
Abstract
PEX14 is an integral membrane protein essential for protein docking onto the peroxisomes and is a bi-functional protein capable of acting as a transcriptional co-repressor and a polypeptide transport modulator. Further studies showed that PEX14 is the sole peroxin that has a unique dual function in peroxisome formation and selective degradation. Its RNA transcripts are ubiquitously expressed; there is, however, no data on the expression profiles of PEX14 at the protein level due to a lack of MAbs suitable for immunohistochemical staining, thus hindering studies on its functions. In the present study, we generated one MAb that could be used in immunocytochemistry and immunohistochemistry and investigated PEX14 expression in normal and malignant human tissues. In contrast to the ubiquitous expression of its RNA transcripts, there is no PEX14 protein expression in normal human tissue, including liver, spleen, lung, rectum, brain, prostate, breast, ovary, and stomach. Protein expression of PEX14 was dramatically upregulated in some malignancies. Presented here are the first data on the expression profiles of PEX14 at the protein level, which will further our understanding of its functions.
Introduction
Considering the important function of PEX14, we cloned full-length PEX14 and expressed GST-PEX14 fusion protein. Successful immunization and cell fusion followed by screening using immunohistochemistry on sections made of PEX14 stable transfected cell lines resulted in one immunocytochemistry-positive MAb named FMU-PEX14. Consequent experiments showed that the MAb could be used in immunohistochemistry to investigate PEX14 expression in normal and malignant human tissues. In contrast to the ubiquitous expression of its RNA transcripts, there is no PEX14 protein expression in human normal tissue, including liver, spleen, lung, rectum, brain, prostate, breast, ovary, and stomach. Protein expression of PEX14 was dramatically upregulated in some malignancies such as carcinoma of lung, rectum, ovary, and esophagus. The unique expression profiles of PEX14 at the protein level would further our understanding of its functions.
Materials and Methods
Expression of recombinant PEX14, immunization, and production of hybridomas
Full-length wild-type PEX14 cDNA was subcloned into the pGEX-4T-1 vector and expressed as a glutathione S transferase (GST) fusion protein in BL21 Escherichia coli. Soluble protein was purified from bacterial cell lysates on glutathione S-Sepharose (Amersham Biosciences, Piscataway, NJ), washed three times in lysis buffer, and eluted with 20 mM glutathione. The protein was then dialyzed into phosphate-buffered saline (PBS) and used as antigen to immunize the mice.
Female BALB/c mice (8 weeks old) were immunized with 20 μg of GST-PEX14 in complete Freund's adjuvant by subcutaneous (s.c.) injection. Then immunizations were performed two more times with 20 μg of recombinant proteins mixed with incomplete Freund's adjuvant by s.c. injection at 4-week intervals. The titer of anti-GST-PEX14 antibodies in blood sera was monitored by ELISA. Subsequently, one immunized mouse was boosted and, three days later, splenocytes from the boosted mouse, and SP2/0 myeloma cells were fused by PEG (MW4000, Merck, Darmstadt, Germany). The positive hybrids were selected for by immunocytochemistry and then subcloned three times using limiting dilution method.
BALB/c mice were injected with 0.2 mL of pristane 7 days before being inoculated with 1×106 hybridoma cells. The ascitic fluid was collected and the fraction of immunoglobulins was precipitated with 40% ammonium sulphate and used for affinity purification by Protein A-Sepharose CL-4B (GE Healthcare, Piscataway, NJ) chromatography. The IgG fractions were pulled together and dialyzed in PBS (pH 7.4).
Establishment of PEX14 transfected stable cell lines
L929 cells were transfected with pCMV-4-PEX14 with transfection reagent Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. Briefly, L929 cells at 1×106 per well in 6-well plates were seeded and incubated for 12–24 h at 37°C in 5% CO2. The DNA-Lipofectamine 2000 complexes were added directly to the wells and mixed gently. Stable cell lines were established by G418 selection. Then the cells were collected, mixed with non-transfected L929 cells in 1:10 ratio, fixed by formalin for 8–10 h, embedded in paraffin, and made into cell blocks.
Immunocytochemistry and immunohistochemistry
Cell blocks were cut at 5 μm thicknesses and mounted on gelatin-coated slides. Endogenous peroxidase activity of slides of cell block and tissue arrays (Chaoying Biotech, Xi'an, China) was blocked with methanol containing 3% H2O2 for 10 min, and non-specific binding was blocked by normal goat serum (10%) for 30 min. Anti-PEX14 monoclonal antibody was added to the slices and incubated for 1 h at room temperature. After three washes in PBS, the slices were incubated with biotin-conjugated goat anti-mouse IgG (Promega, Madison, WI) for 1 h at room temperature followed by streptavidin-HRP complex incubation for 30 min. Antibody complexes were visualized by DAB chromogen and examined using light microscopy. Normal mouse IgG was applied as negative control.
