
Abstract
B and T lymphocyte attenuator (BTLA) is recently identified as the third co-inhibitory receptor with similarities to CTLA-4 and PD-1. Previous reports have shown that BTLA is associated with autoimmune diseases, viral infections and tumor immune evasion. However, the possibility of the existence and role of a soluble form of human BTLA (sBTLA) has not been revealed. Based on our previously generated mouse anti-BTLA monoclonal antibodies, we intend here to develop a novel enzyme-linked immunosorbent assay (ELISA) for detecting sBTLA. Using monoclonal (MAb) 8H9 as coated antibody and the biotin-labeled MAb 7D7 for detection, a sandwich ELISA was developed with good sensibility, line reliability, and specificity. With the established ELISA, the existence and concentration of sBTLA were demonstrated for the first time. It was found that soluble sBTLA existed in the sera of healthy donors and the quantitation of sBTLA climbed along with increasing age, indicating sBTLA may be correlated with immune dysregulation resulting from the aging. Hence, a sandwich ELISA for detecting sBTLA was successfully established and soluble BTLA existing in human serum was first identified, which might play an important role in immunoregulation.
Introduction
BTLA is the most recently represented member of the immunoglobulin superfamily of co-inhibitory receptors(14); it shares an unexpected ligand with LIGHT, LT-α, and HSV1 gD, HVEM, which belongs to the TNFR superfamily.(15,16) In vitro engagement of BTLA on T lymphocytes with HVEM or agonistic monoclonal antibody could repress anti-CD3-mediated T cell proliferation, down-regulated CD25 expression, and inhibited IL-2 production.(17,18) The mechanism of BTLA suppressing antigen-driven T cell proliferation was revealed to lie in that cross-linked BTLA initiated in its cytoplasmic tyrosine phosphorylation and associated with the tyrosine phosphatase SHP-1 or SHP-2 to(14,15,19) In vivo the mice deficient in BTLA expression (BTLA-/-) was employed in addressing BTLA function. Original study on BTLA revealed that the sensitivity to experimental autoimmune encephalomyelitis (EAE) increased in BTLA-/-mice(14) and following research also reported that BTLA-/- mice spontaneously develop autoimmune hepatitis-like disease, suggesting that BTLA may have an important role in preventing autoimmune diseases.(20,21)
Accumulating evidence indicates that soluble forms of co-inhibitory receptors contribute to autoimmune disease. For example, increased production of soluble CTLA-4 (sCTLA-4) correlated with autoimmune thyroid disease,(22,23) and soluble PD-1 (sPD-1) occurred at high concentrations in sera of patients with rheumatoid arthritis (RA) and correlated significantly with titers of rheumatoid factor in RA patients.(24) Previously, we produced four monoclonal antibodies (1F11, 3B6, 7D7, and 8H9) against human BTLA recognizing at least three epitopes,(25) which laid the foundation for developing an ELISA kit for detecting soluble form of BTLA (sBTLA). Since the existence and potential role of sBTLA in physiological and pathologic conditions are still unexposed, we here characterized the establishment of a specific ELISA for detecting sBTLA to evaluate its levels in human sera from healthy donors. Moreover, our initial observation described in this study revealed sBTLA expression in healthy donors and identified that the quantitation of sBTLA in the healthy donors correlated with increasing age. Hence, the establishment of sandwich ELISA for detecting sBTLA laid the foundation for studying the expression and function of sBTLA on disease development.
Materials and Methods
Antibodies and proteins
PE-conjugated goat anti-human IgG was purchased from Immunotech (Marseilles, France). Available mouse anti-human BTLA (clones 7D7 and 8H9) was generated previously.(25) Streptavidin–horseradish peroxidase (HRP) was purchased from Sigma (St. Louis, MO). Goat anti-human BTLA 330104, recombinant human IgG1-Fc, and recombinant human PD-1-Fc, PD-L1-Fc, and B7-H3-Fc fusion proteins and anti-human HVEM clones were all purchased from R&D Systems (Minneapolis, MN).
Cell preparation and culture
Human PD-1, PD-L1, HVEM, and empty vector-transfected L929 cell lines (L929/PD-1, L929/PD-L1, L929/HVEM, and L929/mock) were constructed in our laboratory. These cell lines were cultured in RPMI1640 medium or standard DMEM (Gibco, Carlsbad, CA) supplemented with 10% fetal calf serum (Hyclone, Logan, UT), 100 U/mL penicillin, 100 mg/mL streptomycin, and 2 mM L-glutamine with a 5% CO2, 37°C incubator.
