
Abstract
Prions, or infectious proteins, cause a class of uniformly fatal neurodegenerative diseases. Prions are composed solely of an aberrantly folded isoform (PrPSc) of a normal cellular protein (PrPC). Shared sequence identity of PrPSc with PrPC has limited the detection sensitivity of immunochemical assays, as antibodies specific for the disease-causing PrPSc isoform have not been developed. Here we report the generation of three new monoclonal antibodies (MAbs) to PrP, which were isolated following immunization of Prnp0/0 Balb/cJ mice with highly purified PrPSc isolated from brain lipid rafts. Epitope mapping using synthetic PrP peptides revealed that the three MAbs bind different epitopes of PrP. The DRM1-31 MAb has a conformational epitope at the proposed binding site for the putative prion conversion co-factor “protein X.” The DRM1-60 MAb binds a single linear epitope localized to the β2–α2 loop region of PrP, whereas DRM2-118 binds an epitope that includes sequences within the octarepeat region and near the site of N-terminal truncation of PrPSc by proteinase K. Our novel anti-PrP MAbs with defined PrP epitopes may be useful in deciphering the conformational conversion of PrPC into PrPSc.
Introduction
Unlike PrPC, much of the PrPSc molecule is resistant to limited proteolysis with proteinase K (PK). Thus, in the presence of PK, the protease-sensitive PrPC is hydrolyzed whereas resistant PrPSc is N-terminally truncated and a readily detectable C-terminal fragment (PrP 27–30) remains.(10) Current immunoassays for PrPSc exploit this difference, and detection of PrP 27–30 is used to corroborate disease diagnosis. Antibodies used for these tests bind common epitopes present in both PrPC and PrPSc as no conformation-specific antibody has been generated that can directly distinguish PrPSc from PrPC.(11) Although antibodies that immunoprecipitate PrPSc have been reported,(12,13) none appear to possess the selectivity necessary for development into a reliable immunoassay. Recent studies have described infectious PrP that is not PK resistant, further demonstrating the need for a PrPSc-specific antibody.(14)
Efforts to generate PrPSc-selective antibodies have been disappointing; this lack of success is likely a result of the physiochemical properties shared between PrPC and PrPSc that render the two isoforms indistinguishable immunologically. As expected, wild-type (wt) animals are tolerant to the normal protein PrPC.(15,16) This tolerance probably accounts for the lack of an immune response to PrPSc during prion infection. The generation of the Prnp0/0 mice overcame the barrier associated with self-recognition of PrPSc as these mice elicit a robust immunoresponse when challenged with either PrPC or PrPSc.(17,18) However, the immune system may produce antibodies that recognize epitopes shared by both PrPC and PrPSc. Likewise, immunization with synthetic PrP peptides or recombinant PrPC produced antibodies that bind both PrPC and denatured PrPSc.(17,19,20) Therefore, antibodies that can distinguish PrPSc may recognize rare epitopes that are only occasionally presented to the immune system. In such a case, exploitation of hybridoma technology to dissect the immunological response will be critical for isolating rare monoclonal antibodies (MAbs) with conformational epitopes.
In this report, we describe a series of new anti-PrP MAbs isolated from Prnp0/0 Balb/cJ mice(18) immunized with a highly purified PrPSc immunogen derived from prion-infected hamster brain. Furthermore, we describe a direct-binding ELISA that uses prion-enriched detergent-resistant membranes (DRMs) from normal and Sc237-infected hamster brain as part of an overall hybridoma screening strategy to favor identification of anti-PrP MAbs. We report here the characterization of these new anti-PrP MAbs and identify novel binding epitopes, which have been implicated in the conversion of PrPC into PrPSc.
Materials and Methods
Reagents
All reagents were of the highest grades commercially available. Antibodies were diluted in 10 mM Tris-buffered saline with 1% Tween-20 (TBST) containing 1% IgG-free BSA (Jackson Immunochemicals, West Grove, PA) or TBST containing 3–10% nonfat dry milk (NFDM) powder. Secondary antibodies include: goat-anti-mouse-HRP and goat-anti-rabbit-HRP (diluted 1:10,000; Pierce, Rockford, IL) or goat-anti-mouse-HRP (diluted 1:5000; Sigma, St. Louis, MO). Recombinant Syrian hamster (recSHa) PrP(90–231), recombinant mouse (recMo) PrP(89–230), recombinant ovine (recOv) PrP(23–231), and recombinant human (recHu) PrP(90–231) were generated at UCSF as previously described.(20–22) A series of peptides was synthesized, each of 12 amino acids in length with a 7-residue overlap spanning bovine PrP(25–241) (Table 1). Three lysine residues were added to the N-terminus of each peptide to improve solubility; a single biotin molecule was also added to the N-terminal lysine residue. The C-termini were amidated (Sigma Genysis, Woodlands, TX). Peptides were dissolved in 10% acetonitrile and diluted to 1 mM in 10% ethyl alcohol.
Three lysine residues were added to the N-terminus of each peptide, with the N-terminal lysine labeled with biotin, and the C-terminus was amidated.
Animals
Animals were housed as previously described(18) in compliance with the USDA Western Regional Research Center's Institutional Animal Care and Use Committee approved protocols. Hamster-passaged Sc237 scrapie prions (PrPSc) were propagated in female Syrian golden hamsters,(18,23) and experimental procedures using infectious prions were conducted in BSL2 laboratories. Generation and immunization of Prnp0/0Balb/cJ mice were previously described.(18)
Immunogen
PrPSc was first isolated in DRM fractions from Sc237-infected Syrian hamster brain homogenate and processed to obtain a purified PrP 27–30 immunogen as previously described.(18) Briefly, hamster brains were homogenized (10% w/v) on ice in 25 mM MES (pH 6.5) with 150 mM NaCl, 1% Triton X-100, 60 mM n-octyl-glucoside, 10 mM PMSF, and protease inhibitors (Complete mini; Roche, Nutley, NJ). Brain homogenates were fractionated by ultra-centrifugation using a discontinuous sucrose gradient and the DRM fraction digested with 150 μg/mL of PK for 1 h at 60°C. Protease digestion was then inhibited by 10 mM PMSF. PrPSc was concentrated by phosphotungstic acid (PTA) precipitation, and the PK-resistant PrPSc further separated from the PK and proteolytic products by size exclusion chromatography on a Sephadex G100 column. PrPSc was recovered in the void volume, concentrated by a second PTA precipitation, and resuspended in buffer. This material (referred to as the immunogen) was previously shown to be infectious and to migrate as a single Coomassie staining protein band with a molecular mass consistent with the PK-resistant PrPSc core (PrP 27–30).
