
Abstract
Glucosyltransferase-B (GTFB) of Streptococcus mutans is considered a virulence factor because of its activity in the production of insoluble glucan, which is key to the bacterial attachment onto dental surfaces, leading to the formation of dental caries. Local passive immunization with monoclonal antibodies against GTFB is considered to be an effective way to prevent dental caries. Here we amplified a 1.3 kb fragment of the N-terminal half of the gtfB gene (193–1530) of S. mutans by PCR and expressed the truncated protein (GTFBN). The expressed, purified protein was used as an immunogen in BALB/c mice. We selected and established one hybridoma (HBN8) that was capable of producing anti-GTFBN using ELISA, dot blot, and Western blot analyses. The monoclonal anti-GTFBN antibody was purified by affinity chromatography and its isotype was confirmed as IgG2a. The anti-GTFBN antibody inhibited the enzymatic activity of crude glucosyltransferase of S. mutans GS-5 in a dose-dependent manner. These data suggest that the anti-GTFBN antibody could be used as a vaccine to prevent the aggregation of S. mutans on tooth surfaces, and thus prevent the formation of dental caries.
Introduction
The virulence factors of S. mutans include three glucosyltransferases (GTFs): GTFB (insoluble glucan, 162 kDa), GTFC (insoluble and soluble glucan, 149 kDa), and GTFD (soluble glucan, 155 kDa).(14–17) Monoclonal antibodies against GTFs have been used to study the functions of these enzymes and their role in cariogenicity.(18–22) GTFB and GTFC primarily synthesize water-insoluble glucans, which contribute to the initiation of caries on smooth surfaces and plaque formation.(23,24) These GTFs catalyze the production of adhesive glucans from sucrose, which enhances bacterial colonization on tooth surfaces and promotes the formation of dental plaque, leading to demineralization of the enamel surface.(14,17,23,24) For these reasons, GTFs are considered good targets for anti-caries vaccines. GTFB is an especially important factor in human cariogenesis.(25,26) Several studies of the structure-function relationships of the GTFs of S. mutans and S. sobrinus have revealed that amino acids in the N-terminus of GTFs may play a central role in sucrose splitting and glucan synthesis, while amino acids in the C-terminus are responsible for glucan binding.(14,19,27,28) A previous study showed that the inhibition of insoluble glucan synthesis results in reduced bacterial colonization and cariogenicity.(29) Therefore, we focused on the N-terminal fragment of the gtfB gene to express the GTFBN protein, which was used as an immunogen to generate an anti-GTFBN monoclonal antibody. We investigated the inhibitory activity of the monoclonal antibody against the formation of insoluble glucans that are important in S. mutans and other oral bacteria for bacterial tooth surface attachment and the formation of dental plaque.
Materials and Methods
Construction of GTFBN expression vector
Approximately 1.3 kb of the N-terminal fragment of gtfB gene (gtfBN, 193–1530) was amplified by PCR using the primers gtfBN-F (5′ GTA AGA GCT CAT AAC CGA AGC GAC ATC TAA 3′) and gtfBN-R (5′ GAT AGT CGA CCG CAG CTT ATT GTC CAT ATT 3′) in a 50 μL mixture containing 1 μL of the template DNA (∼20 ng), 5 μL of 10X Taq buffer, 0.5 μL of Taq polymerase (5 U/μL, GeNet Bio, Nonsan, Korea), 3 μL of the 2.5 mM dNTP mixture, and autoclaved distilled water. The reaction was a preliminary denaturation at 95°C for 5 min, followed by 30 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min, and extension at 72°C for 1 min, and a final extension at 72°C for 5 min. The PCR fragment was subcloned into a T vector (Real Biotech, Taiwan), and the sequence was confirmed. SalI and SacI sites in the T vector were cut, and the gtfBN gene was inserted into the SalI and SacI sites of the pET28a(+) vector (Novagen, Madison, WI).