Generation and characterization of recombinant human BTLA-Fc fusion protein
The cDNA encoding human BTLA extracellular domain without signal sequence was amplified by PCR using the specific primers: an upstream primer with NheI restriction: 5′-AC
Analysis of reactivity and specificity of BTLA-Fc with HVEM by flow cytometry
The cell lines L929/mock, L929/PD-1, L929/PD-L1, or L929/HVEM (5×105 cells/test) were incubated, respectively, with purified BTLA-Fc fusion protein or human IgG1 for 30 min at 4°C and washed with PBS. PE-goat anti-human IgG as secondary antibody was added for another 30 min at 4°C. After being washed, the stained cells were analyzed by flow cytometry and the Beckman–Coulter's Expo32 MULTICOMP software. To evaluate the quality of the recombinant BTLA-Fc fusion protein, the binding reactivity of different concentrations of BTLA-Fc (1, 5, or 10 μg/mL) with L929/HVEM was also detected and compared with that of anti-human HVEM.
Western blot analysis
To study the specificity and reactivity of BTLA-Fc protein with anti-human BTLA, BTLA-Fc fusion protein and commercial PD-1-Fc and PD-L1-Fc protein were separated on 12% SDS-PAGE gels. The protein bands were transferred to nitrocellulose membranes according to the manual of semi-dry transfer (Bio-Rad, Hercules, CA). The membranes were blocked with PBS containing 0.2% Tween-20 and 5% non-fat dried milk, and the target bands were stained with the commercial anti-human BTLA 330104 (5 μg/mL). The membranes were visualized with HRP-labeled goat anti-rat IgG Ab and SuperECL Plus reagents (Applygen Technologies, Beijing, China). To identify the existence of sBTLA in human serum, several serum samples containing high concentrations of sBTLA detected by ELISA were analyzed by SDS-PAGE electrophoresis with or without the reducing agent β-mercaptoethanol and Western blot analysis, according to the same procedure as that described above, with BTLA-Fc as a positive control and mouse serum samples as the negative controls.
sBTLA ELISA and specificity assay
ELISA 96-well plates (Costar, Corning, NY) were pre-coated with 3μg/mL mouse anti-BTLA (clone 8H9) overnight at 4°C. Unoccupied binding sites on the plates were blocked with 2% BSA in PBS at 37°C for 2 h. Purified recombinant human BTLA-Fc fusion protein was used as a standard. Samples and standards were added to the wells for 2 h at 37°C in duplicate. Specific binding was detected with 1 μg/mL biotinylated mouse anti-BTLA (clone 7D7) for 1 h at 37°C, followed by HRP-streptavidin (Sigma) at 1:2000 for 1 h at 37°C and then revealed with the substrate TMB. The plates were washed five times with PBS containing 0.05% Tween-20 after each of these steps. Absorbance was measured at 450 nm by a microplate reader (Bio-Rad). The specificity of ELISA was determined by other homologous proteins in the same superfamily. The serial, two-fold dilutions of the soluble PD-L1-Fc, PD-L1-Fc, and B7-H3-Fc proteins starting from 20 ng/mL were detected by the ELISA to assess the specificity of the established system.
Human serum samples
All human serum samples were collected by the venipuncture technique from healthy donors, obtained with appropriate informed consent. Briefly, blood collected by venipuncture was allowed to clot for 30 min at room temperature for serum samples and then centrifuged for 10 min at 2500 rpm. The serum samples for the preliminary validation study were obtained from normal donors, ages 16 to 75 years. All the samples were grouped according to the donors' ages, as follows: 16–20 years old (n=23, male/female=10/13), 21–30 years old (n=52, male/female=32/20), 31–50 years old (n=25, male/female=12/13) and 51–75 years old (n=48, male/female=20/28).
Statistical analysis
Two-tailed p<0.05 were considered statistically significant. The bars shown for every group in Figure 3 represent the mean, and the number presented in Table 1 is the mean±standard error of the mean (SEM). The statistical software GraphPad Prism V5.0 (La Jolla, CA) was used for statistical tests.
Data are indicated as means and SEM of sBTLA in the serum of healthy donors of different ages (y, years). Significant differences of sBTLA between different ages is indicated with letters a1-3, b1-2, and c (a1, a2, and a3, compared with age 16–20; b1 and b2, compared with age 21–30; and c, compared with age 21–30). The p values are: a1, b1, and c, p<0.05; a2, a3, and b2, p<0.01.