Immunization
Prnp0/0 Balb/cJ mice (25 day old) were immunized with two bilateral intraperitoneal (i.p.) inoculations of 100 μL of purified hamster PrPSc immunogen (11.6 μg, measured by BCA, Pierce) in adjuvant (Sigma Adjuvant System; Sigma) separated by 10 days.(18) A third immunization boost was performed 10 days after the second injection without adjuvant and 3 days prior to cell fusion.
Antibody generation
Cell fusions were performed as previously described.(24) Ten to 14 days following cell fusion, media from culture wells containing hybridoma cells were screened using a direct-binding ELISA for antibody binding to prion-enriched DRM preparations obtained from diseased hamster brain homogenates (PrPSc-DRM).(18) Since the majority of hybridoma cells were not expected to produce antibodies, the averaged ELISA activity on a plate was taken to represent background. Cells from positive wells, those with a response at least five times the background, were transferred to 24-well culture plates, allowed to grow for at least 24 h, and the media analyzed by ELISA for antibody binding to recSHaPrP(90–231) and to DRM fractions obtained from normal (PrPC-DRM) and diseased (PrPSc-DRM) brain treated with and without PK. Finally, hybridoma cells from ELISA-positive wells were cloned at least twice by limiting dilution and media evaluated by Western blot analysis. Cloned hybridoma cells were used to generate ascites in Balb/cJ mice (Covance Immunology Services, Denver, PA) and MAbs purified by affinity chromatography on protein-G Sepharose (Pierce).
ELISA
Antigens were diluted in 0.1 M sodium bicarbonate buffer (pH 9.4) and 100 μL absorbed to 96-well Maxisorb plates (Nunc, Rochester, NY) overnight at 4°C. Plates were washed in TBST, blocked in 10% NFDM for 1 h, incubated for 1 h with primary antibody, washed, incubated for 1 h with HRP-conjugated secondary antibody, washed, resolved by chemiluminescence (Supersignal; Pierce) detection using a Victor 2 plate reader (PerkinElmer, Waltham, MA), and results expressed as relative light units (RLU). Isotype analysis was determined by ELISA using isotype-specific antibodies (Southern Biotechnology, Binghamton, AL).
Antibody sequencing
The mRNA coding for the PrP MAbs were extracted and purified from hybridoma cells, converted to cDNA, and the heavy (Hc) and light (Lc) chains of each antibody amplified using polymerase chain reaction (PCR).(25) Circularized vector pCR2.1 (Invitrogen, Carlsbad, CA) was digested with EcoRV and treated with calf intestinal phosphatase (New England BioLabs, Bethesda, MD). PCR products were gel-purified and treated with polynucleotide kinase (New England BioLabs) ligated into vector pCR2.1 and transformed into TOP10 cells (Invitrogen) according to manufacturer's instruction. Colonies were picked and grown aerobically in 5 mL of Luria-Bertani (LB) medium at 37°C supplemented with 100 μg/mL ampicillin.(26) Plasmids were purified using the QuickClean 5M range of kits (GenScript Corp, Piscataway, NJ). Automated DNA sequencing was performed using the Big Dye Terminator Version 3.1 and XTerminator reagents, and a 3730 DNA Analyzer (Applied Biosystems, Foster City, CA). Primers M13F (GTAAAACGACGGCCAG) and M13R (CAGGAAACAGCTATGAC) were used for sequencing the DNA insert in plasmid pCR2.1.(27)
All Lc clones were amplified with the same primers, 3′ Kc and 5′ Mk, and the Hc primers are as follows: DRM1-15, 5′ MH2 and 3′ IgG2b; DRM1-26, 5′ MH2 and 3′ IgG2b; DRM1-31, 5′ MH2 and 3′ IgG2b; DRM1-60, 5′ MH2 and 3′ IgG2b; and DRM2-118, 5′ MH2 and 3′ IgG1 as described.(25)
Western blot analysis
Western immunoblotting was performed as described.(23) Briefly, samples (brain homogenates and DRM preparations) were separated by electrophoresis through 4–12% Bis-Tris gels with MES running buffer (Novex, Invitrogen). Proteins were transferred to nitrocellulose (Bio-Rad, Hercules, CA), washed in TBST, blocked with 10% NFDM, probed with antibodies, protein bands resolved by ECL (Supersignal, Pierce), and imaged on a Fluorochem HD documentation system (Alpha Innotech, San Leandro, CA).
Epitope determination
A series of 12-residue peptides spanning bovine PrP(25–241) (NCBI accession # CAA39368), which corresponds to hamster sequences 23–231 (NCBI accession # P04273) (see Table 1), was synthesized and used in ELISA, as previously described,(20) to localize antibody binding to specific epitopes. Each peptide contained 12 amino acids of the PrP sequence with a 7-residue overlap. Three lysine residues were added to the N-terminus of each peptide to improve solubility, the N-terminal lysine residue was biotinylated, and the C-termini were amidated. Two peptides in this series corresponding to Bo145MSRPLIHFGSDY156 and Bo200VTTTTKGENFTE211 failed in the synthesis and were not available.
