
Abstract
As a transmembrane enzyme, ATP synthase plays an important role in energy metabolism of organ tissues, as well as in tumors. In this study we generated a monoclonal antibody, 6G11, to the catalytic subunit of F1-F0 ATP synthase (ATP5B). The SDS-PAGE result demonstrated that the hybridoma clone had a molecular weight of 50 and 27 kDa components that could be the heavy and light chains of the monoclonal antibody, respectively. Chromosome analysis of the hybridoma clone proved that they had 98 to 102 chromosomal numbers that were the sum of the SP2/0 and spleen cells. Western blot assay revealed that the hybridoma clone reacted specifically with the ATP synthase beta subunit, but not with other proteins. In addition, the subclass of the hybridoma clone was identified as IgG1 by capture ELISA. Furthermore, it demonstrated that the antibody retained stability after half a year. These results indicated that the hybridoma clone 6G11 was a monoclonal antibody with significant stability and special reactivity to ATP5B antigen.
Introduction
How does this system work? Many scientists have constructed models to explain the mechanisms.(10,11) The transmembrane enzyme consists of two dominant domains, including the proton-transporting membrane F0 domain and the catalytic peripheral F1 domain.(12–15) ATP synthase uses physical rotation of its own subunits as the catalysis step, and its work depends on the systemization of F1F0-ATPase.(16) When the energy levels are enough or beyond what is required, the membrane potential decreases and inhibits it, and the F1F0-ATPase starts to work as a proton pump and hydrolysis ATP.(17–20)
Polyclonal antibodies against ATP synthase have been achieved.(21–24) With the development of somatic cell hybridization techniques, large quantities of high titer monoclonal antibodies can be produced and are currently applied to cancer therapeutics. In this report, a murine monoclonal IgG1 immunoglobulin has been developed with specificity for β-subunit of ATP synthase.
Materials and Methods
Expression vectors of pET-28a (+)-atp5b and pET-32a (+)-atp5b were previously constructed by our laboratory. Isopropyl β-D-1-thiogalactopyranoside (IPTG) and β-mercaptoethanol were purchased from Sangon (Shanghai, China). Ni2+-NTA columns were purchased from GE Healthcare Biosciences (Uppsala, Sweden). BCA protein assay kit was purchased from Beyotime (Haimen, Jiangsu, China) and the BALB/c mice were purchased from Wushi Animal Laboratory (Fuzhou, China). The non-secretory cell line SP2/0 was purchased from Shanghai Cellbank (Shanghai, China). Horseradish peroxidase (HRP) labeled anti-mouse IgG, HAT (hypoxanthine, aminopterin, thymidine), HT (hypoxanthine, thymidine), and RPMI 1640 medium were purchased from Sigma-Aldrich (St. Louis, MO). Fetal calf serum was obtained from Hyclone (Logan, UT).
Cell culture
SP2/0 cells were grown in RPMI 1640 medium with 10% fetal calf serum (FCS). Fusion hybridoma clones were grown in RPMI 1640 medium containing HAT or HT during the fusing stage. All cells were grown at 37°C in a humid air incubator with 5% CO2.
Expression and purification of ATP synthase subunit
The recombinant vectors of pET-28a (+)-atp5b and pET-32a(+)-atp5b have been successfully constructed and identified previously and were transferred into Escherichia coli BL21 (DE3) cells to express the fusion protein. The recombinant protein was then induced by IPTG (1 mmol/L) overnight at 25°C, and the expressed protein was purified by affinity chromatography using Ni2+-NTA columns. Finally, the purified protein was analyzed by 12% SDS-PAGE, and its concentration was detected by a BCA protein assay kit.
Immunization and anti-serum titer assays
Six-week-old female BALB/c mice were immunized with a mixture of 100 μg purified recombinant protein and equal volume complete Freund's adjuvant by s.c. injection as the first immunization. After 3 weeks, the mice were immunized in the same manner, but using incomplete Freund's adjuvant. After four immunizations, titer of the anti-serum was determined by indirect non-competitive enzyme-linked immunosorbent assay (ELISA).