Results
Preparation and characterization of recombinant human BTLA-Fc fusion protein
Recombinant human BTLA-Fc fusion protein was purified from the cultural supernatants of the transfectant CHO/BTLA-Fc, and the reactivity of binding to HVEM was determined by flow cytometry. Moreover, the characteristics of the product being recognized by anti-human BTLA s was analyzed by Western blot. The assay showed the purified product was not only bound to HVEM expressed on the transfectant L929/HVEM (Fig. 1A, B), but also specifically recognized by anti-human BTLA (Fig. 1C). Moreover, the reactivity of BTLA-Fc protein bound to L929/HVEM presents concentration dependent and is slightly less than that of anti-HVEM (Fig. 1B), suggesting that the recombinant BTLA-Fc fusion protein is of good quality and may be subsequently used for the standard curve of the sandwich ELISA.

Characterization of reactivity of recombinant human BTLA-Fc fusion protein with HVEM. (
Establishment of novel sandwich sBTLA ELISA and specificity analysis
For quantitative determination of sBTLA, a sandwich ELISA was established by using 8H9 as coated antibody and the biotin-labeled 7D7 for detection. The result of standard line indicated a good sensibility and line reliability of sBTLA ELISA method from 0.313 to 20.0 ng/mL. The R2 is >0.99 (Fig. 2A). The absorbance value of different soluble proteins showed the established ELISA was special to detect sBTLA, and the absorbance did not have any correlation with different concentrations of sPD-1, sPD-L1, or sB7-H3 recombinant protein (Fig. 2B).
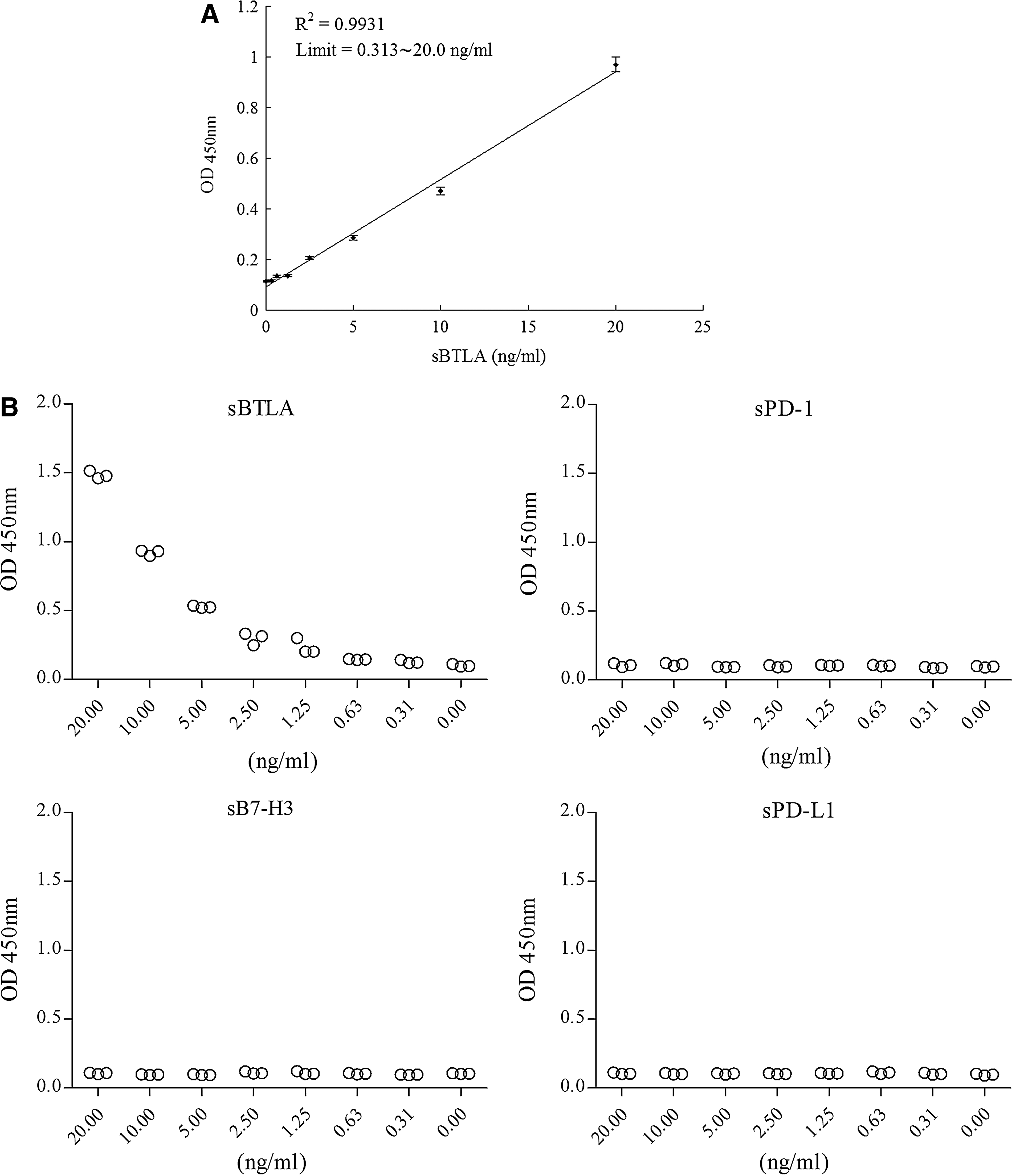
Establishment of sBTLA ELISA. (
sBTLA exists in serum of healthy donors and increases with age
