second pFB carrying the HC gene fused to the ER retention signal peptide KDEL (HCK) and the LC gene (pFB CO17-1A K) was constructed (Fig. 1). Two different recombinant bacmids containing MAb CO17-1A and MAb CO17-1A K were generated with the plasmids pFB CO17-1A and pFB CO17-1A K according to the Bac-to-Bac system (Invitrogen) protocol. The HCK and LC genes of MAb CO17-1A K were cloned into the pFB vector under the control of the polyhedron (PPH) and P10 promoters, respectively.(18,19) The recombinant pFB CO17-1A K vector was transformed into Escherichia coli DH10Bac in order to transpose the MAb HCK and LC expression cassettes into the AcMNPV bacmid to yield CO17-1A bacmids expressing the HCK and LC genes under the control of the PPH and PP10 promoters, respectively.
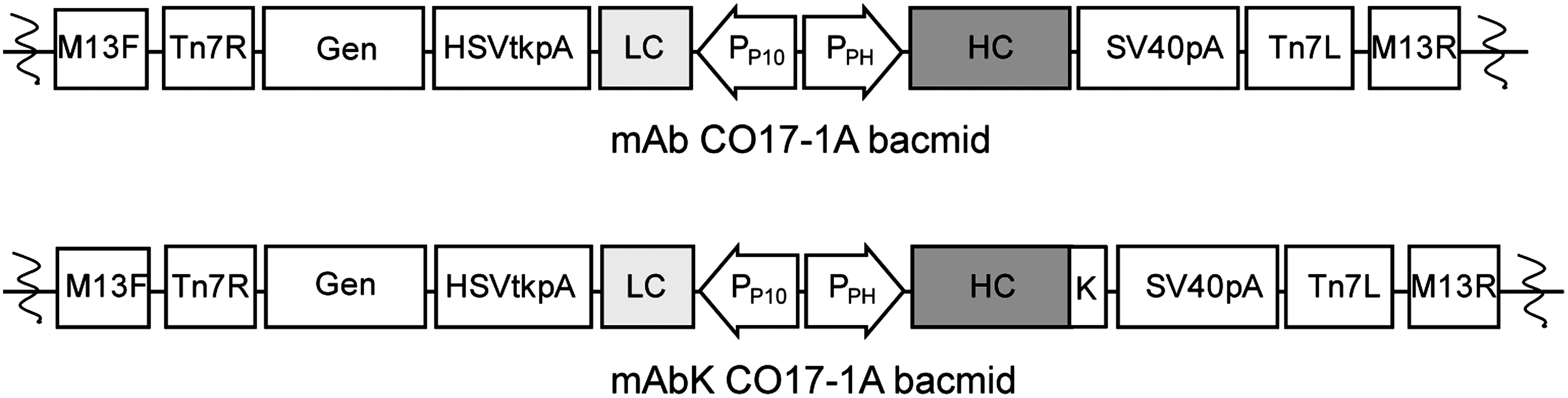
Arrangement of MAb CO17-1A with or without the KDEL gene expression cassette in a recombinant bacmid. LC and HC genes were placed codirectionally under control of the PP10 and PPH promoters, respectively. Tn7R and Tn7L permit site-specific transposition of the gene of interest into the baculovirus genome; M13F, M13 forward primer; HSVtkpA, Herpes simplex virus thymidine kinase polyadenylation signal; LC, light chain of MAb CO17-1A; PP10, p10 promoter; PPH, polyhedrin promoter; HC, heavy chain of MAb CO17-1A; K, KDEL ER retention signal; SV40pA, SV40 polyadenylation signal; M13R, M13 reverse primer.