Bacterial strains and growth conditions
The pET28a(+) vector with the inserted gtfBN gene (pGTFBN) was transformed into Escherichia coli BL21 cells and was cultured overnight at 37°C in 2 mL of LB broth containing kanamycin (50 μg/mL). For the preparation of crude GTFs, S. mutans GS-5 was inoculated into 2 mL of brain heart infusion (BHI) broth and cultured overnight at 37°C. The following day, 100 μL of S. mutans GS-5 was transferred into 1 L of BHI broth and cultured overnight at 37°C.
Expression and purification of GTFBN protein
The 2 mL culture of BL21 containing pGTFBN was transferred into 200 mL LB broth with kanamycin (50 μg/mL) on the following day and incubated at 37°C. When the culture reached an OD of 0.6–0.8, expression of the gene was induced by adding isopropylthio-β-D-galactoside (IPTG, 0.8 mM) at 28°C for overnight incubation. The culture was centrifuged the following day at 5000 g for 10 min, and the pellet was resuspended in an 8 M urea lysis buffer and agitated overnight in a shaking incubator at 28°C. The culture was subsequently centrifuged at 10,000 g for 15 min, and the cleared lysate was loaded onto a Ni-NTA column (Qiagen, Valencia, CA) equilibrated with 8 M urea lysis buffer. The column was washed twice with an 8 M urea wash buffer and protein was eluted with elution buffer. The size of the eluted GTFBN protein (about 70 kDa) was confirmed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The eluted protein was dialyzed in a dialysis tube in distilled water for 24 h, freeze-dried, resuspended in phosphate buffered saline (PBS), and stored at −20°C until further use.
Immunization
Four-week-old female BALB/c mice (Damul, Daejeon, Korea) were purchased and raised for 2 weeks before injection. A homogeneous emulsion of Freund's complete adjuvant (Sigma Chemical Co., St. Louis, MO) and GTFBN protein (about 80 μg in PBS) was intravenously injected at a 1:1 volume ratio. Two weeks after the first injection, a booster of Freund's incomplete adjuvant with GTFBN was performed subcutaneously, and blood was collected from each mouse one week after the second immunization. Serum obtained from the blood of the mice was screened at a 1:1000 dilution by Western blot analysis against the GTFBN protein (10 μg/mL in PBS) and stored at −20°C until further use. Mice exhibiting the highest antibody titer were subcutaneously administered a third immunization (80 μg in PBS) with the antigen emulsified in Freund's incomplete adjuvant. The procedures for the experimental use of animals were approved by the Institutional Animal Care and Use Committee of the Chonbuk National University (approval no. CBU 2010-0028), and the guidelines suggested by the committee were followed.
Generation of hybridomas and monoclonal antibodies expressing the anti-GTFBN antibodies
On the sixth day after the third injection, spleen cells were collected from the mice and fused with SP2/0 myeloma cells using polyethylene glycol 1500 (PEG 1500, Roche Diagnostics, Mannheim, Germany) at a cell ratio of 4:1. After a series of fusion steps, the pellet of the fused cells was diluted in Dulbecco's modified Eagle's medium (DMEM, GIBCO Laboratories, Grand Island, NY) containing hypoxanthine-aminopterin-thymidine (HAT) supplemented with 18% fetal bovine serum. The medium was plated onto 96-well microtiter plates (SPL, Pocheon, Korea), cultured for 10 to 15 days, and the culture supernatants assayed for the presence of GTFBN-specific antibodies by ELISA. The findings were confirmed by dot blot and Western blot analyses. GTFBN-specific hybridomas were cloned once at 10 cells/well in the same 96-well plates, then cloned three more times at 1 cell/well.