The expression levels of sBTLA in the serum of healthy donors with different ages were determined by the established sBTLA ELISA. The results showed that sBTLA was expressed in normal human serum (Fig. 3). Interestingly, the levels of sBTLA in human serum increased with age. The group of 16–20 years old had the lowest level of sBTLA, the concentration being about 2.90±0.23 ng/mL, while the group of 51–75 years old had the highest level of sBTLA (6.32±0.35 ng/mL). The sBTLA levels of the two middle young-adult groups were 4.16±0.24 and 5.18±0.43 ng/mL, respectively (Table 1). sBTLA expression levels in human serum exhibited a distinct age-dependent manner.
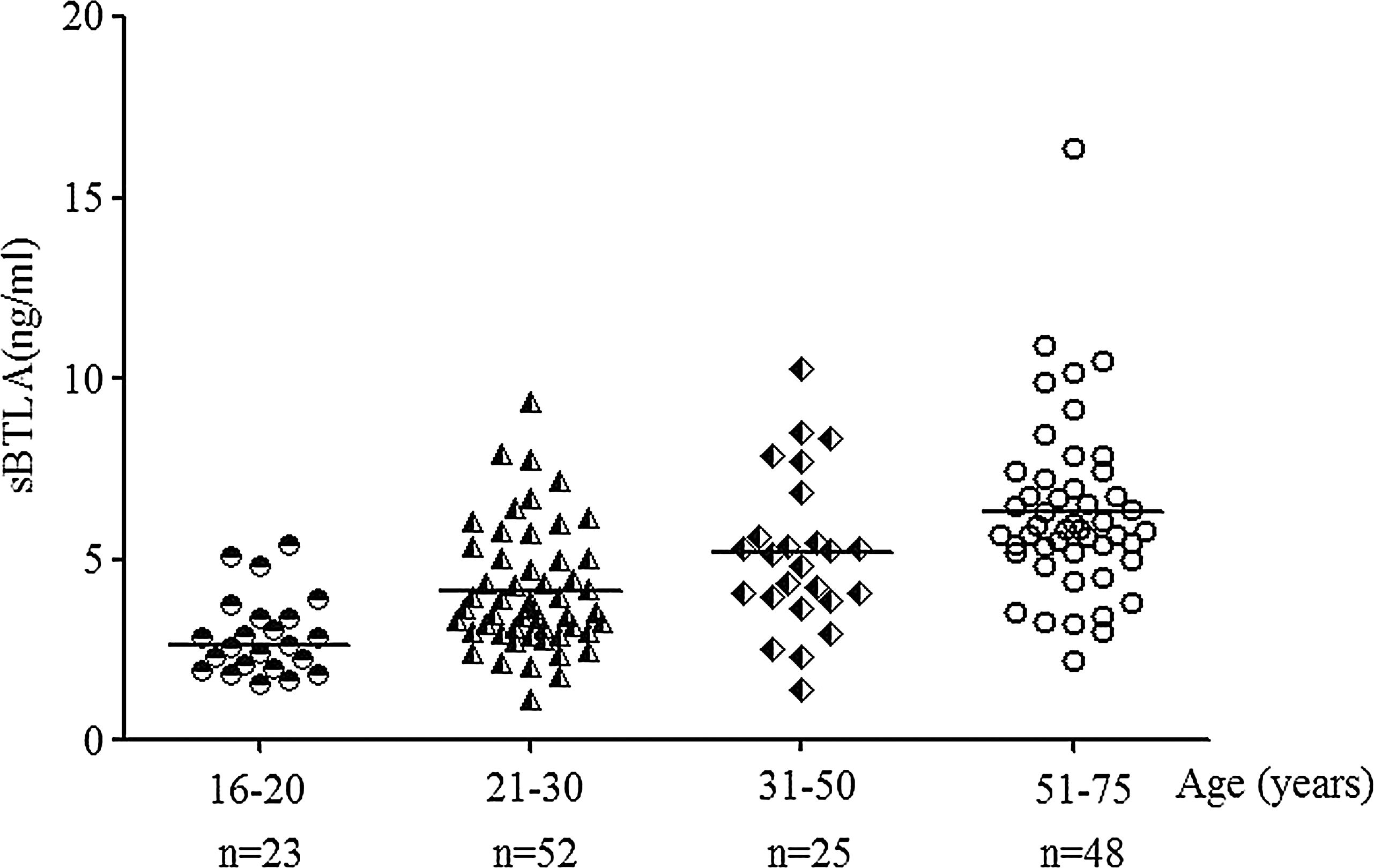
Expression levels of sBTLA in human serum from healthy donors of different ages. Sera from healthy donors were collected and grouped by age. Then the sBTLA was detected by the ELISA system. The level of sBTLA in the serum from 16- to 20-year-old donors was the lowest among the four groups, while the 51- to 75-year-old donors had the highest concentration of sBTLA. sBTLA expression level has a distinct age-dependent manner. The bar represents the mean value of every group.
To further identify the presence of sBTLA, several serum samples were analyzed by Western blot using the commercial anti-human monoclonal antibody. The results showed that the protein from the serum samples was detected with a molecular weight of about 45.5 kDa, which is less than that of BTLA-Fc protein (Fig. 4).
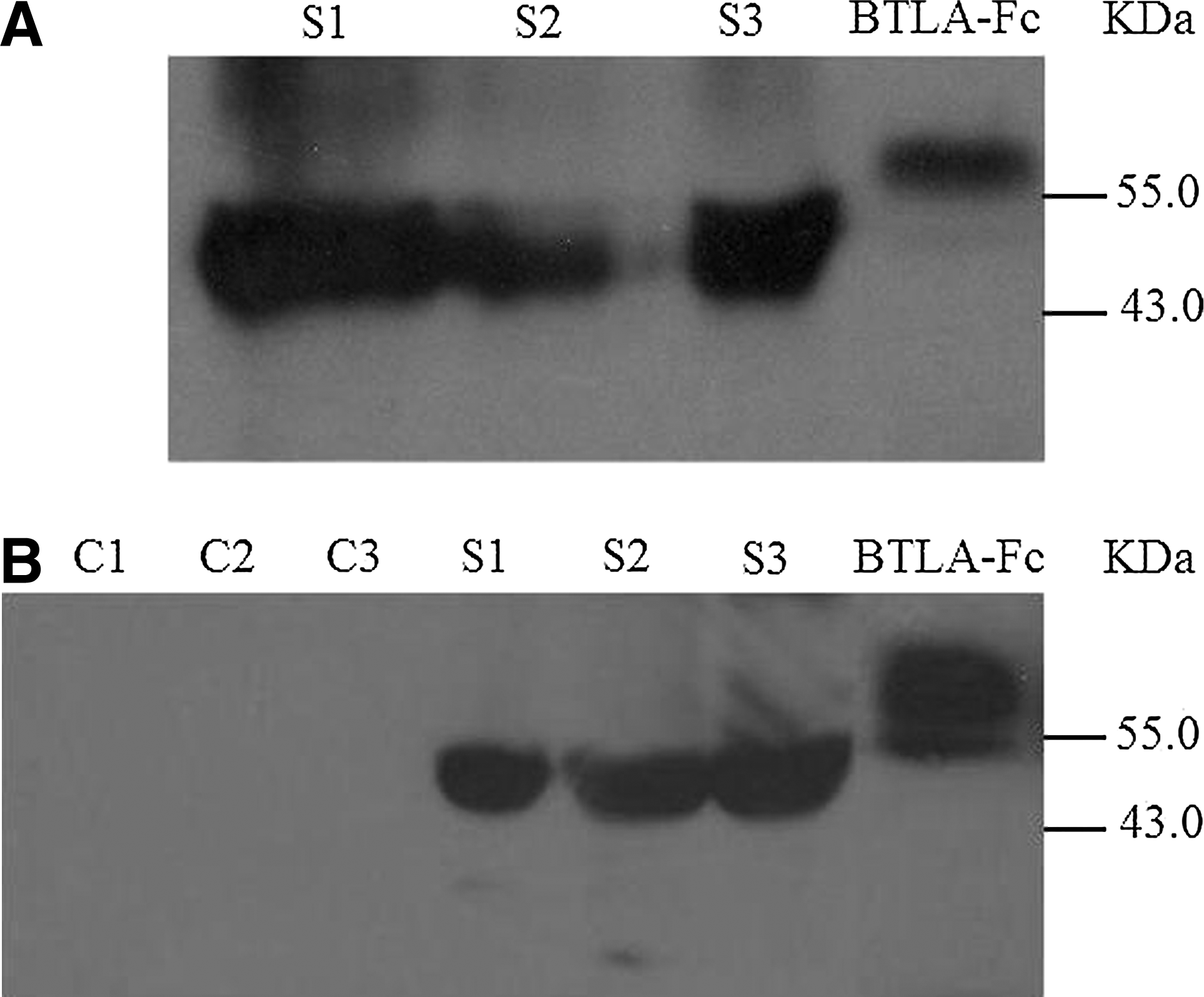
Western blot analysis of sBTLA from human serum samples. Purified BTLA-Fc fusion protein (positive control), three human serum samples containing high levels of sBTLA (S1∼S3), and three mouse serum samples (C1∼C2) were loaded on SDS–PAGE electrophoresis and transferred onto the nitrocellulose membranes. The membranes were incubated with the commercial anti-human BTLA (clone 330104) followed by goat anti-rat IgG-HRP. The SDS-PAGE was respectively run with (
