
Abstract
IL-18 is a pro-inflammatory cytokine that is produced from T cells and NK cells. IL-18 has been implicated in the pathogenesis of various inflammatory and cardiovascular diseases. IL-18 binding protein (IL-18BP) is a natural inhibitor of IL-18 that possesses higher affinity to IL-18 than that of the IL-18 receptor alpha chain on the cell surface. Human isoform a and c among four isoforms of IL-18BPs have an inhibitory effect on IL-18-induced cytokines whereas mouse IL-18BP isoforms exist only in two isoforms: c and d. Fc-fusion protein is a molecule in which the immunoglobulin Fc is fused genetically to a protein of interest, such as an extracellular domain of a receptor, ligand, or enzyme. In this study, we expressed and purified human Fc-IL-18BPa and c isoforms from CHO-DG44 cells and their biological activities were compared to each other. This is the first time that expressed recombinant human Fc-IL-18BPc has been examined for its biological activity on IL-18-induced IFNγ in human PBMC and IL-6 in A549/IL-18Rβ.
Introduction
IL-18 has been implicated in the pathogenesis of various inflammatory disorders and processes, including diabetes mellitus, atherosclerosis, sepsis, chronic liver disease, and several autoimmune diseases, including rheumatoid arthritis, multiple sclerosis, and Crohn's disease.(6–13) Cells expressing IL-18 have been reported within neuroinflammatory lesions in human multiple sclerosis,(6) and both IL-18 and caspase-1 mRNA levels were considerably elevated in Lewis rats with experimental autoimmune encephalomyelitis, which is a mouse model for multiple sclerosis.(9) In animal models of rheumatoid arthritis, administration of IL-18 provoked the course of the disease while a combination of IL-18 with a neutralizing anti-IL-18 antibody significantly reduced joint swelling.(14)
It has been shown that IL-18 can be a predictive biomarker for human death due to cardiovascular disease.(15,16) The plasma IL-18 level is correlated with a number of risk factors for cardiovascular disease independent of C-reactive protein and IL-6, including LDL cholesterol and triglyceride levels.(8) IL-18 has also been implicated in the pathogenesis of atherosclerosis. As circulating levels of IL-18 are higher in subjects with increased carotid- intima-media thickness,(17) increased IL-18 expression has been related with unstable plaque.(18,19)
IL-18 binding protein (IL-18BP) is a natural antagonist for IL-18.(13) In fact, the balance between circulating IL-18 and IL-18BP affects the severity of some inflammatory diseases.(20) Transgenic mice overexpressing human IL-18BPa (IL-18BPa Tg) produce high levels of bioactive IL-18BPa in the circulation, providing protection against inflammatory stimuli.(21) Although IL-18BP slightly resembles the IL-1 and IL-18 membrane receptors, it is not a variant of the extracellular ligand-binding portion of these receptors. IL-18BP is a distinct gene composed of a single immunoglobulin domain.(13,22)
Interestingly, several poxviruses encode IL-18BP orthologues as virulence factors that contribute to immune escape by suppressing antiviral immune responses of the host.(23–26) These viral IL-18BP orthologues are evidence that IL-18BP has a pivotal role in viral infection as a mediator of cytotoxic immune responses. Although IL-18BP is produced constitutively, its expression is noticeably increased by IFNγ through two transcription factors, IFN regulatory factor 1 (IRF1) and CCAAT/enhancer binding protein beta (C/EBPβ). In addition, these provide negative feedback.(27–29)
There are several reports that IL-18BP can prevent or attenuate the development of disease in animal models(30,31); however the secreted amount of IL-18BP induced by IFNγ seems insufficient to defeat the quantity of IL-18 in circulation under certain pathological circumstances.(32) Four human isoforms of IL-18BP have been characterized (isoforms a, b, c, and d) but only isoforms IL-18BPa and IL-18BPc have affinity to IL-18 and down-regulate its function in vitro.(33) Similar to human IL-18BP, murine IL-18BPc and murine IL-18BPd neutralize the IL-18 activity at equal-molar concentrations.(33)
Fc fusion proteins are molecules in which the immunoglobulin Fc is fused genetically to a protein of interest. Fc fusion proteins have some antibody-like properties such as long serum half-life and easy expression and purification, making them an attractive platform for therapeutic drugs.(34–36) An Fc fusion-based drug, such as Enbrel,® is currently on the market, and many more are in different stages of clinical trials, demonstrating that Fc fusion proteins have become an alternative to monoclonal antibody therapeutics. In this study, we used pCAG.neo-hIgG1-Fc vector with a strong chicken beta-actin promoter and expressed recombinant Fc-IL-18BPa and c purified for experimental use.