Cell fusion and screening of hybridoma clones
Three days after the final boost, the spleen was removed and the dissociated spleen cells were fused with SP2/0 myeloma cells at the rate of 5:1. Polyethylene glycol was used as fusion reagent. After fusion, the cells were cultivated in four 96-well plastic plates using RPMI 1640 medium containing 10% FCS and HAT. Subsequently, HAT medium was replaced by HT. The supernatant of the 96-well microtiter plates that contained hybridoma clones were screened by indirect non-competitive ELISA.
Identification of monoclonal antibody
To identify the monoclonal antibody, SDS-PAGE was adopted to test the supernatant of the hybridoma clone. Chromosome analysis was carried out by treatment with colchicum. Harvested cells were treated with colchicum with the final concentration of 0.1 μg/mL at 37°C, 5% CO2 for 10 h. Then the cells were centrifuged at 1000 r/min for 10 min. KCl (0.075 mol/L, 5 mL) was added to the precipitate and incubated at 37°C , 5% CO2 for 30 min to obtain low osmosis. After centrifugation, the precipitate cells were suspended by HAc-Methanol stationary liquid (1:3) and kept stationary for 30 min at room temperature. Then, the last step was repeated. The glass slide loading stationary liquid above (containing precipitate cells) was stained with 10% Giemsa dye for 10 min. Finally, the chromosome was observed under the microscope.
Cross-reactivity assay
To test the specificity of the anti-ATP5B hybridoma clones, Western blot analysis was performed. Briefly, GST, SIRT5, IDH3α, and ATP5B were dissolved in 2×SDS sample buffer at the rate of 5:1, respectively. After electrophoresis, the gel containing aim bands were transferred onto polyvinylidene difluoride. Subsequently, the membrane was blocked in blocking buffer (5% nonfat milk, Tris buffer) at 37°C for 2 h. After washing three times, the membranes were hybridized with the supernatant of the hybridoma clone at 37°C for 30 min. During this wash step, the second antibody of the goat anti-mouse with AP label was added onto the membrane at 37°C for 2 h. The membrane was washed three times, and then substrate solution was added and incubated at 37°C for 15 min. Finally, 2 mol/L H2SO4 was added to stop the reaction.