ELISA
The levels of specific anti-GTFBN antibody activity were determined by an enzyme-linked immunosorbent assay (ELISA) in 96-well flat-bottomed polystyrene microtiter plates (Nunc, Roskilde, Denmark). The wells were coated with GTFBN protein (50 ng/well in a coating buffer of 0.05 M sodium carbonate and 0.05 M sodium bicarbonate) and incubated overnight at 4°C. After incubation, 200 μL of the blocking agent (5% skim milk) was added followed by further incubation for 40 min at room temperature (RT). The wells were washed three times for 5 min each with TBST between reactions. The media supernatant that contained the primary antibody (anti-GTFBN antibody) to be tested was added and incubated for 2 h at RT, followed by incubation with the secondary antibody, alkaline phosphatase-labeled goat anti-mouse immunoglobulin G (Sigma), for 30 min. The titer of the anti-GTFBN antibody was detected with the substrate of p-nitrophenyl phosphate (Sigma) and read at 405 nm using an ELISA reader (Packard Instrument, Downers Grove, IL).
Dot blot analysis
The presence of the anti-GTFBN antibody was confirmed by dot blot analysis. GTFBN (100 ng in 1 μL) was spotted onto a nitrocellulose membrane and dried. The membrane was soaked in 5% skim milk for 40 min at RT and was washed three times with TBST between reactions. The membrane was incubated with secondary antibody at RT for 30 min. After washing four times with TBST, the presence of the anti-GTFBN antibody was detected with TMN (100 mM Tris [pH 9.5], 100 mM NaCl, and 5 mM MgCl2) containing 66 μL of nitro blue tetrazolium (NBT) and 33 μL of 5-bromo-4-chloro-3-indoyl phosphate (BCIP).
Western blot analysis
The GTFBN protein was separated using SDS-PAGE and transferred onto nitrocellulose membrane. The membrane was blocked with 5% skim milk for 40 min at RT and washed three times for 5 min each with PBST. The membrane was then soaked in media supernatant for 2 h at RT and washed three times. The positive control was serum at a 1:1000 dilution (100 μg/mL in PBS). The membrane was treated with the secondary antibody, alkaline phosphatase-labeled goat anti-mouse immunoglobulin G (Sigma) for 1 h and then washed three times for 5 min each with PBST. The anti-GTFBN antibody was detected with TMN containing NBT and BCIP.
To confirm the existence of GTFB in crude GTFs, 10 μg of crude GTFs were separated by SDS-PAGE and transferred onto a nitrocellulose membrane. The membrane was blocked in 5% skim milk for 30 min at RT and washed three times for 5 min each with PBST. The membrane was then soaked in media supernatant for 2 h at RT and washed three times. The positive control was serum at a 1:1000 dilution (100 μg/mL in PBS). The membrane was treated with the secondary antibody, horseradish peroxidase-conjugated goat anti-mouse immunoglobulin G (Santa Cruz Biotechnology, Santa Cruz, CA) for 1 h, then washed twice with distilled water (DW) and three times with PBST for 10 min each. The anti-GTFBN antibody was detected with enhanced chemiluminescence (ECL, Intron, Sungnam, Korea).
Monoclonal antibody purification and determination of the antibody isotype
Monoclonal antibody (MAb) in the media supernatant of HBN8 was purified using a protein A sepharose column (Sigma). The column was washed with PBS (pH 7.2) to remove air bubbles and was filled with protein A beads. The column was then washed with PBS (pH 7.2) before the sample was loaded. The column was washed again with PBS (pH 7.2), and the IgG was eluted with elution buffer (0.1 M citric acid, pH 2.75). The isotype of the antibody produced from the stabilized hybridoma cell line was determined using an isotyping kit (Sigma) according to the manufacturer's instructions.
Reactivity of purified MAb
The purified IgG was tested for its reactivity with the N-terminus of the GTFB protein using an indirect ELISA. A 96-well plate was coated with 100 ng of GTFBN, GTFCN, GTFDN, Ag I/II protein, and crude GTFs obtained from S. mutans GS5 at 4°C overnight. The concentration of the proteins was determined by Bradford protein assay. The wells were blocked with 3% skim milk at RT for 30 min, followed by the addition of 0.5, 1.0, or 1.5 μg of the purified MAb. The plate was incubated for 2 h at 37°C and washed three times with TBST for 5 min between reactions. This step was followed by incubation with the secondary antibody, alkaline phosphatase-labeled goat anti-mouse immunoglobulin G for 30 min at 37°C. The activity of the purified IgG was determined using p-nitrophenyl phosphate as a substrate, with spectrophotometric analysis at 405 nm using an ELISA reader.