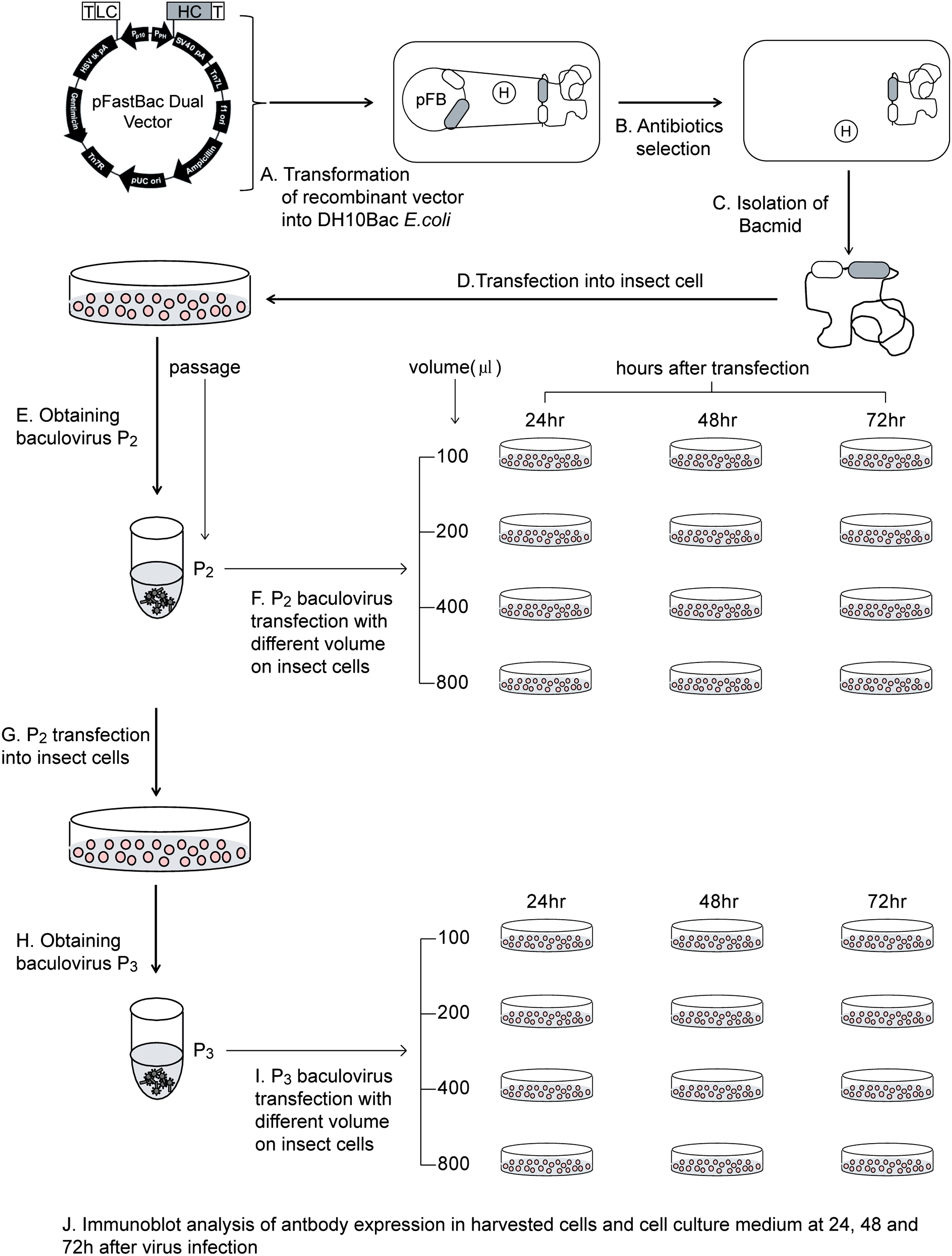
Schematic of MAb CO17-1A expression optimization in the baculovirus system. (
Insect cell culture and bacmid transfection
Spodoptera frugiperda (Sf9) insect cells were suspension cultured in T-25 flasks containing 5 mL of Sf-900 growth medium (Gibco BRL, Gaithersburg, MD) at an agitation rate of 50 rpm and a temperature of 27°C. Sf9 cells were transfected using Cellfectin reagent (Invitrogen) according to the manufacturer's instructions. Cells (9 × 105 cells/well) were seeded into a 6-well tissue culture plate (Nalge Nunc International, Rochester, NY) with 2 mL of growth medium containing antibiotics and incubated at 27°C for 1 h to attach the cells to the plate. CO17-1A bacmid DNA (1 μg) was mixed with 6 μL of Cellfectin lipid reagent in 200 μL of antibiotic-free Sf-900 growth medium and was incubated for 30 min at room temperature. Medium containing the transfection agent was removed 6 h after transfection and replaced with 2 mL of fresh growth medium, and the incubation was continued for 72 h at 27°C to generate P1 baculovirus.
Generation of P2 and P3 recombinant baculoviruses
The P1 baculovirus-containing supernatant was collected from the transfected cells after 72 h of culturing by centrifugation. Sf9 cells (2 × 106 cells/well) were attached to a 6-well plate in Sf-900 II growth medium for 1 h at room temperature, and then 100 μL of P1 viral stock were added to each well and incubated for 48 h at 27°C. Cell supernatant containing P2 baculovirus was collected by centrifugation. Subsequently, Sf9 cells (2 × 106 cells/well) were infected with P2 virus (0.5 mL) in a 20 mL culture in an Erlenmeyer flask for 72 h at 27°C to produce P3 virus.
Optimization of baculovirus transfection and cell harvest time
Sf9 cells (1.2 × 106 cells/well) 5 mL cultured in a T-25 flask were infected with different volumes of baculovirus (P2 and P3 viral stocks; 100, 200, 400, and 800 μL) (Fig. 2). Insect cells and culture medium were harvested at 24, 48, and 72 h after infection. The insect cells and the cell culture medium were separated by centrifugation, and the insect cells were lysed with lysis buffer (50 mM Tris [pH 8.0], 500 mM NaCl, 0.1% Triton 100) and broken by sonication. The harvested cells and culture medium were stored at −20°C until analysis.
