
Abstract
The conserved domain of bacteria-derived flagellin coupling Toll-like receptor 5 (TLR5) activates NF-κB and MAPK signaling transductions, which subsequently regulate the transcription and expression of genes encoding immune mediators. However, whether the flagellin binding monoclonal antibody (MAb) obstructs TLR5-associated signaling is unclear. Here we report on the production and characterization of MAb 5G10 that specifically recognizes flagellin. The MAb 5G10 was produced by the hybridization of mouse myeloma cell SP2/0 with splenocyte from a flagellin immunized BALB/c mouse. We observed that deletion of the conserved amino acid residues 89-96 made flagellin lose its capacity for binding 5G10. Additionally, MAb 5G10 remarkably suppressed the expression of cytokine IL8 of Caco-2 cell by blocking the flagellin-TLR5 signaling. Furthermore, this MAb would be useful for cytosolic localization of flagellin and would facilitate the elucidation of the physiological function of specific pathogen-associated molecular patterns.
Introduction
Here we report on the production and characterization of a monoclonal antibody that specifically recognizes the amino acid residues 89-96 of the bacteria-derived flagellin. This monoclonal antibody would be useful for localization studies for certain flagellin and bacteria, and should aid in the elucidation of the physiological function of this specific pathogen-associated molecular pattern (PAMP).
Materials and Methods
Preparation of monoclonal antibodies against flagellin
The preparation of His-tagged flagellin (FliC) or 89-96 animo acid-deleted flagellin (FliCΔ89-96) and MAbs against the flagellin were generated as previously described.(7,8) In brief, 5-week-old female SPF BALB/c mice were immunized subcutaneously with 100 μg of the recombinant FliC at 2-week intervals. Four weeks after the last booster and 3 days before cell fusion, the mice were boosted with 200 μg of the FliC. Three days later, mice splenocytes were harvested and fused with SP2/0 using 50% polyethyleneglycol (Sigma-Aldrich, St. Louis, MO). Hybridoma culture supernatants were screened using ELISA; meanwhile the FliCΔ89-96 served as a negative selection control. The positive hybridoma cells were cloned by a limiting dilution and the stable hybridoma clones were injected into liquid paraffin-pretreated abdominal cavities of BALB/c mice. Subsequently, the MAbs were harvested and purified from the seroperitoneum with an antibody purification kit according to the manufacturer's specifications (NAb™ Protein A/G Spin Kit, Thermo Scientific, Pittsburgh, PA).
ELISA
Recombinant His-fused FliC and FliCΔ89-96 proteins (5 μg/mL) in coating buffer (40 mM Na2CO3, 60 mM NaHCO3 [pH 9.6]) was adsorbed to the surface of 96-well flexible microplates (Greiner Bio-one, Frickenhausen, Germany) at 37°C overnight. The hybridoma supernatants were respectively incubated in the FliC and FliCΔ89-96 coated microplates for 1 h at room temperature. After washing three times with PBS-T, the plates were incubated for 45 min at room temperature with alkaline phosphatase-conjugated anti-mouse IgG antibody. After washing with PBS-T seven times, immunoreactivity was visualized by means of a pNPP phosphatase substrate system (KPL, Gaithersburg, MD).
Immunoblotting
The purified proteins FliC and FliCΔ89-96 were separated by 10% SDS-PAGE and then electrophoretically transferred to nitro-cellulose transfer membrane (GE Healthcare, Little Chalfont, United Kingdom). The membrane was blocked for 1 h at room temperature with blocking solutions containing 1% BSA in TBS (20 mM Tris-HCl [pH 7.5], 150 mM NaCl) and then incubated overnight with hybridoma supernatants. After washing with T-TBS, the membrane was incubated for 30 min with HRP-conjugated anti-mouse IgG (Jackson ImmunoResearch Laboratories, West Grove, PA). After washing with T-TBS, the membrane was developed by treatment with ECL Western Blotting Detection Reagents (GE Healthcare/Amersham, Piscataway, NJ).
IL8 releasing assay
One×105 Caco-2 cells/well was seeded in the 24-well plate overnight. 0.01, 0.1, 1, 10, 100, or 1000 ng of the FliC were incubated with or without 1000 ng MAb 5G10 at 37°C for 1 h, then the mixtures were respectively added to the Caco-2 cell culturing for 24 h at 37°C under 5% CO2 atmosphere. The indicated concentration of FliCΔ89-96 was used as control. The cytokines released from differently treated Caco-2 were measured by ELISA kits (eBioscience, San Diego, CA) according to the manufacturer's guidelines. The concentrations of cytokines in the samples were calculated from the standard curves.