Peptides were dissolved in 10% acetonitrile and diluted to 1 mM in 10% ethyl alcohol. On the day of the experiment, streptavidin-coated microtiter wells (Pierce) were blocked for 60 min at room temperature (RT) by adding 300 μL/well of 5% NFDM-TBST. Peptides were diluted in TBST (pH 7.4) to a final concentration of 5 μM, then added (50 μL) to the plates and incubated for 1 h at RT. Anti-PrP antibodies were then added (50 μL) and the plate incubated for 1 h at 37°C. Following three washes with 200 μL/well of TBST, 100 μL/well of HRP-conjugated, goat anti-mouse IgG (Pierce), diluted 1:10,000 in 1% BSA-TBST, were added and the plates incubated for 1 h at RT. Finally, the plates were washed three times with TBST; then 100 μL/well of substrate (Supersignal, Pierce) was added and luminescence measured with a Victor 3 plate reader (PerkinElmer). Peptide sequences were aligned for maximum sequence conservation.
Results
Antibody production
Recent studies demonstrated production of high-titer anti-PrP sera following immunization with highly purified PK-resistant hamster PrPSc in recently bred Prnp0/0 on a Balb/cJ background but not wt mice.(18) We used splenocytes from Prnp0/0 Balb/cJ mice immunized with a purified PrPSc immunogen derived from brain DRMs to generate hybridomas. The hybridomas were subsequently screened for production of anti-PrP antibodies using a direct-binding ELISA on microtiter plates coated with PrPSc-DRM preparations purified from brain homogenates of prion-infected Syrian hamsters. Hybridoma cells from wells with chemiluminescence signals greater than five times the plate background (Fig. 1; dotted line) were selected, and antibody binding further characterized using a panel of antigens consisting of recSHaPrP(90–231), PrPC-DRMs, and PrPSc-DRMs (before and after PK treatment). Those cells producing antibodies suggestive of binding PrPSc (i.e., minimal binding on recSHaPrP(90–231) and PrPC-DRMs, but greater than five times binding on PrPSc-DRMs) were cloned by limiting dilution. Subsequent ELISA tests suggested that these MAbs bound both PrPC and PrPSc. Monoclonal antibodies were then produced as ascites and purified by protein-G affinity chromatography. Five MAbs were identified and designated DRM1-15, DRM1-26, DRM1-31, DRM1-60, and DRM2-118. Isotype analyses revealed that DRM1-15, 26, 31, and 60 have IgG2b heavy chains while DRM2-118 is an IgG1 antibody, all with kappa light chains.
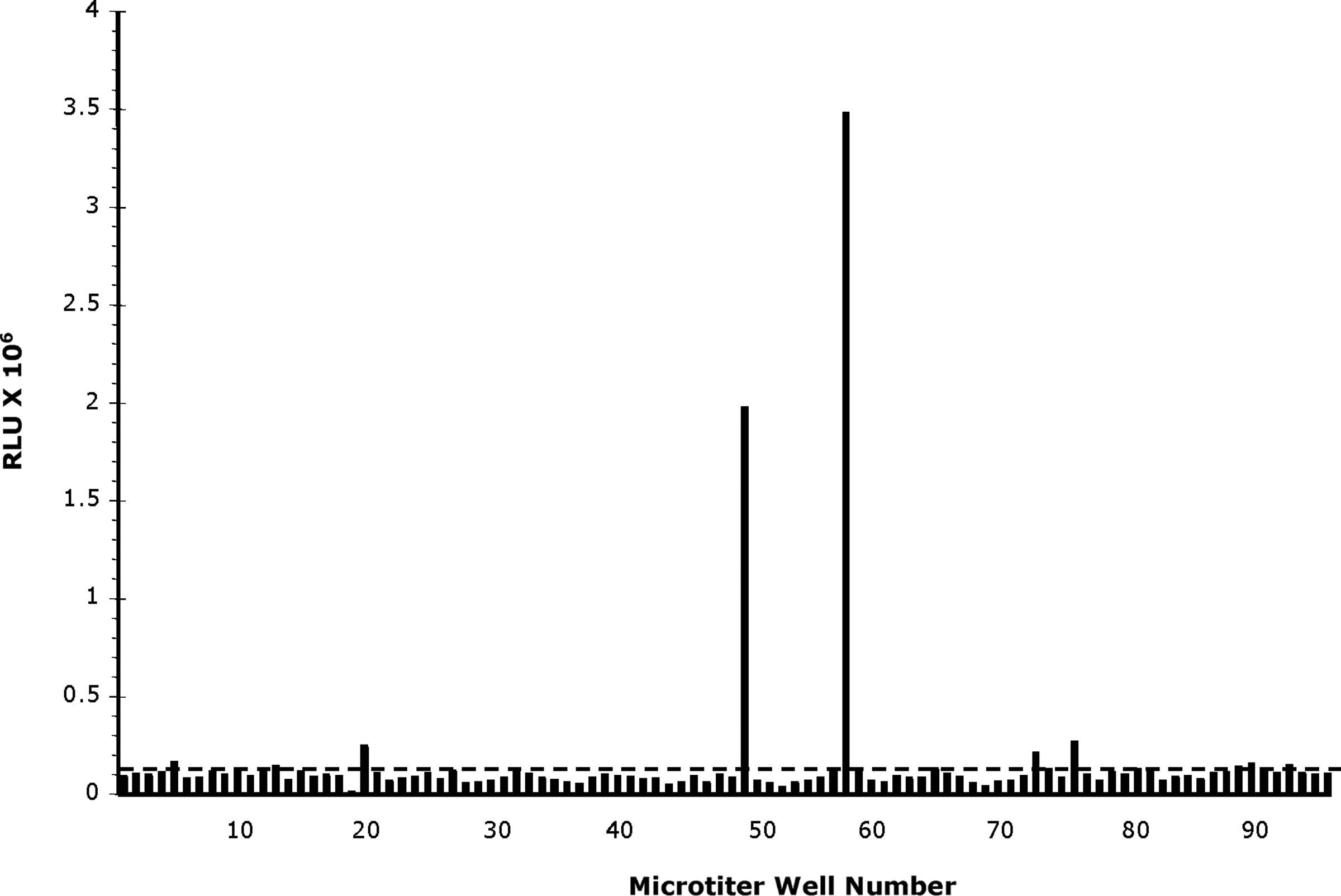
Direct-binding ELISA results from the initial screening of a cell-fusion experiment using splenocytes of PrPSc-immunized mice. A microtiter plate was coated with purified PrPSc-DRM following PK digestion. Since the majority of wells of a screening plate are not expected to contain hybridomas secreting specific antibody, the plate background was calculated as the average (dashed line) relative luminescence units (RLU), 0.15×106 RLU for the plate shown.