Semi-quantitative immunoblot analysis
The cell lyaste and medium were boiled for 5 min in 5 × protein sample loading buffer (60 mM Tris-HCl, 25% glycerol, 2% SDS, 5% 2-mercaptoethanol, 0.1% bromophenol blue). Total soluble proteins were loaded into each lane of a 12.5% SDS-PAGE gel and electrophoresed and then electrotransferred to nitrocellulose membranes (Millipore, Billerica, MA).
Membranes were first incubated in blocking solution [3% (w/v) skim milk (Fluka, Buchs, Switzerland) in 1 × PBS], and then with goat anti-murine Fcγand F(ab')2 fragment specific antibodies conjugated to horseradish peroxidase (1:3000 dilution; Jackson ImmunoResearch Labs, West Grove, PA) to detect the HC and LC, respectively. Protein bands were visualized using a chemiluminescence substrate (SuperSignal, Pierce, Rockford, IL); then the membrane was exposed to X-ray film (Fuji, Tokyo, Japan). Mammalian-derived MAb CO17-1A (100 ng/20 μL) was used as a positive control. Antibody levels were analyzed by densitometry using Image Quant v2005 software (GE Healthcare, Chalfont St. Giles, UK).
Results and Discussion
We investigated several variables that affect expression of the anti-colorectal cancer monoclonal antibody in Sf9 cells: baculovirus passage, virus inoculum volume, harvest time after virus transfection, harvest sample (cell culture medium or cell lysate), and ER localization of the MAb by KDEL (an ER retention signal peptide). In this study, the parameters mentioned above were investigated to obtain the highest expression of MAb CO17-1A using BES.
The LC and HC genes were cloned into pFastBac dual vector under the control of the PP10 and PPH promoters, respectively. To investigate the effect of KDEL on MAb CO17-1A expression in the baculovirus-insect cell system, the HC gene was fused to KDEL (Fig. 1). The recombinant pFastBac dual vector carrying the LC and HC/HCK genes of MAb CO17-1A were transformed into DH10Bac E. coli cells to yield the MAb CO17-1A and CO17-1A K bacmids (Fig. 1). Successful insertion of the MAb CO17-1A and CO17-1A K genes into the bacmids was confirmed by PCR analysis (data not shown). Both bacmids were stored at −20°C until further study.
The optimal infection period, an important factor for the acquisition of a high yield of recombinant protein, was investigated, and the harvest time after virus transfection was varied––24, 48, and 72 h (Fig. 2). At 24 h after virus infection, none of the insect cells infected with the MAb CO17-1A K and CO17-1A baculoviruses P2 and P3, and 100 and 200 μL inoculum volume] showed any protein signal on Western blot analysis, except for cells infected with the P3 MAb CO17-1A K baculovirus (400 and 800 μL). In these samples, trace protein bands (50 kDa) were just barely detected in the cell lysate (data not shown, see Fig. 2). No protein band was observed in the culture medium from cells infected with P3 virus. It seems that 24 h infection should not be considered an optimal infection period regardless of any conditions or parameters.
At 48 h after infection, HC protein bands were detected in insect cell cultures infected with both the MAb CO17-1A and CO17-1A K baculoviruses (Fig. 3). In general, cell lysate (CL) samples showed stronger HC protein bands (50 kDa) than the cell culture medium (CM). With both the P2 and P3 baculoviruses, the HC expression level was higher in the 400 μL infection. In addition, the HC expression level of MAb CO17-1A was significantly higher than that of MAb CO17-1A K, and P3 baculovirus-infected cells showed a higher HC expression level than those infected with P2 baculovirus. In the CL samples from P3 MAb CO17-1A virus-infected insect cells, three major bands were detected on the Western blot (Fig. 3B). In general, the protein band density was nearly two times stronger in MAb CO17-1A baculovirus-infected cells than in MAb CO17-1A K virus-infected cells. However, hydrolytically degraded protein-like bands (25 and 17.5 kDa) were detected in the CL (Fig. 3A and 3B; black and open arrows, respectively). At 48 h after virus infection, 400 μL was the optimal baculovirus inoculum volume. Considering virus passage, the cells infected with the P3 baculovirus showed better expression than those infected with the P2 virus. Regarding the harvest sample, better expression was observed in the CL than in the CM. Expression of the MAb with the HC fused to KDEL was not higher than the MAb without KDEL, indicating that ER retention does not enhance the MAb production in either the CL or the CM.