Materials and Methods
PCR and molecular cloning
Human IL-18BPa and IL-18BPc cDNA was amplified from vectors previously constructed(13) with the common forward primer containing XbaI recognition site and Kozak consensus sequence (5′-ACCATCTAGAGCCACCATGAGACACAACT GGAC-3′) and MluI site containing reverse primer for IL-18BPa (5′ATTTACGCGTACCCTGCT GCTGTGGACT-3′) or IL-18BPc (5′-ATTTACGCGTCAGGCTGCTCTGGCAGAG-3′). PCR reaction was performed as described above. Inserts were digested with XbaI and MluI and ligated to pCAG.neo-hIgG1-Fc cut with the same restriction enzymes. The DNA sequence of cloned vectors was also analyzed and the vectors were named Fc-IL-18BPa and Fc-IL-18BPc.
Cell culture
The dihydrofolate reductase (DHFR)-deficient Chinese hamster ovary (CHO) cell line, DG44, was used for stable expression of recombinant Fc-IL-18BP chimeric proteins. The CHO-DG44 cells were grown at 5% CO2, 37°C in DMEM/F12 (Invitrogen, Carlsbad, CA) supplemented with hypoxanthine (10 mg/L), thymidine (10 mg/L), glycine (50 mg/L), glutamine (587 mg/L), glucose (4.5 mg/L), 10% FBS, and antibiotics-antimycotics (Invitrogen).
Production of chimeric recombinant protein
hFc-IL-18BP plasmids were co-transfected with pSV-DHFR vector (ATCC, Manassas, VA) into DHFR-deficient CHO cell line (DG44), and stably transformed cell lines were selected in a medium containing G418 (500 μg/mL) and subsequently subjected to methotrexate (MTX) selection for gene amplification, as described previously.(37) The CHO cells secreting Fc-IL-18BP were grown in serum-free medium (CHO-SSFMII, Invitrogen), and the culture supernatant was subjected to affinity chromatography on protein A-sepharose 4B column (GE Healthcare, Piscataway, NJ), as described previously.(37)
Immunoprecipitation
Recombinant human IL-18 (100 ng, YbdYbiotech, Seoul, Korea) was incubated with freshly purified recombinant Fc-IL-18BPa or Fc-IL-18BPc (500 ng) in 50 μL volume of PBS for 1 h on ice, and then the mixture was immunoprecipitated with protein A-agarose beads for 4 h at 4°C. The precipitate was subjected to SDS-PAGE for Western blot analysis with rabbit anti-human IL-18 antibody (YbdYbiotech).
Bioassay and ELISA
Human PBMC was isolated as previously described.(38) 1.0×106 PBMCs were seeded in a 96-well plate containing 10% FBS. IL-12 (2 ng/mL) and IL-18 (20 ng/mL) were mixed with Fc-IL-18BPa or Fc-IL-18BPc (2 μg/mL) for 30 min at room temperature and added to PBMCs for 24 h. The culture supernatant was collected for measurement of IFNγ by ELISA, as described in the manufacturer's manual (R&D Systems, Minneapolis, MN).
A549-IL-18Rβ cells were cultured as previously described.(38) 2.5×104 cells were seeded in a 96-well plate containing 10% FBS. IL-18 (50 ng/mL) was mixed with Fc-IL-18BPa or Fc-IL-18BPc (2 μg/mL) for 30 min at room temperature and added to A549-IL-18Rβ cells overnight. The culture supernatant was harvested for measurement of IL-6 by ELISA (R&D Systems).
Statistical analysis
Data are expressed as means±SEM. Statistical significance of differences was analyzed by the unpaired, two-tailed Student's t test. Values of p<0.05 were considered statistically significant.