Results and Discussion
The 89-96 amino acids of the flagellin were deleted by overlapping polymerase chain reaction (PCR) (Fig. 1). The recombinant proteins 6×His-fused FliC and FliCΔ89-96 (about 53 kDa) were initially expressed and purified from bacterial source. To obtain specific monoclonal antibodies (MAbs) against the 8 amino acid residues (RVRELAVQ) of the FliC, the recombinant protein FliCΔ89-96 served as a negative screening protein. After three rounds of selection, we obtained an MAb, named 5G10, that reacts with FliC, but not with FliCΔ89-96 in ELISA and Western blot analysis (Fig. 2A). Subsequently, different recombinant proteins, such as FliCΔ190-278, FliCΔ220-320, FliCΔ180-400, and FliCΔ297-471, were used to exclude the possible cross-reactivity of MAb 5G10. The results showed that there was no cross-reactivity with the other regions of the flagellin (data not shown).
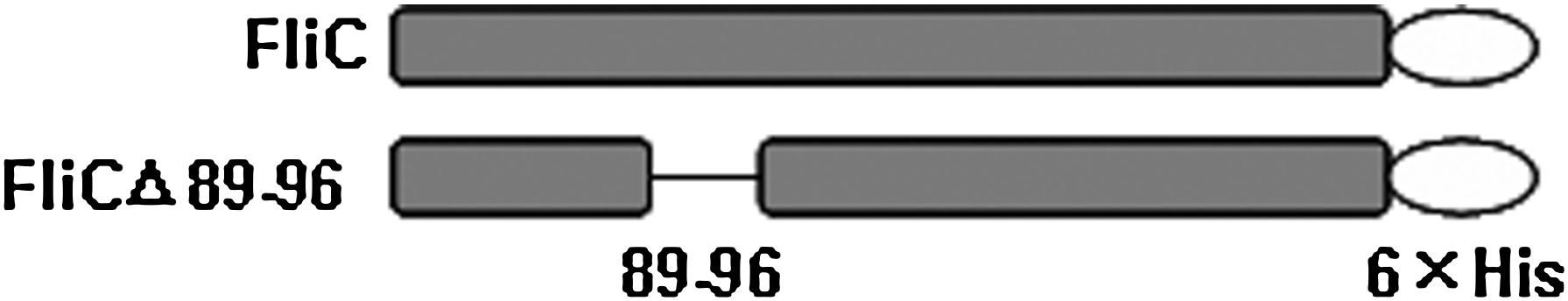
Schematic of bacterial flagellin proteins. FliC, full-length flagellin; FliCΔ89-96, 89-96 amino acid-deleted flagellin.

(
As a typical PAMP, the flagellin recognizes the 552-561 amino acids (EILDISRNQL) of TLR5,(9) the key component of the immune system that detects microbial infection and triggers antimicrobial host defense responses including the production of immune mediators.(10) In our previous study, a panel of flagellin mutants (FliCΔ190-278, FliCΔ220-320, and FliCΔ180-400) have been shown to posses the capability of stimulating the cytokine expression, such as IL-8, MCP-1, and TNF-α.(7) However, whether the flagellin-binding MAb interferes with the expression of inflammatory factors is unknown. Here we observed that MAb 5G10 significantly suppressed the flagellin-induced IL8 production by intercepting the flagellin-TLR5 interaction (Fig. 2B). The purified 5G10 alone did not induce IL8 production.
Meanwhile we obtained the recombinant flagellin derived from commensal bacteria K-12 (designated KF) to examine the cross-reactivity of MAb 5G10 with different bacteria-derived flagellins. Consequently, the MAb 5G10 showed significant cross-reactivity with the KF (data not shown). The following sequence alignment analyses show that RVRELAVQ and RVRELTVQ are from pathogenic bacteria Salmonella and nonpathogenic bacteria K-12, respectively, and both of them keep high homology in the crucial conserved amino acid sequences. Lodes and colleagues identified that the commensal bacterial flagellin could be the dominant antigen, which contributed to the pathogenesis of inflammatory bowel disease (IBD).(11) Therefore, it implied that MAb 5G10 could be used as a potential material for clinical detection of pathogenic and nonpathogenic bacteria-derived flagellin.
In addition, cytosonic recognition of flagellin by sensor Naip5 and Ipaf/NLRC4 is recently becoming a research hotpoint for identification of pathogenic and nonpathogenic bacteria by host.(4) This MAb could contribute to the localization of intracellular flagellin and elucidation of its physiological function.
Footnotes
Acknowledgments
This work was financially supported by grants from the Ministry of Science and Technology of China (Key R&D Program, no. 2007BAI28B04; National S&T Major Project on Major Infectious Diseases, nos. 2008ZX10001-010 and 2012ZX10001-008).
Author Disclosure Statement
The authors have no financial interests to disclose.