Antibody specificity
The binding specificity of the five MAbs to recombinant PrP was evaluated in ELISA titration experiments; we used recPrP harboring the sequences from SHa (90–231), mouse (Mo; 89–230), sheep (Ov; 23–231), and human (Hu; 90–231)
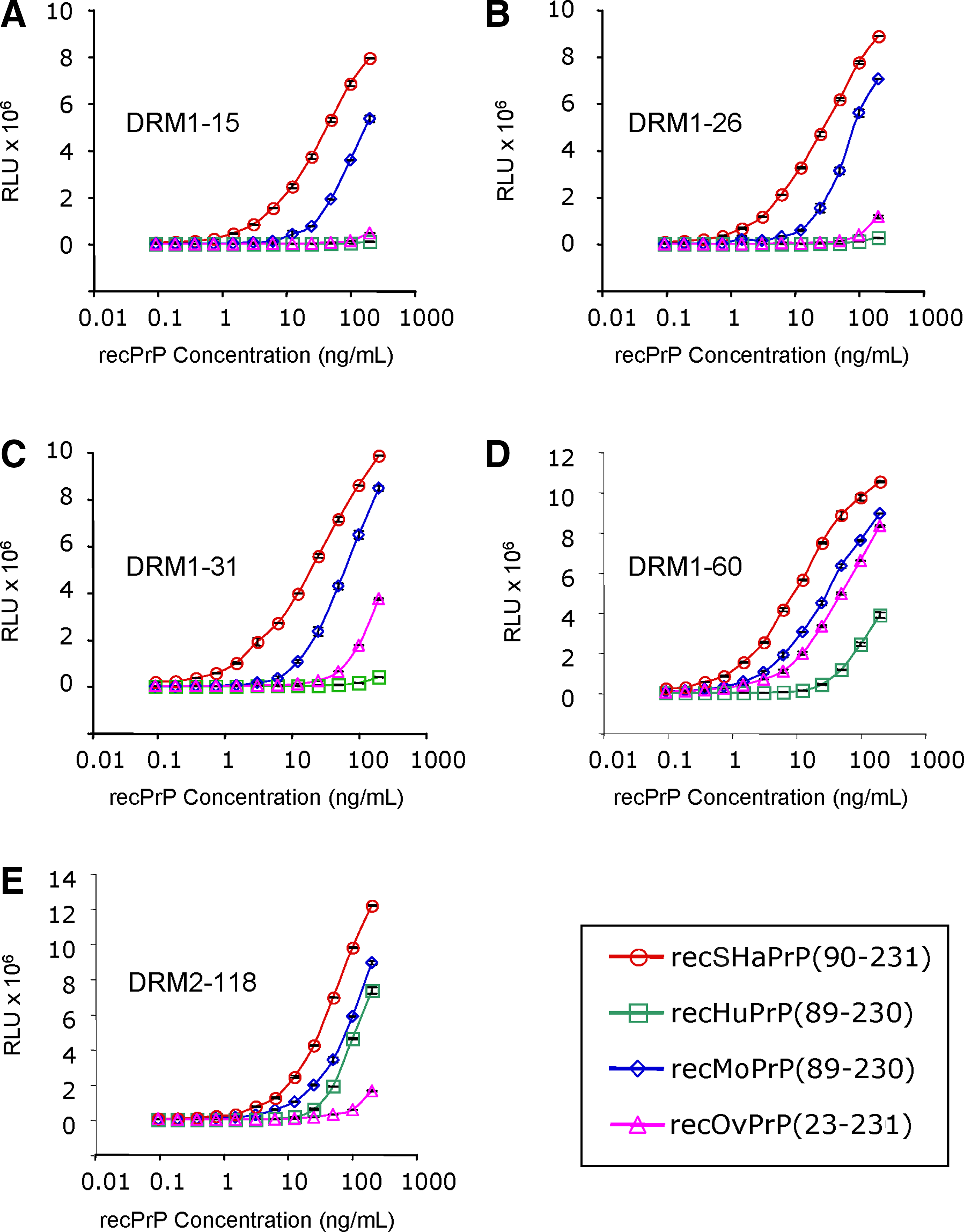
ELISA binding of five MAbs—(
Antibody binding to full-length PrPC from brain homogenates derived from eight different species was compared by Western blot analysis (Fig. 3). As expected, all five MAbs recognized denatured, full-length, glycosylated SHaPrPC containing the GPI anchor (Fig. 3). MAb DRM2-118 recognized PrPC from all the species tested except for cervid (Fig. 3; top panel). Antibody DRM1-60 detected PrPC from hamster, mouse, sheep, cow, pig, and white-tailed deer, but not horse or human PrPC (Fig. 3; second panel). DRM1-31, DRM1-26, and DRM1-15 showed strong binding to SHaPrPC and weak but detectable binding to MoPrPC and CerPrPC (Fig. 3, bottom three panels).