Results
Expression and purification of recombinant human Fc-IL-18BP
The schematic structures of Fc-IL-18BPa and Fc-IL-18BPc are shown in Figure 1. The recombinant human Fc-IL-18BPa and c were purified through protein A-agarose columns. Eluted fractions of recombinant Fc-IL-18BPa and c were subjected to SDS-PAGE and silver-staining in non-reduced or reduced conditions. The molecular size of the non-reduced protein was approximately 130 kDa while the reduced protein was 55 kDa (Fig. 2A). Western blotting against IL-18BP and human IgG-Fc confirmed that those bands are Fc-IL-18BPa (Fig. 2B). In reduced conditions, the affinity-purified IL-18BPa and c appeared as a single band with silver staining and Western blot analysis.

Structures of Fc-IL-18BPa and Fc-IL-18BPc. Fc-IL-18BP proteins are expected to be expressed in homodimeric form coupled by disulfide bonds. Both Fc-IL-18BPa (
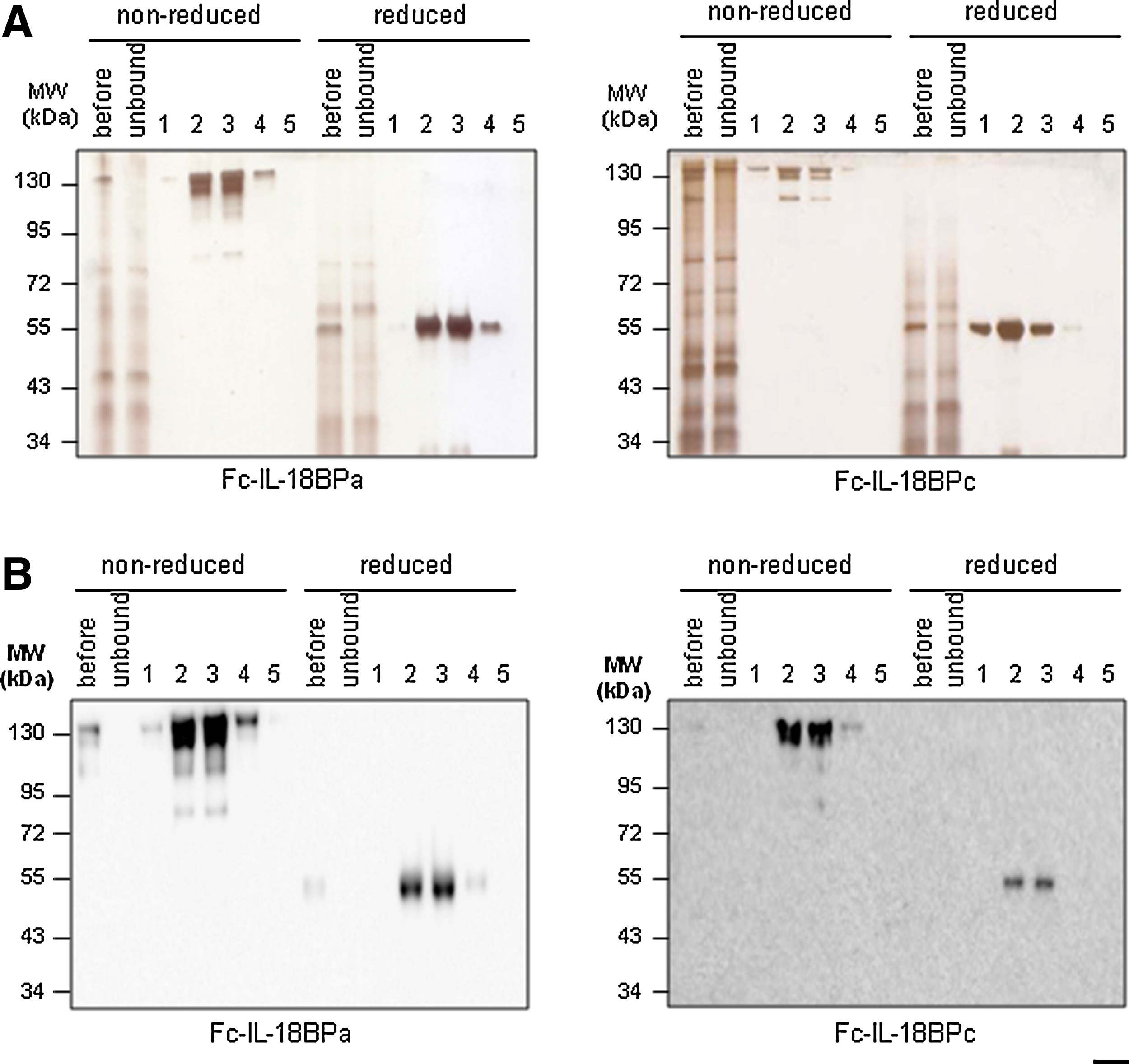
Purification of recombinant human Fc-IL-18BPs. (
Fc-IL-18BP binds to IL-18BP
We performed immunoprecipitation of recombinant IL-18 with purified Fc-IL-18BPa or Fc-IL-18BPc to confirm whether purified proteins bind to IL-18 properly. Both Fc-IL-18BPa and Fc-IL-18BPc were precipitated with recombinant IL-18. Recombinant Fc-IL-18BPa and Fc-IL-18BPc protein was bound to mature IL-18 (20 kDa) and precursor IL-18 (30 kDa) that was not fully cleaved with Tobacco Etch virus (TEV) enzyme (Fig. 3A). We therefore repeated the experiment with precursor IL-18, and the result showed that precursor IL-18 was also precipitated with Fc-IL-18BPa or Fc-IL-18BPc (Fig. 3B).