Inhibition of anti-GTFB antibody against crude glucosyltransferases of S. mutans GS-5
To test the inhibitory activity of the anti-GTFBN antibody against GTFB from S. mutans GS-5, we measured the level of insoluble glucan. Crude GTFs were prepared according to the method described by Song and colleagues(30) with some modifications. In brief, 1 L of S. mutans GS-5 was cultured and centrifuged at 4000 rpm for 15 min. The pellet was sonicated with three 10 s pulses at 10 s intervals and centrifuged at 10,000 rpm for 10 min. The crude GTFs were precipitated from the supernatant by adding ammonium sulfate (60%) and stirring overnight at 4°C. The suspension was centrifuged the following day at 6000 rpm for 30 min, and the pellet was dialyzed against 0.01 M KH2PO4 (pH 6.8) at 4°C overnight. The reaction mixture to test the inhibitory activity of the anti-GTFBN antibody against glucosyltransferases consisted of 300 μL of crude GTFs, 200 μL of 0.01 M KH2PO4 (pH 6.8) containing 1 mM NaN3, and 500 μL of 0.01 M KH2PO4 (pH 6.8) containing 5% sucrose and 1 mM of NaN3. Incubation was at 37°C for 24 h. The control was 300 μL of crude GTFs, 200 μL of 0.01 M KH2PO4 (pH 6.8) containing 1 mM of NaN3, and 500 μL of 0.01 M KH2PO4 (pH 6.8) containing 1 mM of NaN3. Purified anti-GTFBN monoclonal antibody was added to the solutions at 0, 50, 100, 150, or 200 ng/mL. The concentration of the produced glucans was determined by measuring optical density at 550 nm.
Statistical analysis
The mean values and standard error of the mean (S.E.M.) were statistically analyzed using Origin 7 (Origin Lab, Northampton, MA). Data were expressed as the reciprocal of the geometric log2 of the least sample that gave an OD 550 of 0.07, which was the value of PBS. Data were considered statistically significant when the p value was <0.05.
Results
The N-terminal half of the gtfB gene produced a GTFBN protein of about 70 kDa, as determined by SDS-PAGE (Fig. 1). The purified GTFBN protein was used as an immunogen. Nineteen clones were initially selected based on the ELISA, dot blot, and Western blot analyses. After four repeated selection rounds, the clone that displayed the highest affinity was selected and was named HBN8 (Fig. 2). For the tests performed in this study, the MAb in the culture supernatant of HBN8 was purified using protein A sepharose (Fig. 3). The isotype of the MAB was IgG2b.
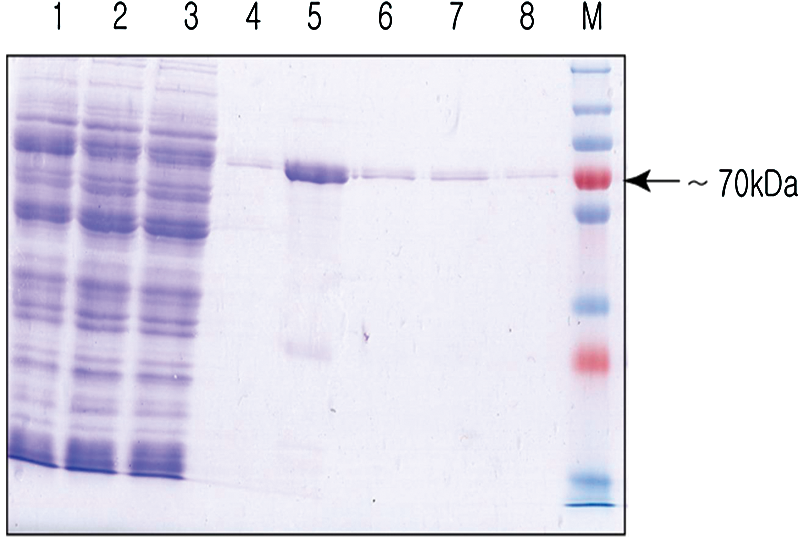
Purification of the recombinant GTFBN protein. Approximately 1.3 kb of the N-terminal fragment of the gtfB gene produced a protein with a molecular weight of about 70 kDa. The protein was induced at 28°C overnight and was purified with a Ni-NTA column. M, protein marker; lane 1, supernatant; lane 2, flow-through; lane 3, wash I; lane 4, wash II; lanes 5–8, eluants 1–4.