Discussion
Mounting data demonstrated that soluble forms of co-signal molecules frequently exist in pathologic progress and correlate with disease development.(26–28) As soluble forms of co-inhibitory receptors, sCTLA-4 and sPD-1, which may be capable of interfering with their ligand interaction, were identified to be consistent with some specific autoimmune diseases.(22–24) Although BTLA-/- deficiency, as well as BTLA polymorphism, was confirmed to benefit autoimmune disorders,(20,21,29,30) whether sBTLA occurs and contributes to autoimmune damage is still not illustrated. Sensitive and specific ELISA was used widely to study soluble proteins in liquid sample. In this study, we described the development of a specific ELISA for the first time on the basis of our previously generated BTLAs as well as immediately produced BTLA-Fc protein. This ELISA kit for detecting BTLA-Fc was seen to obtain a good sensibility and line reliability sensibility.
With the established ELISA, sBTLA expression was detected in human sera from healthy donors, different from sCTLA-4 and sPD-1, as they were not detected in healthy persons.(24) The reason may lie in the different regulatory mechanism of expression between sBTLA and sCTLA-4, as well as sPD-1, just as that membrane-bound CTLA-4 and PD-1 are strictly regulated on activated T cells,(31–33) while membrane-bound BTLA is expressed on both naïve and activated T lymphocytes.