IL-18 was specifically immunoprecipitated by Fc-IL-18BP. Immunoprecipitation of IL-18 was performed with recombinant Fc-IL-18BPa or Fc-IL-18BPc. Both Fc-IL-18BPa and Fc-IL-18BPc precipitated precursor and mature IL-18 (indicated by arrows) compared to negative control in the first lane. (
Fc-IL-18BP decreases the activity of IL-18
IL-18 induces IFNγ from human PBMCs in combination with IL-12. Therefore, we stimulated human PBMCs with IL-12 and IL-18 in the presence or absence of recombinant Fc-IL-18BPa and c. IFNγ was not induced when IL-12 was solely treated; however it was highly induced when IL-12 was treated in combination with IL-18. Fc-IL-18BPa and c sufficiently suppressed IFNγ production in human PBMCs. The inhibitory effect of Fc-IL-18BPa was more significant than Fc-IL-18BPc (Fig. 4A). The combination of Fc-IL-18BPa and IL-18BPc also decreased IL-12 and IL-18 induced secretion of IFNγ (data not shown). We further confirmed the activity of Fc-IL-18BPs with A549-IL-18Rβ cells. IL-18 was treated on A549-IL-18Rβ cells with or without Fc-IL-18BPs. The A549-IL-18Rβ cells treated with IL-18 induced a high level of IL-6; however the induced level of IL-6 was prominently blocked in the presence of Fc-IL-18BPa or Fc-IL-18BPc (Fig. 4B).
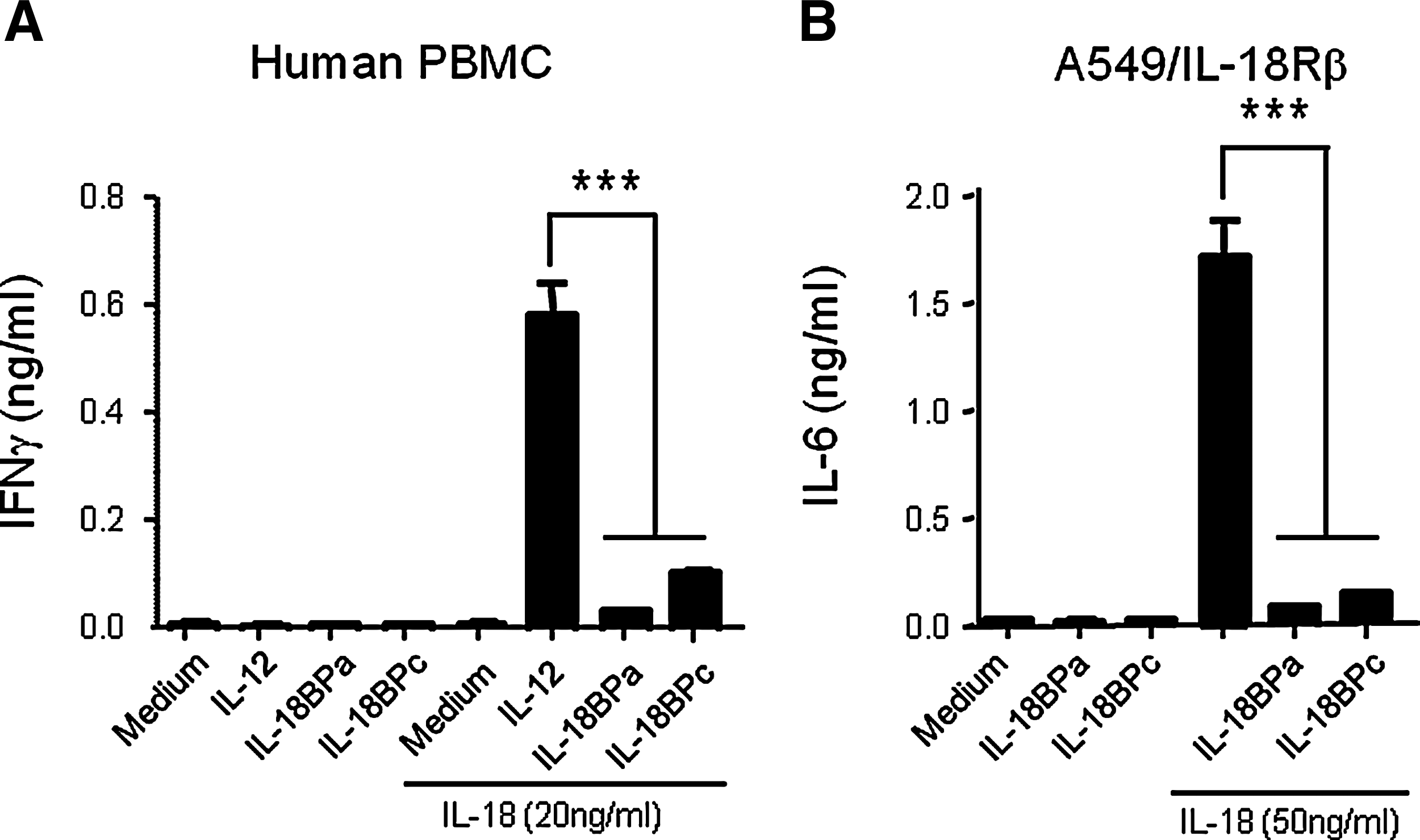
Fc-IL-18BP decreases the biological activity of IL-18. (
Discussion
With a single Ig domain, IL-18BP resembles an extracellular segment of the third domain of the IL-1 family receptor, which is a significant domain for IL-18 receptor alpha chain binding to its ligand.(13,22) However, IL-18BP is a distinct protein from the IL-1 and IL-18 receptor families and locates in chromosome 11q13 at the inverted position of the nuclear mitotic apparatus protein-1.