Determination of titer and Ig subclass of anti-ATP5B antibody
Determination of antibody titer was carried out by indirect non-competitive ELISA. The primary antibody was the supernatant of the hybridoma clone diluted at the rate of 1:100, 1:200, 1:400, 1:800, 1:1600, 1:3200, 1:6400, and 1:12,800, and the second antibody was the anti-mouse antibody with HRP label. To analyze the Ig subclass, the coating antigens were IgM, IgA, IgG2a, IgG2b, IgG1, and IgG3, respectively.
Determination of relative affinity constant of anti-ATP5B antibody
Measurement of monoclonal antibody affinity constant (Ka) was carried out according to the procedure described by Beatty and colleagues.(25)
Results
Expression and purification of ATP synthase β-subunit
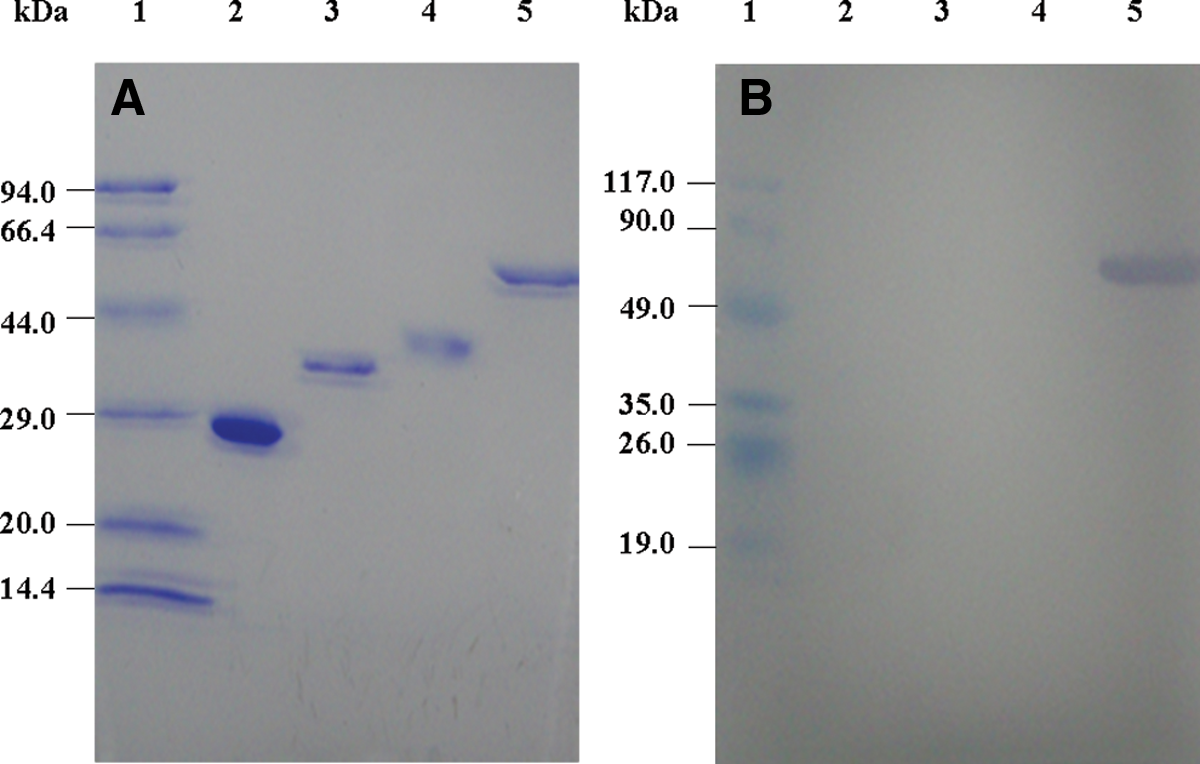
In order to get purified protein to immunize the mouse, the recombinant vectors of pET-28a (+)-atp5b and pET-32a (+) -atp5b were expressed in E. coli BL21-DE3 and purified by Ni-NTA affinity chromatography. Figure 1 shows that ATP5B and ATP5B-Trx were successfully expressed and purified, respectively. In addition, the concentration of ATP5B and ATP5B-Trx were determined by BCA assay as 217.43 and 426.62 μg/mL, respectively.
Expression and purification of the beta subunit of ATP synthase. All proteins were analyzed by 12% SDS-PAGE. (
Determination of anti-serum titer
In this study, ATP5B-Trx fusion protein was used as antigen to immunize the mouse. ATP5B was adopted to coat protein to determine the anti-serum titer using indirect ELISA. A detection system was set up containing an optimal coating concentration of antigen (4.35 μg/mL) and an optimal diluted concentration of the second antibody (1:20,000). Figure 2 shows that the immunized mice 1A, 2A, and 2B generated a higher positive titer among the six mice (1A, 1B, 2A, 2B, 3A, 3B). Mouse 2A had the highest anti-ATP5B titer, so the spleen cells of the 2A mouse was subsequently chosen for cell fusion.