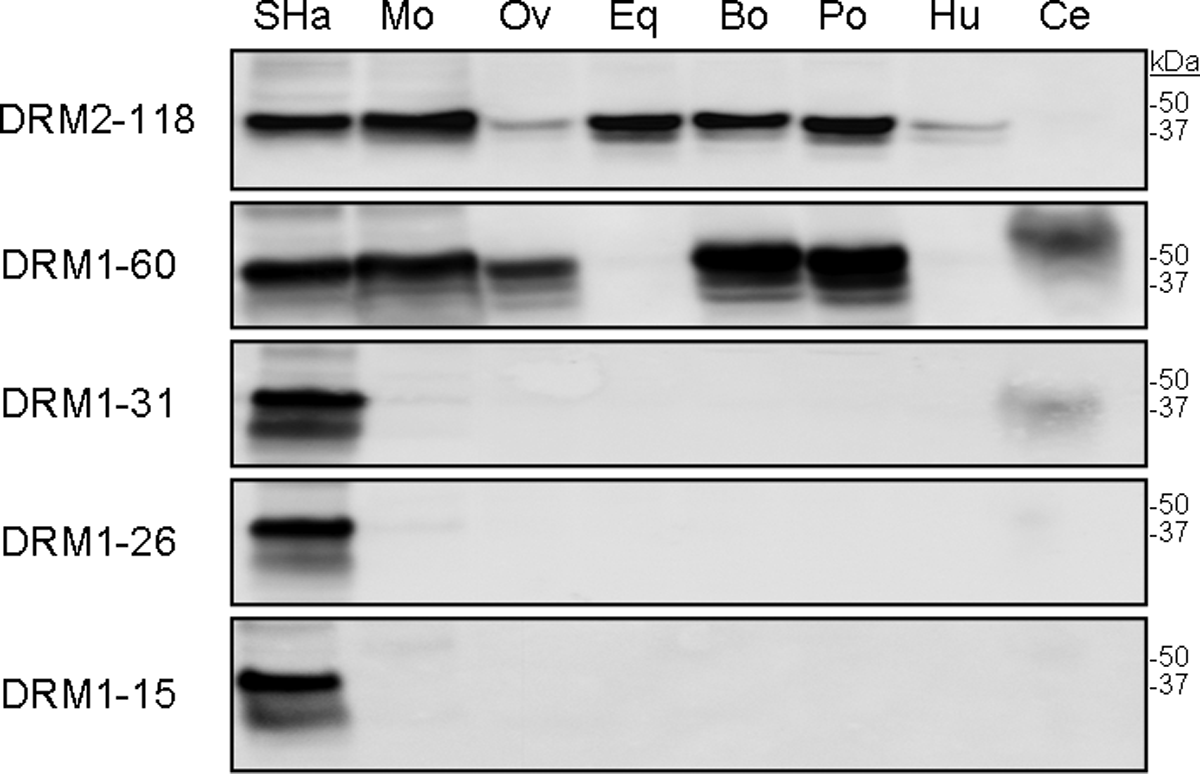
Western blot detection of PrPC in 40 μg of normal brain homogenates with the five MAbs. Homogenates were prepared from uninfected whole Syrian hamster brain (SHa); whole mouse brain (Mo); sheep cortex (Ov); horse cortex (Eq); cow cortex (Bo); pig cortex (Po); whole human brain (Hu); and white-tailed deer obex (Ce). For each MAb, 2 μg/mL was used. Molecular masses of protein standards are shown in kilodaltons (kDa).
Antibody variable region sequences
The binding patterns observed with MAbs DRM1-15, DRM1-26, and DRM1-31, to both recombinant PrP and native PrPC derived from normal brain homogenates of different species, suggest that they bind the same or a closely related epitope. To clarify their relationship to each other, the nucleic acid sequence and derived amino acid sequences of the variable region for both the Lc and Hc of each antibody were determined (sequences available from GenBank: accession # DRM1-15_Lc HQ218926; DRM1-26_Lc HQ218927; DRM1-31_Lc HQ218928; DRM1-60_Lc HQ218929; DRM2-118_Lc HQ218930; DRM1-15_Hc HQ218931; DRM1-26_Hc HQ218932; DRM1-31_Hc HQ218933; DRM1-60_Hc HQ218934; DRM2-118_Hc HQ218935). Alignment of the amino acid sequences is shown for the entire cloned region from the beginning of framework region one (FRW-1) to the end of the fourth framework region (FRW-4). The J-region is shown in bold, the amino acids corresponding to the PCR primers are underlined, and the complementarity determining regions (CDRs) are shaded (Fig. 4). The leader sequences, framework regions, CDRs, and J-regions were assigned by comparison of the Hc and Lc of these five MAbs to other antibody sequences.(28–32) Antibodies DRM1-15, DRM1-26, and DRM1-31 have identical variable Hc and Lc amino acid sequences (Fig. 4), and most likely represent independent cell fusions of a clonally expanded lymphoid population. Therefore, for these three MAbs, only DRM1-31 was further characterized. Both MAbs DRM1-60 and DRM2-118 have variable region amino acid sequences distinct from each other and from the DRM1-15, -26, and -31 group (Fig. 4).

Amino acid sequences for the variable regions of DRM1–15, DRM1-26, DRM1-31, DRM1-60, and DRM2-118 kappa light and heavy chains. The deduced amino acids, from the first (FWR-1) to the end of the fourth framework region (FWR-4) are shown. The three complementarity determining regions (CDRs) are shaded, J-region is shown in bold, amino acids coded for by the primers are underlined, and framework regions (FWR-1, FWR-2, FWR-3, and FWR-4) are indicated.