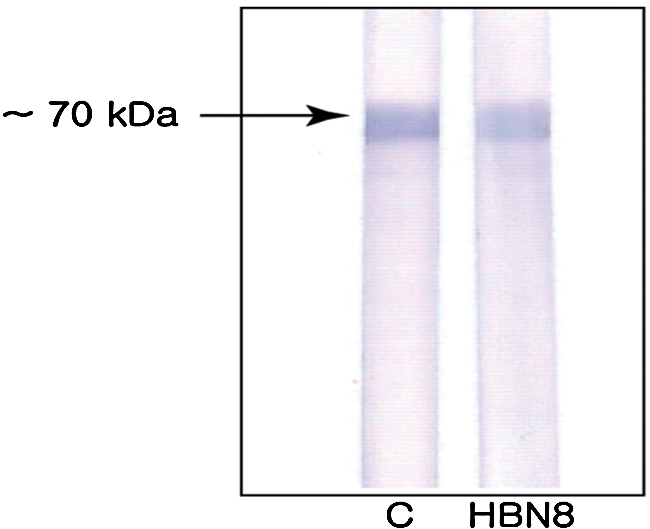
Western blot analysis of the final selection step. Five wells were selected. One well showed a strong band, which was named HBN8. Membrane 1, control (1:1000 dilution of the GtfBN positive serum in PBS); membrane 2, HBN8.
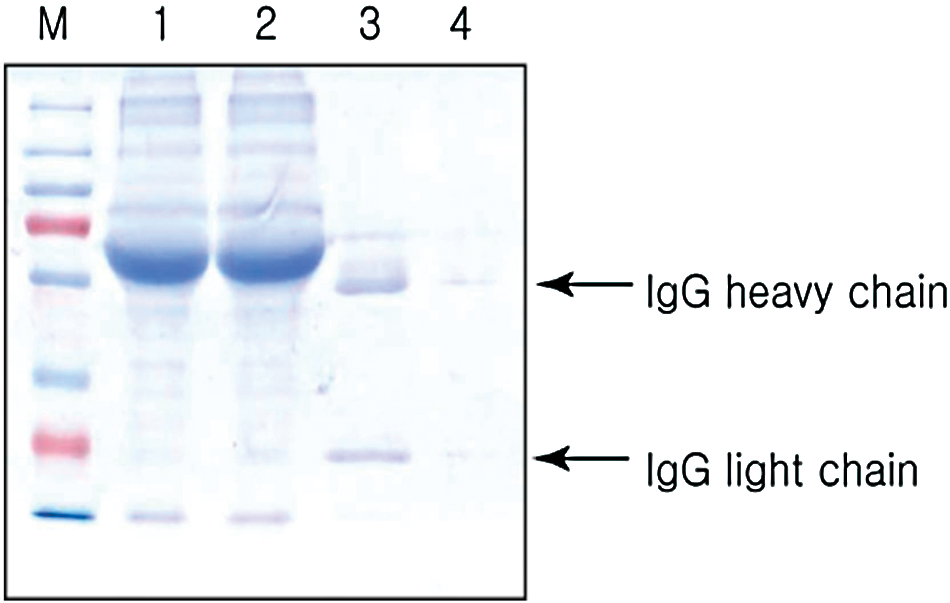
Purification of anti-GtfBN monoclonal antibody. Anti-GtfBN MAb was purified from culture supernatant with protein A sepharose. M, protein marker; lane 1, culture supernatant; lane 2, flow-through; lanes 3–4: eluants 1–2.