(17,18,25,34) Furthermore, it was surprisingly shown that sBTLA concentration climbed with increasing age and presented the statistical significance of differences between investigated age ranges, indicating the elevation of this soluble co-inhibitory molecule in human sera may be consistent with human aging. As we know, the function of immune system declines with aging, which leads to increased frequency and severity of diseases. Whether sBTLA is correlated with immune dysregulation and has any aberrant impact on immune system of the elderly deserve further investigation.
Both sCTLA-4 and sPD-1 were derived from the isoform encoded by the alternative transcript, which lack the transmembrane region but contain complete extracellular domain.(24,35,36) Whether sBTLA comes from the isoform needs further study. However, sBTLA detected by Western blot has about 45.5 kDa of molecular weight, which is larger than the extracellular domain of BTLA, suggesting that it does not shed from the full-length membrane-bound BTLA by protease like other soluble co-stimulatory molecules do.(37,38)
In summary, a novel ELISA system was developed for detecting sBTLA, and our results originally demonstrated that sBTLA existed in normal human serum. However, the particular role of sBTLA in the pathological conditions of diseases requires further investigation.
Footnotes
Acknowledgments
This research was supported by the National Natural Science Foundation of China (no. 30600548), the National Key Natural Science Foundation of China (30930085), and the Qin Lan Project of Jiangsu Province and the Project of Scientific and technical development of Suzhou (no. SYS201101).
Author Disclosure Statement
The authors declare that they do not have commercial associations that might cause a conflict of interest in connection with this work.