Results
Generation of MAbs and application of MAbs in immunocytochemistry
Successful immunization and cell fusion followed by screening using immunohistochemistry on slices made of PEX14 stable transfected cell lines resulted in one immunocytochemistry-positive MAb named FMU-PEX14. The immunoglobulin isotype of this MAb was IgG1 (κ). As shown in Figure 1, FMU-PEX14 antibody recognized endogenous rat BAP31 in pCMV-4-PEX14 transfected L929 cells, which is visualized by strong DAB staining in the cytoplasm (Fig. 1A). In all instances, no reactivity could be detected in control MAbs (Fig. 1B).
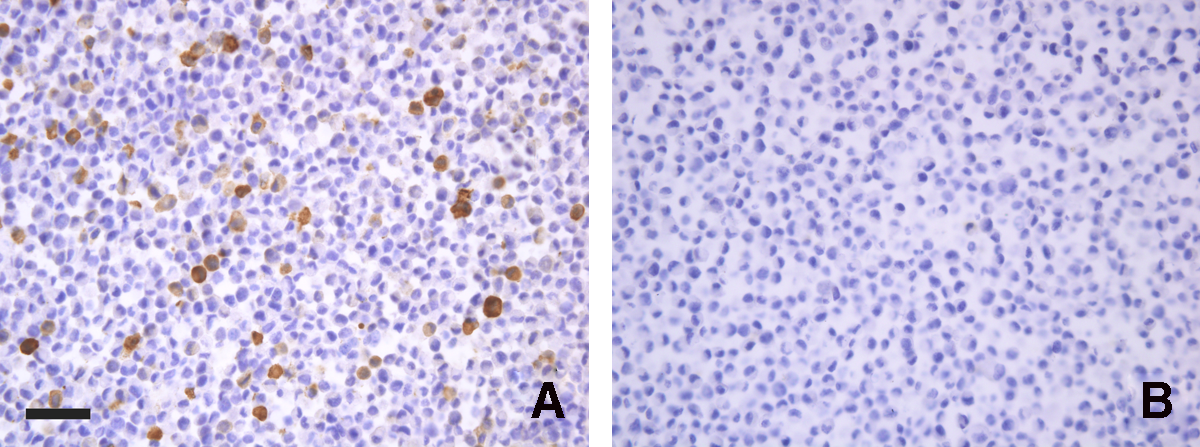
Immunoperoxidase staining of sections of PEX14 gene transfected cells by FMU-PEX14. Coding sequence of human PEX14 was inserted in pCMV-4 vector and was transfected into L929 cells. Stable cell lines were established by G418 selection. Then the cells were collected, mixed with non-transfected L929 cells in 1:10 ratio, fixed for 8–10 h, embedded in paraffin, and made into cell blocks. In all instances, no reactivity could be detected in control MAbs (
Localization of PEX14 immunoreactivity in human normal and malignant tissues
Our subsequent experiments showed that the FMU-PEX14 MAb could be used in immunohistochemistry, so we investigated PEX14 expression profiles in human normal and malignant tissues. Tissue arrays of normal human tissues were stained with newly developed FMU-PEX14 MAb. In contrast to the ubiquitous expression of its RNA transcripts, there is no PEX14 protein expression in liver, spleen, lung, rectum, brain, prostate, breast, ovary, and stomach detected by immunohistochemistry (Fig. 2). The mechanism of the discrepancy of the expression level between the mRNA and protein of PEX14 should be investigated more intensely.