Peptide mapping
In an effort to further define the epitopes, antibody binding to a library of synthetic peptides spanning the entire PrP sequence was evaluated. The library, used in a previous study,(19) consisted of 12-residue peptides spanning the bovine PrP sequence (NCBI accession # CAA39368), each overlapping by seven amino acids (Table 1). Because 11 of the 16 differences between SHa and Bo PrP sequences are strongly conserved amino acids (two represent deletions), the Bo peptide library was used as a tool to evaluate antibody binding. To improve peptide solubility, three lysine residues were added to the N-terminus of each peptide and the terminal lysine was biotinylated. The peptides were oriented and immobilized using streptavidin-coated, 96-well microtiter plates. Results of DRM antibody binding to this library are summarized in Figure 5. The simplest binding pattern was observed with MAb DRM1-60; only peptide #29 was recognized (Fig. 5B), suggesting a linear binding epitope within the sequence 170NQVYYRPVDQYS181 corresponding to SHaPrP(159–170) (NCBI accession # P04273). The carboxy-terminal Ser (S) at bovine position 181 is not critical for binding since the hamster sequence has an Asn (N) in this position. Antibody DRM1-31 displayed a more complex peptide-binding pattern (Fig. 5A), binding to spatially separated peptide #29, and peptides #37–39. These data suggest a discontinuous or conformational epitope, incorporating the three segments: 170NQVYYRP176 (the β-2 sheet region); the amino end of the second α-helical region, 185NFVHDCV191; and a segment near the carboxyl end of the third α-helical region, 222EQMCITQY229. Antibody DRM2-118 bound a number of peptides located in the N-terminal non-structured region of PrP (Fig. 5C). Specifically, strong binding was observed to peptides #9, #14–#17, as well as to peptide #44, which is a modification of peptide #16, in which T106 and G108 were both substituted with Ser (Table 2). Peptide alignment that results in the largest invariant sequences suggests an epitope motif containing the sequence GWGQ (Table 2). This sequence contains elements of the octarepeat region. The motif is broadly represented in PrP sequences of different species as part of the octarepeat, which is consistent with the broad species reactivity observed (Fig. 3) and suggests the possibility of multiple binding sites for DRM2-118.
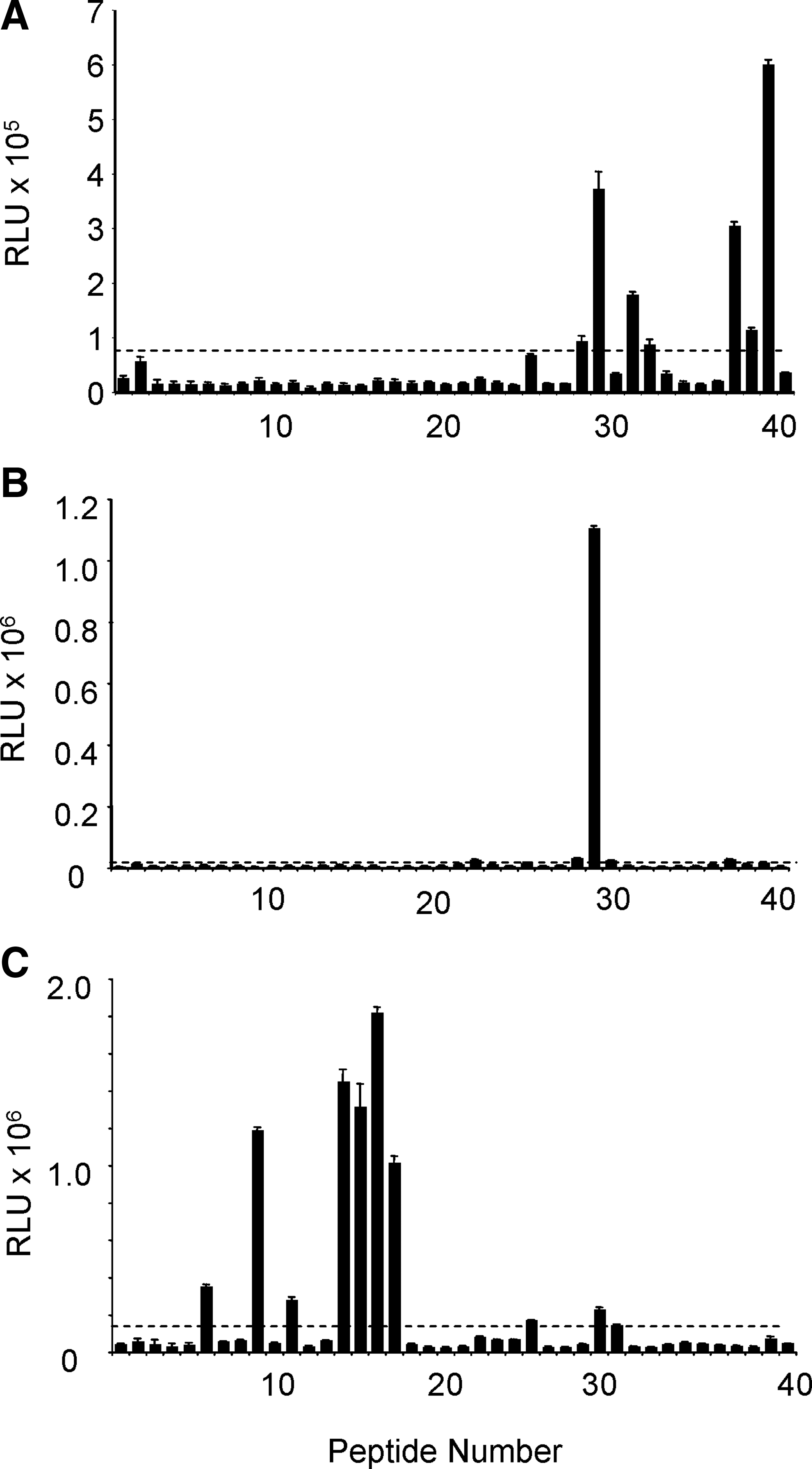
Epitope mapping of MAbs DRM1-31 (
Peptide #44 corresponds to OvPrP(92-103) with two differences from peptide #16 indicated in bold. ELISA binding from Figure 5C is summarized here for convenience. Potential epitope motif is GWGQ.
Bo, bovine (NCBI accession # CAA39368); Ov, ovine (NCBI accession # CAA04236); ++, moderate binding; +++, strong binding.