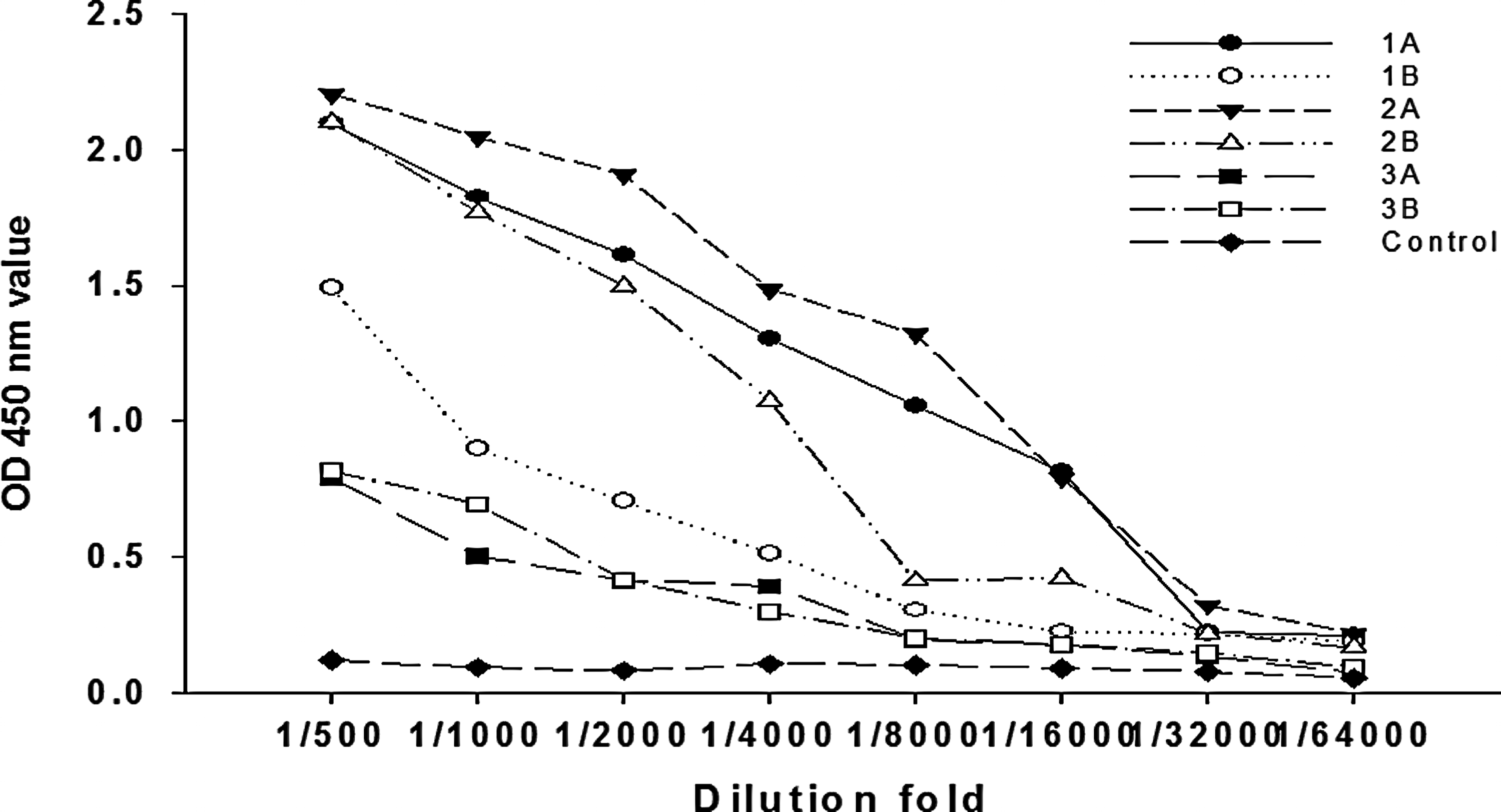
Determination of anti-serum titer by indirect non-competitive ELISA. After 7 d from the third immunization, the mice serum was determined by indirect ELISA.
Cell fusion and screening of hybridoma clones
After being fused for 4 d, two or six cells were prone to cling together. These cells were likely successful fusion hybridoma. After 6 d, dead cells were weeded out by renewing the medium and the fusion cell numbers increased greatly. The bottom of 96-well microtiter plates were covered with hybridoma clones after 10 d. At the same time, the supernatant titer was determined by indirect non-competitive ELISA. The fusion rate of cells reached 72.53%, while the positive rate of fused cells was 29.78%. These results indicated that the spleen cells of the ATP5B immunized mice and myeloma cells were fused successfully. Among these positive clones, it is worth noting that 6G11 was significant. Subsequently, the subcloning of the 6G11 hybridoma clone was carried out by limiting dilution method. After subcloning, the positive rate of 6G11 hybridoma clone reached 100% (Fig. 3).

The subcloning of 6G11 hybridoma cells via microscopy. (
Identification of hybridoma clone
In order to test the ability of 6G11 to secrete monoclonal antibody, SDS-PAGE and chromosome counts were performed. The results showed that the 6G11 hybridoma clone had a band of heavy chain at 50 kDa and a band of light chain at 27 kDa (Fig. 4). It was coincident with the component of an actual monoclonal antibody.

SDS-PAGE analysis of supernatants antibody of 6G11 hybridoma cell. Lane 1, protein maker; lane 2, unpurified antibody from cell culture supernatant.
The chromosome count showed that the chromosome numbers of SP2/0 cells and spleen cells were about 60 and 40, respectively. The number of 6G11 hybridoma cells was from 98 to 102, which was the sum of a spleen cell and a SP2/0 cell (Fig. 5), indicating that the hybridoma was coming from SP2/0 cells and spleen cells of the immunized mouse.