The specific reaction of the monoclonal antibody produced from HBN8 in the media supernatant with recombinant BN was compared to its reaction with recombinant CN (not published data), DN,(31) Ag I/II protein,(32) and crude GTFs and confirmed by ELISA (Fig. 4). The purified MAb was found to specifically bind to GTFBN protein and interact slightly with GTFCN, but not with other proteins. To determine the inhibitory activity of the MAb against glucosyltransferase, we prepared crude GTFs of S. mutans GS-5. The result showed that the MAb reduced the activity of the crude GTFs in a dose-dependent manner (Fig. 5), and the reduction was significant at 200 ng of the monoclonal anti-GTFBN antibody (p<0.05).
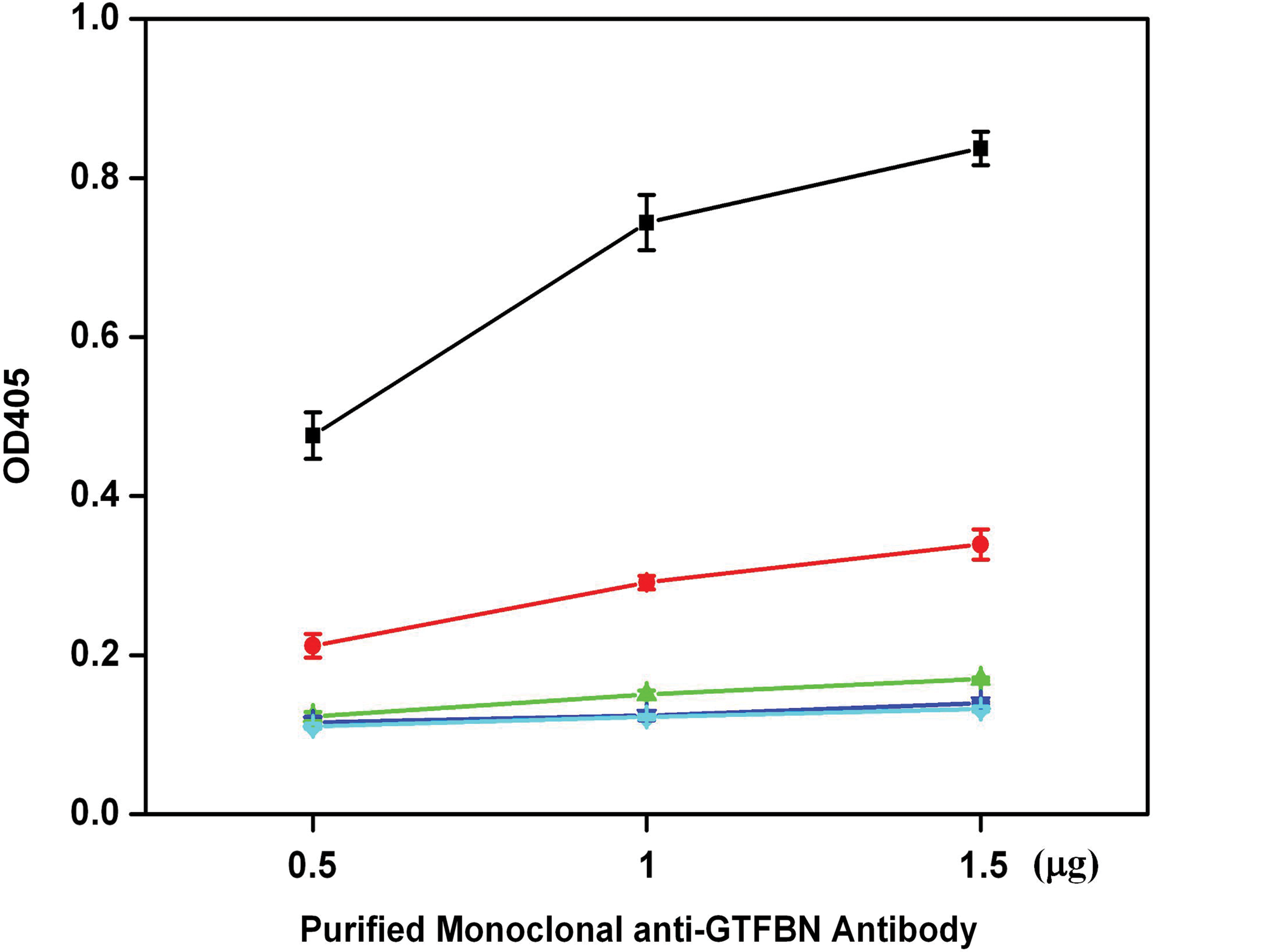
Reactivity of MAb IgG of HBN8 to components of S. mutans GS-5. The MAb (0.5, 1.0, or 1.5 μg) was examined for its reactivity to GTFBN (■), crude GTFs (•), GTFCN (▲), GTFDN (▼), and Ag I/II (♦). Data represent the mean±SD of triplicate assays.
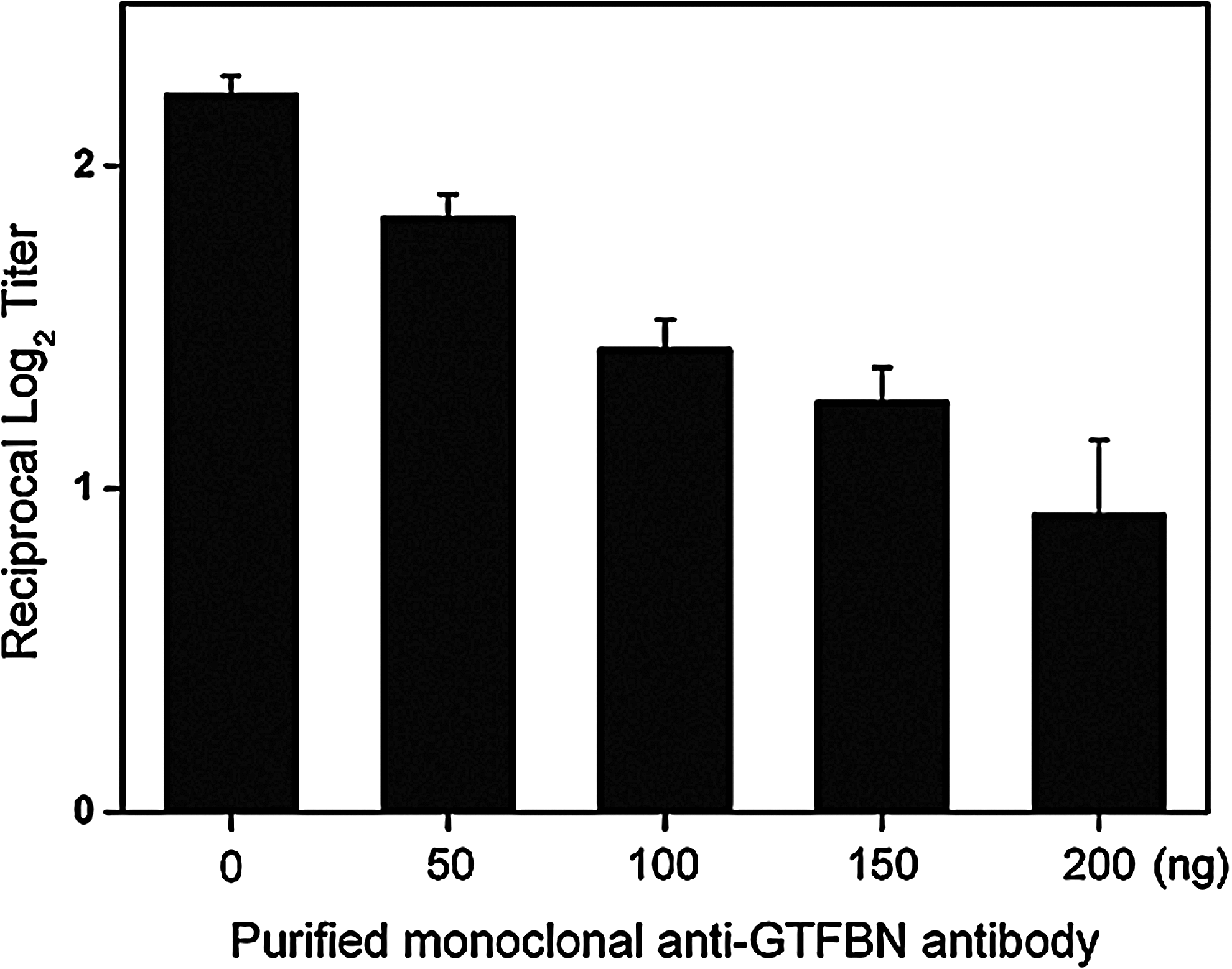
Inhibitory activity of anti-GTFBN monoclonal antibody against the crude glucosyltransferases of S. mutans GS-5. Purified anti-GTFBN MAb was added to a solution of 0.01 M KH2PO4 (pH 6.8) with or without 5% sucrose at 0, 50, 100, 150, or 200 ng/mL. The optical densities of the produced glucans were measured at 550 nm. Data represent the mean±SD of triplicate assays.