(13) The human IL-18BP genomic DNA encodes at least four different isoforms derived from alternative splicing isolated from several human cDNA libraries. Two murine IL-18BP isoforms were found and murine IL-18BPc and d are similar to human IL-18BPc and a, respectively.(33)
IL-18BP is a unique soluble protein, lacking a transmembrane domain, and it specifically binds to IL-18 with an extraordinarily high affinity (100 picomolar range).(33) IL-18BP neutralizes the bioactivity of IL-18 with an equimolar ratio and shares high homology with several poxviral proteins, which indicates the importance of this molecule in resistance to viral infections.(13,23–26) IL-18 has been reported to be related to various diseases, including diabetes mellitus, atherosclerosis, sepsis, chronic liver disease, rheumatoid arthritis, multiple sclerosis, and Crohn's disease.(6–13) IL-18 is also related to cardiovascular diseases.(15,16) Fc-IL-18BP can be used in studies related to those diseases; furthermore this can be a substitutive or additive drug.
In this study, we expressed Fc-IL-18BPa and c isoforms and examined their biological activities on the inhibition of IL-18-induced cytokines. As described above, recombinant Fc-IL-18BP proteins obtain some antibody-like properties such as long serum half-life and easy expression and purification, making them attractive for use in different experimental disease models.(34–36)
Footnotes
Acknowledgments
This work was supported by Konkuk University.
Author Disclosure Statement
The authors have no financial interests to disclose.