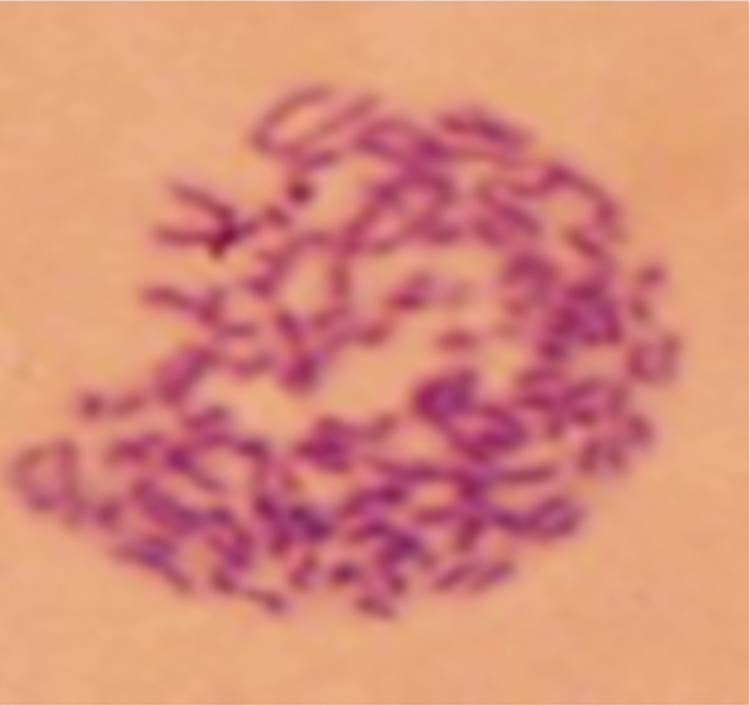
Chromosome staining of 6G11 hybridoma cell.
Cross-reactivity assay
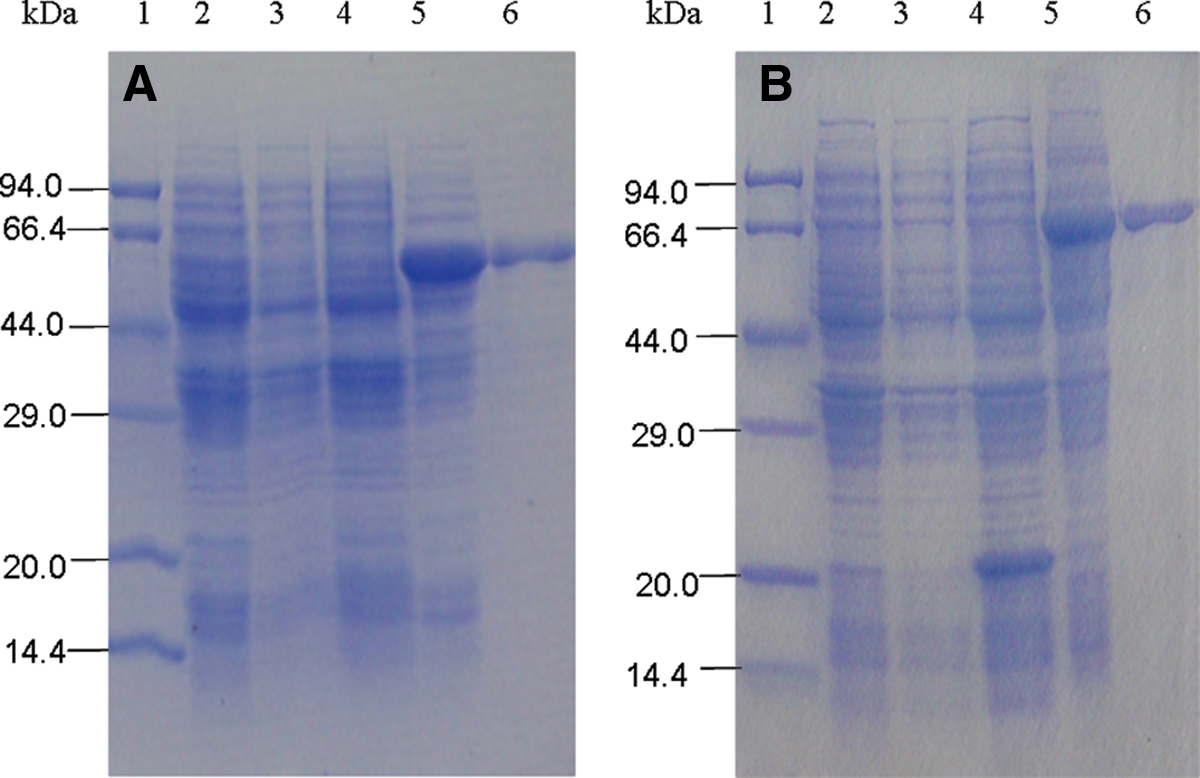
To determine the specificity of the monoclonal antibody, Western blot analysis was performed (Fig. 6). It was observed that protein bands of GST, SIRT5, and IDH3α (negative control) were on SDS-PAGE (Fig. 6A) but not on Western blots (Fig. 6B). Band of ATP5B protein was observed on SDS-PAGE (Fig. 6A) as well as on Western blots (Fig. 6B). This result revealed that the monoclonal antibody reacted with the ATP synthase β-subunit, but not with GST, SIRT5, and IDH3α. Based on this, it was concluded that the hybridoma clone was a specific monoclonal antibody against ATP5B.