Discussion
Dental caries is the most prevalent and costly oral infectious disease.(33) Passive immunization against S. mutans is considered to be an effective means to avoid side effects of systemic immunization for the prevention of dental caries. The formation of dental plaque is initiated by insoluble glucans produced by S. mutans GTFB and GTFC, which provide binding sites for other oral bacteria. According to Koo and colleagues,(34) the function of the gtfB gene is crucial for clustering and stabilization of bacterial cells, which leads to the formation of cariogenic dental plaque. A number of studies have shown that antibodies to S. mutans inhibit the enzymatic activity of the bacteria.(9–13) However, they were to peptides of the virulence factors of S. mutans, to GTFs isolated from the bacteria, or to whole cell while we amplified the gtfB gene by PCR and expressed to produce the GTFBN protein. The GTFBN recombinant protein was used as an immunogen to obtain a hybridoma cell line (HBN8) that produced an anti-GTFBN antibody. To be used as a vaccine through passive immunization, the monoclonal antibody must have the ability to inhibit the glucan-producing activity of glucosyltransferase. For this test, crude GTFs were prepared as a mixture of proteins secreted from S. mutans GS-5. Less polysaccharide was produced in the presence of the MAb compared with the control. We thus concluded that the MAb inhibited the activity of GTFB and some amount of GTFC in crude GTFs, since GTFB shares some homogeneity with GTFC, and the MAb interacted slightly with GTFCN by Western blot analysis (data not shown).
In conclusion, we established a hybridoma cell line, HBN8, that produced MAb against the GTFBN protein of S. mutans GS-5. This MAb was capable of inhibiting the glucan-producing activity of crude GTFs in a dose-dependent manner, demonstrating its potential as a vaccine candidate for preventing the formation of dental plaque and dental caries. We are currently investigating its use to prevent the formation of biofilms by S. mutans. Further studies on the inhibitory activity of this monoclonal antibody against purified GTFB should be conducted.
Footnotes
Acknowledgment
This study was supported by a grant (no. 1103000725) from the National Research Foundation of Korea and by a research fund of Chonbuk National University in 2010.
Author Disclosure Statement
The authors have no financial interests to disclose.