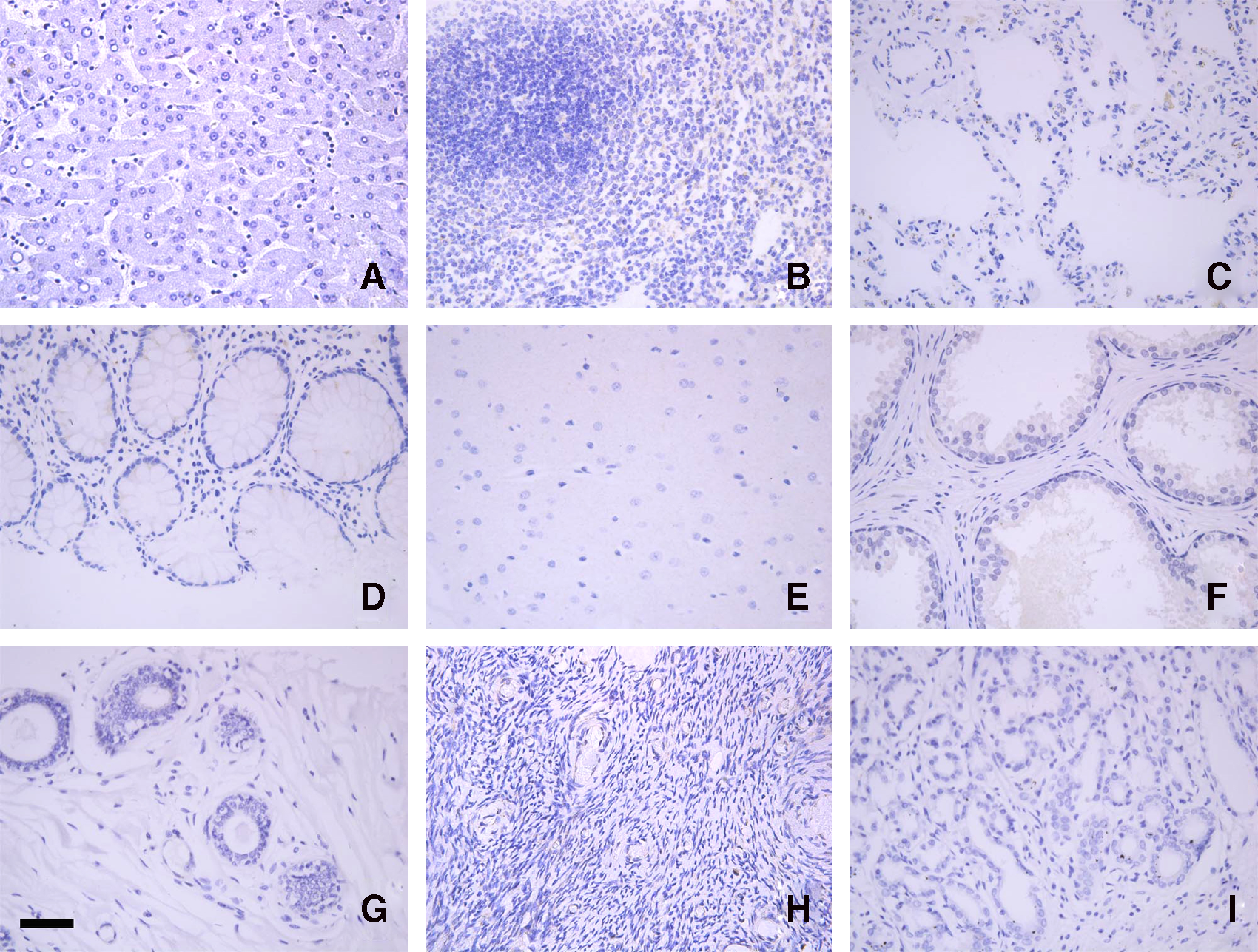
Expression profiles of PEX14 in normal human tissue. To assess the protein expression levels of PEX14 in normal human tissue, we employed immunohistochemical analysis using tissue arrays. In contrast with the ubiquitous expression of its mRNA, PEX14 protein immunoreactivity was not observed in liver (
To assess immunoreactivity of PEX14 in human malignant tissues, we examined immunoperoxidase-stained tissue arrays consisting of four human malignancies (carcinoma of lung, rectum, ovary, and esophagus). Strong brown reaction product, indicative of a high level of PEX14 protein expression, was in cancer cells of the lung, rectum, ovary, and esophagus but not in the mesenchymal cells (Fig. 3).