Analysis of specificity of anti-ATP5B antibody via Western blot analysis. (
Determination of titer and Ig subclass of anti-ATP5B antibody
The titer of the monoclonal antibody was determined by indirect non-competitive ELISA. The titer was defined as the dilution times when the absorbance value was twice the control. The results showed that the titer of the supernatant of 6G11 reached 1:1600. After 120 d, the absorption value at 450 nm changed little, so the titer of the monoclonal antibody had high stability.
The hybridoma clone was identified by an Ig subclass assay kit. The results are shown in Table 1, indicating that 6G11 belonged to IgG1 subclass.
Determination of relative affinity constant of anti-ATP5B antibody
The affinity constant produced by 6G11 hybridoma was measured by non-competitive enzyme immunoassay using serial dilutions of both coated antigen and MAb. Calculated by a formula, the result showed that the relative affinity of the 6G11 was about 3.1×109 L/mol.
Discussion
In this study, a hybridoma clone that secreted monoclonal antibody against ATP5B antigen was prepared by cell hybridization technique. There were some issues confronted during the production of the anti-ATP5B antibody. The first obstacle was how to increase the immunogenicity of ATP5B that existed in mice naturally. In the study, two prokaryotic recombinant vectors were constructed to express ATP5B-Trx and ATP5B, which were used to immunize mice and to test the production of antibody, respectively. The second challenge was how to increase the fusion rate and the positive rate being key indicators to evaluate cell fusion. In the experiment, the SP2/0 cell with the best growth state was chosen, and the ratio of the fused myeloma to spleen cell was 1:5. In addition, it was necessary to establish a test system by indirect ELISA that contained the optimal concentration of antigen and antibody.
SDS-PAGE and chromosome analysis were used to analyze the clone, and it is demonstrated that this clone was a hybridoma. By Western blot analysis, the hybridoma clone reacted with the β-subunit of ATP synthase, but not with GST, SIRT5, and IDH3α. In addition, the antibody titer of the hybridoma clone was determined by indirect non-competitive ELISA, and the 6G11 clone reached up to 1:1600. This would be a low titer for polyclonal antibody, but for a monoclonal antibody, the titer was sufficient. Ig subclass assay found that it belonged to IgG1 antibody. Based on these data, we concluded that this hybridoma clone produced a specific MAb against ATP5B antigen. Compared with the previously reported MAb with an affinity of about 405 nmol/L (2.47×106 L/mol),(26) the relative affinity constant of 6G11 was much higher. Because of its high affinity, specificity, and stability, this MAb will be a much more helpful tool in the future.
Footnotes
Acknowledgment
This work was supported by the Doctoral Discipline Foundation for Young Teachers in the Higher Education Institutions of Ministry of Education, China (no. 20113515120001), the Science and Technology Foundation of Education Office of Fujian Province, China (no. JA10101), and the Natural Science Foundation of Science and Technology Department of Fujian Province, China (no. 2011J05049).
Author Disclosure Statement
The authors have no financial interests to disclose.