Variable expression of MAb CO17-1A and MAb CO17-1A K in insect cell lysate and cell culture medium 48 h after virus infection. (
At 72 h after P2 virus infection, MAb CO17-1A K virus-infected cells showed increasing protein bands with increasing volume of virus inoculum in the CL until 400 μL (Fig. 4A). However, in the cell culture medium, where the MAb CO17-1A K virus-infected insect cells grew, no band was detected (Fig. 4A). Consistent with the results observed 48 h after infection, MAb CO17-1A virus-infected cells showed higher-density protein bands in the CL than in the CM. At 72 h after P3 virus infection, in both MAb CO17-1A K and CO17-1A MAb virus-infected cells, the CL of the 400 μL baculovirus infection showed the highest band density (Fig. 4B). However, no band was detected in the CM of MAb CO17-1A K virus infected-cells (Fig. 4B, CM). The HC protein band density in both the CL and CM in the P3 MAb virus-infected cells was generally stronger than that of the P2 MAb virus-infected cells. The HC protein band density was not significantly different in insect cells infected with the P2 and P3 MAb CO17-1A K virus (Fig. 4A,B). The MAb band was not detected in the CM of cells infected with the MAb CO17-1A K virus (Fig. 4).

Variable expression of MAb CO17-1A and MAb CO17-1A K in insect cell lysate and cell culture medium 72 h after virus infection. (
In general, higher HC protein density was observed at 72 h after virus infection than at 48 h. To compare the expression of MAb CO17-1A under all conditions mentioned, a 3D graph was constructed with Y = HC expression level, X = volume of infecting virus, and Z = virus passage (P2 or P3), ER retention signal (MAb or MAb K), and harvest samples (CM or CL) (Fig. 5). There was no expression signal in the cell culture medium from cells infected with the virus carrying the gene encoding MAb HC fused to KDEL (MAb CO17-1A K); however, in the lysate of cells infected with the P2 virus carrying the gene encoding MAb HC with KDEL (MAb CO17-1A K), the highest protein level (110 ng/μL) was expressed at an infection volume of 400 μL. In the cell lysate harvested from insect cells infected with P3 virus carrying the gene encoding MAb CO17-1A K, the highest MAb expression level (170 ng/μL) was observed at 72 h after infection.

3D visualization of HC protein expression Y = HC expression level affected by X = the volume of the infecting virus and Z = virus passage (P2 or P3), ER retention signal (MAb or MAb K), and harvest samples (CM or CL).
In the cell culture medium, proteinase secreted into the medium from cells might be involved in protein hydrolysis or degradation.(20) Indeed, hydrolytically degraded protein-like bands (25 and 17.5 kDa) were detected in the CL from cells infected with MAb CO17-1A virus; however, no degraded protein bands were detected in the CL from cells infected with MAb CO17-1A K virus (Figs. 3 and 4). These results indicate that localization of the protein in the ER can prevent protein degradation.
Therefore, to obtain MAb CO17-1A without any protein degradation, using MAb CO17-1A K, which is retained in the ER by KDEL, is a strategy that should be considered for better accumulation resulting in high production. Since the ER compartment has very low hydrolytic activity and is very plastic,(21) the ER is considered a good storage compartment that can tolerate unusually high protein accumulation without any physiological side effects on growth.(22) However, as shown in this study, the MAb CO17-1A with KDEL was observed in the CL but not in the CM; therefore, the ER-retained proteins were barely secreted from the cells. To obtain MAb CO17-1A from the cell culture medium without the cell lysis process, which might be easier than MAb purification from cell lysate, the MAb should not be retained in a subcellular compartment like the ER. To determine whether the proteins are localized in the ER or secreted through the outer membrane, the glycan structure on the protein expressed in insect cells should be considered. For instance, in rabies MAb,(16) regardless of purification process, the protein should be localized in ER and has high mannose-type glycan structure.
In this study, the optimal volume of virus inoculum was 400 μL. The cells infected with P3 virus showed a higher expression level than those infected with P2 virus. The optimal infection time for MAb CO17-1A expression was 72 h. Taken together, the best expression conditions were found in the lysates from cells infected for 72 h with a 400 μL baculovirus inoculum of MAb CO17-1A carrying expression cassettes without KDEL. Our data suggest that several parameters for the expression of the anti-colorectal cancer MAb CO17-1A can be optimized in a baculovirus expression system without the use of a commercial titration kit.
Footnotes
Acknowledgments
The authors wish to thank Dr. Hilary Koprowski (Biotechnology Foundation, Thomas Jefferson University, Philadelphia, PA) for suggestions and expertise. This study was supported by a grant from the National Research Foundation of Korea funded by the Korean Government (MEST) (NRF-2010-0024259, NRF-2010-0027694).
Author Disclosure Statement
The authors have no financial interests to disclose.