Detection of PrP Sc
To determine whether the three MAbs recognize PrPSc, we evaluated their binding to purified PrPSc-DRM by Western immunoblotting (Fig. 6). As a control, binding to PrPC-DRM was also tested. All three MAbs DRM1-31, DRM1-60, and DRM2-118 (1 μg/mL) were able to detect PrPSc by Western blot (8 μg/lane; Fig. 6). Following PK treatment, the antibodies effectively detected PK-resistant PrP 27-30, as defined by the molecular weight shift on Western blots [compare undigested samples (-) to digested (+) samples]. In contrast, PK digestion effectively abolished detection in the DRM-PrPC samples by Western blot. In these experiments, detection of PrP 27–30 and total PrP (undigested PrPC+PrPSc) in PrPSc-DRM samples was greater compared to PrPC in DRMs, which likely reflects the greater amount of accumulated PrPSc in DRM preparations compared to PrPC in DRM samples, as observed previously.(18)
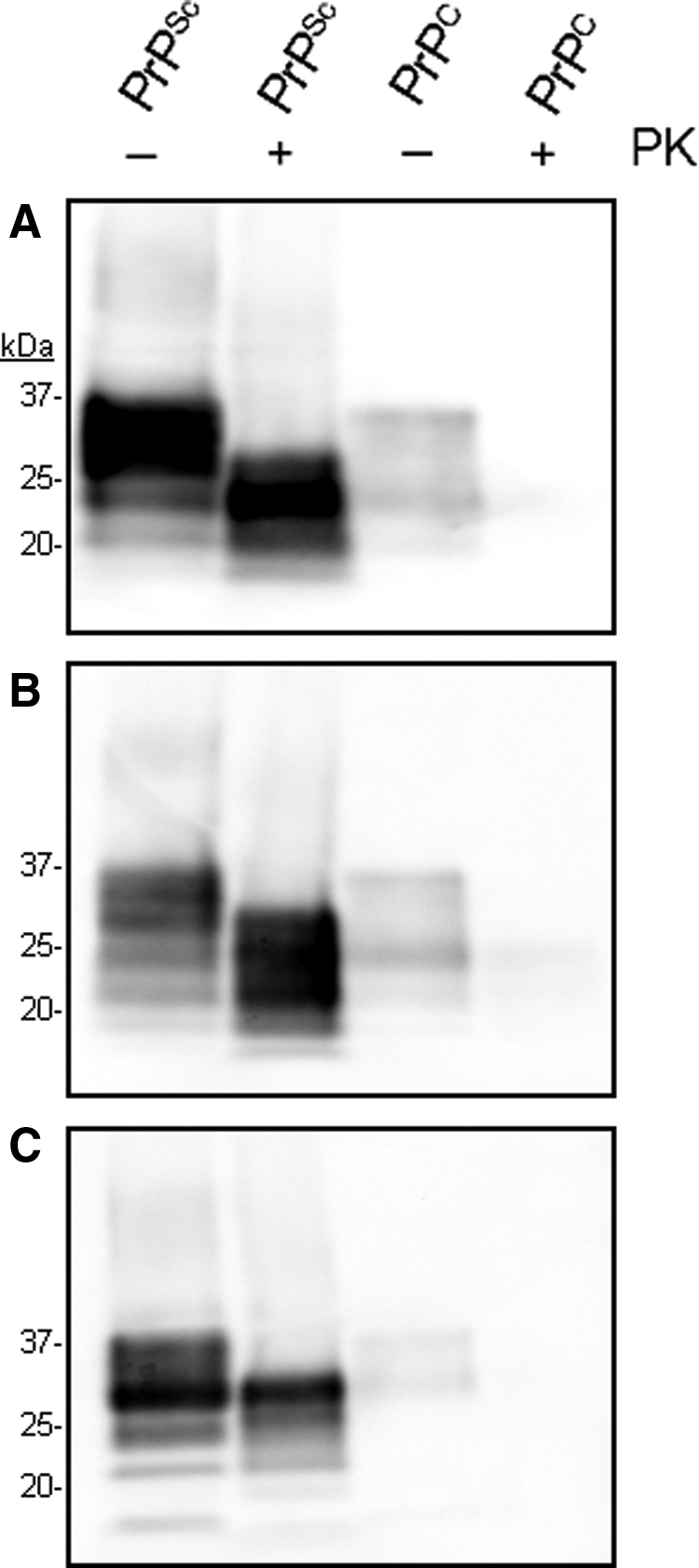
Detection of PrPC and PrPSc in DRMs by MAb DRM1-31 (
Differential antibody binding
Next, we determined whether removal of the disulfide bond in PrP affected binding of the three MAbs. Brain homogenates from prion-infected Syrian hamsters were prepared and boiled for 1 min in electrophoresis sample buffer containing 10% β-mercaptoethanol, then 10 μg was loaded per lane, subjected to SDS-PAGE and Western immunoblotting (Fig. 7). Removal of the disulfide bond in PrP reduced the binding of DRM1-31 and DRM1-60, but had little effect on the binding of DRM2-118.

Effect of disulfide bond disruption in PrP on MAb binding. Scrapie-infected hamster brain homogenates were treated with (+) and without PK (-) in the presence (+) or absence (-) of 10% 2-mercaptoethanol (2-ME) to reduce the single disulfide bond present in PrP. Brain homogenates were loaded at 10 μg/lane and DRM MAb used at 1 μg/mL. Molecular masses of protein standards are shown in kilodaltons (kDa).
Discussion
We have previously demonstrated a novel method to purify and concentrate PrPSc from lipid rafts (DRM), and that immunization of a newly derived Prnp0/0 Balb/cJ mouse line with PrPSc-DRM generated antisera reactive on Western blots to PrP, but not to other proteins present in brain homogenate.(18) The objective of this study was to use hybridoma technology to dissect the immune response in order to isolate novel antibodies that might be employed in novel diagnostics. The most useful antibodies for diagnostics would be those capable of directly distinguishing PrPSc from PrPC. Thus, we employed a differential screening strategy early in the selection to identify antibodies that showed strong binding by ELISA to PK-digested PrPSc-DRM preparations derived from scrapie-infected hamster brain, and little or no binding to PrPC-DRM preparations derived from non-diseased hamster brain homogenate. Of the nearly 100 positive wells identified in our initial ELISA screen (Fig. 1), only five were selected for cloning and further characterization following the results obtained from a secondary screening for activity toward PrPC, both undigested and PK-digested PrPSc as well as recSHaPrP(90–231) (Fig. 2).