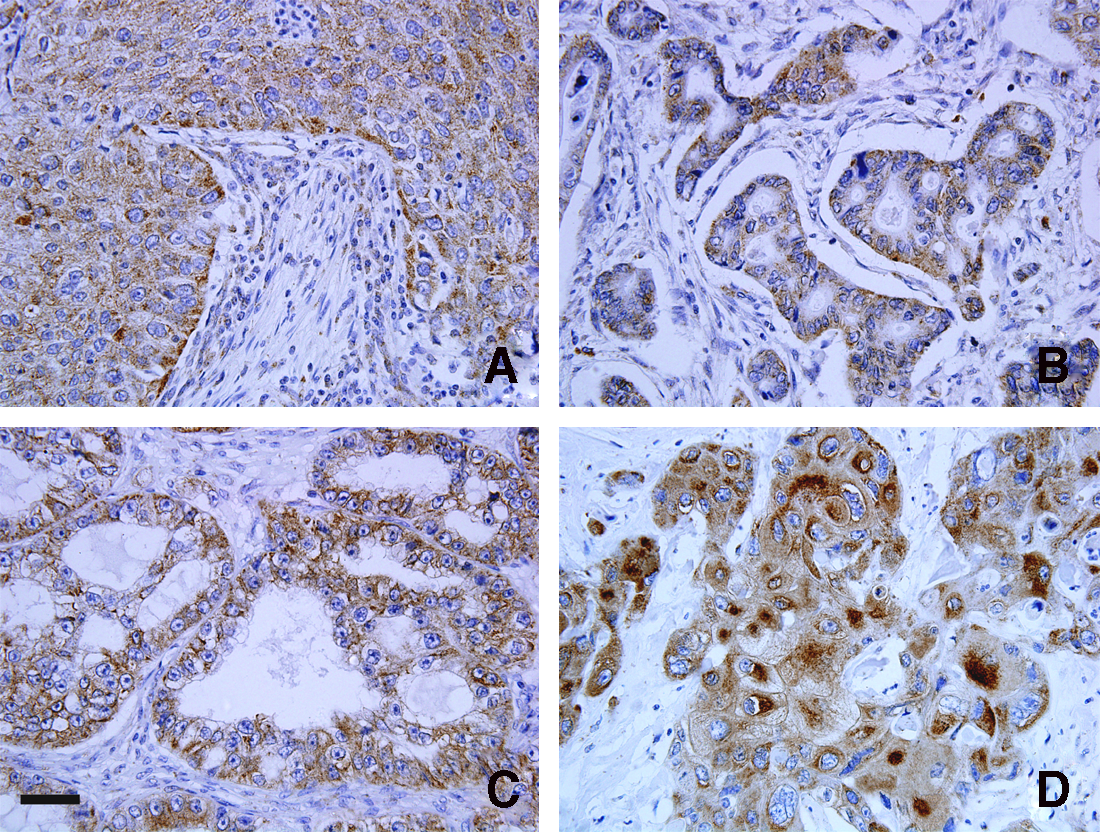
Expression profiles of PEX14 in human malignancies. Representative expression patterns of PEX14 in carcinomas of lung (
Discussion
Of all the cellular organelles, the biogenesis of peroxisomes is the least well understood.(5) The peroxisomal protein import machinery differs fundamentally from known translocons (endoplasmic reticulum, mitochondria, chloroplasts, bacteria) as it allows membrane passage of folded, even oligomerized proteins. The membrane-associated import receptor PEX5, together with its docking partner PEX14, forms a gated ion-conducting channel that can be opened to a diameter of about 9 nm by the cytosolic receptor-cargo complex.(6,7)
The peroxisomal docking complex is a key component of the import machinery for matrix proteins. PEX14 is thought to represent the initial docking site for the import receptors PEX5 and PEX7. Associated with this complex is a fraction of PEX13, another essential component of the import machinery.(8) Bharti and co-workers(9) have demonstrated that almost all known human peroxins involved in protein import were identified as constituents of the PEX14 complexes and tubulin was discovered to be the major PEX14-associated protein. The direct binding of PEX14 and tubulin was also demonstrated. Thus, peroxisomal remnants in PEX14-deficient cells lost their ability to move along microtubules, and human PEX14 is a multi-tasking protein that not only facilitates peroxisomal protein import but is also required for peroxisome motility by serving as a membrane anchor for microtubules.(9)
In the present study, we produced recombinant human PEX14 proteins and successfully generated a MAb against this protein. The identification of this newly generated human PEX14-specific MAb is focused on its application in immunocytochemistry and immunohistochemistry. Interestingly, we found that the expression of human PEX14 in the protein level is significantly upregulated in some tumor tissues when compared with normal tissues. Upregulation of the protein of PEX14 might be involved in the progress of tumor genesis; the unique expression profiles of PEX14 at the protein level would further our understanding of its functions. Moreover, FMU-PEX14 also could be used in Western blot analysis, and the cross-reaction of this MAb with OmicsLink™ ORF Cell Free Expression library was tested. Approximately 13,000 human ORF clones were transferred into several pIVEX-based expression vectors in the library. All of the specially constructed ORF expression clones were tested for expression synthesis by either SDS-PAGE or Western blot with an anti-tag antibody. No cross-reaction of FMU-PEX14 with other proteins was found (data not shown).
In conclusion, we generated a novel MAb specific for PEX14 in immunocytochemistry and immunohistochemistry and localized PEX14 in human normal and malignant tissues. Further studies should be done to demonstrate the different expression profiles of human PEX14 at the protein level in human normal and malignant tissues.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (no. 30901358; no. 81072116).
Author Disclosure Statement
The authors have no financial interests to disclose.