While the secondary screening suggested that MAbs DRM1-15, DRM1-26, DRM1-31, DRM1-60, and DRM2-118 preferentially bound PrPSc, subsequent studies demonstrated that all five MAbs also bound to PrPC (Fig. 3). Western blots of brain homogenates generated from healthy hamster, mouse, sheep, horse, cow, pig, human, and white-tailed deer demonstrated that all five MAbs bound hamster PrPC, but differed in their binding to PrPC of other species. Furthermore, the binding patterns observed by our biochemical analysis indicated that the five antibodies represent three epitope groups. Sequencing of the variable regions of the Hc and Lc supported assignment into three epitope groups and further suggested that the antibodies DRM1-15, DRM1-26, and DRM1–31 most likely represent hybridomas derived from independent cell fusions of a clonally expanded lymphocyte population, whereas DRM1-60 and DRM2-118 represent distinct hybridomas (Fig. 4).
Results from epitope mapping (Fig. 5) and disulfide bond reduction (Fig. 7) experiments indicate that MAb DRM1-31 has a conformational epitope, incorporating amino acids from three separated regions: SHaPrP(159–170) (NQVYYRPVDQYS); SHaPrP(174–179) (NFVHDC); and SHaPrP(225–236) (EQMCITQY). These regions correspond to the β2 loop, the N-terminal residues of α-helix 2, and the C-terminal region of α-helix 3, respectively.(7,33,34) Interestingly, the β2–α2 loop and the C-terminal end of the α3 helix form a continuous surface postulated to be the binding site for the hypothetical “protein X” thought to be important in the conformational conversion of PrPC to PrPSc.(35) The epitope motifs from each of these regions were mapped onto the 3-dimensional NMR structure of BoPrP and HuPrP (Fig. 8), demonstrating a juxtaposition of these regions.

Location of DRM MAb binding mapped onto space-filled models of PrPC. The amino acids that contribute to the epitope of DRM1-31 from each of the three peptides clusters (Bo170NQVYYRPVDQYS, red; Bo180NFVHDC, green; and Bo225EQMCITQY, blue) are shown in the left panel. Amino acids Bo170NQVYYRPVDQYS (red) that contribute to DRM1-60 MAb binding are depicted in the center panel. Amino acids GWGQ (purple) that contribute to DRM2-118 binding are shown in the right panel. Space-filled models used for DRM1-31 and DRM1-60 from Protein Database (PDB) file 1DX0 and for DRM2-118 from PDB file 2KUN.
Characterization of MAbs DRM1-60 suggest a linear epitope (Figs. 5 and 7; Table 2). DRM1-60 showed strong binding to only one peptide (#29: NQVYYRPVDQYS) suggesting a linear epitope corresponding to SHaPrP(159–170), located in β2-α2 loop. This epitope assignment is consistent with the binding of DRM1-60 to PrPC to all species tested, except human and horse, which both harbor substitutions in this region. V166 and Q168 in SHaPrPC are Met and Gha in HuPrPC, respectively. For the horse sequence, Ser is expressed at codon 170 instead of Asp (Table 3). Disruption of the disulfide bond by 2-mercaptoethanol reduced DRM 1–60 did not affect binding to PrP, an observation consistent with a linear epitope (Fig. 7).
SHaPrP, Mesocricetus auratus, NCBI accession #P04273; MoPrP, Mus musculus, AAA39996; BoPrP, Bos taurus, CAA39368; HuPrP, Homo sapiens, AAH22532; EqPrP, Equine caballus, ACG59276; CePrP, Odocoileus virginianus, AAQ23191; and OvPrP, Ovis aries, CAA04236. Only the amino acids that differ from SHaPrP are indicated; dots denote identical amino acids.
By using a highly purified preparation of PrPSc obtained from lipid rafts and novel Prnp0/0 Balb/cJ mice, we identified three new anti-PrP monoclonal antibodies, DRM1-31, DRM1-60, and DRM2-118. While antibodies with epitopes similar to DRM2-118 have been described,(36) DRM1-60 and DRM1-31 have unique epitopes not previously reported. DRM1-60 appears to bind a linear epitope localized to the β2-α2 loop, while DRM1-31 has a conformational epitope located on the exposed surface formed by the close proximity of the β2-α2-helix loop, the N-terminus of the α2 helix, and the C-terminus of the α3 helix.
The antibodies described here, while clearly binding recombinant PrP in ELISA and PrPC by Western blot, appear to have a preference for PrPSc that is increased 3-fold following limited PK digestion (PrP 27–30). PK digestion of samples may remove unstructured portions of prion aggregates, exposing more antibody-binding epitopes that results in an increase in total signal detected by immunoassay.
Conclusion
We report here the generation and characterization of three new anti-PrP MAbs obtained following immunization of Prnp0/0 Balb/cJ mice using purified PrPSc-DRM. Characterization of these antibodies revealed binding to PrP with distinct species specificity, a strong affinity for PrPSc, and binding to novel epitopes in the PrP structure not previously reported. Some of these antibodies may prove useful in identifying putative co-factors or binding partners involved in PrPC conversion to PrPSc.
Footnotes
Acknowledgments
Support for these studies was provided by USDA CRIS Project 5325-32000-008-00D, by a USDA Cooperative Agreement (58-5325-3-246), by grants from the National Institutes of Health (AG02132, AG10770, AG-031220, and AG021601), and by a gift from the Sherman Fairchild Foundation.
Author Disclosure Statement
The authors have no financial interests to disclose.