
Abstract
Factor VII (FVII) is a serine protease-coagulating element responsible for the initiation of an extrinsic pathway of clot formation. Here we generated and characterized a high affinity monoclonal antibody that specifically recognizes human FVII. Recombinant human FVII (rh-FVII) was used for the production of a monoclonal antibody using BALB/c mice. The specificity of the antibody was determined by Western blot using plasma samples from human, mouse, sheep, goat, bovine, rabbit, and rat. Furthermore, the antibody was used to detect transiently expressed rh-FVII in BHK21 cell line using Western blot and sandwich ELISA. A mouse IgG1 (kappa chain) monoclonal antibody clone 1F1-B11 was produced against rh-FVII. The affinity constant (Kaff) of the antibody was calculated to be 6.4×1010 M−1. The antibody could specifically recognize an epitope on the light chain of hFVII, with no reactivity with factor VII from several other animals. In addition, transiently expressed rh-FVII in BHK21 cells was recognized by 1F1-B11. The high affinity as well as the specificity of 1F1-B11 for hFVII will facilitate the affinity purification of hFVII and also production of FVII deficient plasma and minimizes the risk of bovine FVII contamination when fetal bovine serum–supplemented media are used for production and subsequent purification of rh-FVII.
Introduction
The TF-FVIIa complex performs a proteolytic cleavage of factor X (FX) to form the activated protease FXa.(1) In association with active factor V (FVa), the FXa performs a proteolytic cleavage of prothrombin to form thrombin. Thrombin cleaves fibrinogen to fibrin, promoting formation of a fibrin clot and activates platelets for inclusion in the clot.(2,3)
Human factor VII contains four domains including N-terminal Gla domain, followed by two EGF-like domains, EGF 1 and EGF 2, and a catalytic domain. Through limited proteolysis of the Arg152-Ile153 peptide bond, factor VII is broken into the light and heavy chains.(4,5)
After the discovery and isolation of factor VII and its important role in coagulation,(6) studies were focused on the cloning, expression, and antibody production against human factor VII.(7,8) Different types of antibodies have been produced against factor VII for different purposes. In 1988, three different clones of mouse monoclonal antibody, namely RFF-VII/l, RFF-VII/2, and RFF-VII/3, were produced against different epitopes of human FVII.(9) Among them, the high affinity antibody clone RFF-VII/1, coupled to sepharose, was used to deplete factor VII from normal plasma leading to FVII deficient plasma.(10)
Upon activation of FVII, the structure of the protein changes, creating a novel antigenic epitope in the C-terminus of the light chain.(11,12) A rabbit polyclonal antibody has been raised against a 12-mer synthetic peptide derived from this novel epitope. This antibody is commercially used in enzyme-linked immunosorbent assays (ELISA) to detect FVIIa in organic samples.(13) In another study, monoclonal antibodies (MAbs) were used for recognizing the mutated site of a dysfunctional FVII variant that possessed a different FVII activity. By using an ELISA on patient samples with the monoclonal antibody that recognizes a specific epitope located in the three-dimensional structure in the first EGF-like domain of human FVII, the mutated site of a dysfunctional FVII variant was determined.(14)
One of the aims of antibody production is affinity purification of proteins.(15) In addition, the high specificity of monoclonal antibodies decreases background noises and cross-reactivity, providing reproducible results and ensuring efficiency in affinity purification. In this study, we have produced a monoclonal antibody with a high affinity to human factor VII. This antibody can be used for purification of recombinant human factor VII from supernatants of stably transfected cell lines as well as for producing human FVII-deficient plasma by depletion for research applications.
Materials and Methods
Immunization of mice and generation of hybridoma cells
To produce antibodies against FVII, BALB/c mice were immunized with a human recombinant factor VII (NovoSeven®, Novo Nordisk, Bagsvaerd, Denmark). Two mice were subcutaneously injected with 10 μg r-FVII plus 50 μL Immaccel (Pick Cell Laboratories, Amsterdam, Netherlands) plus 60 μL complete Freund's adjuvant. In the second and third injections, incomplete Freund's adjuvant was used and boosted similarly at weeks 2 and 4 with a 4-week interval using 20 μg of factor VII. After 21 days, blood samples were collected. Serum level of the antibody was measured by ELISA, as described below. The mouse with a higher antibody titer was selected for fusion (Fig. 1). Four days before fusion, the selected mouse was treated by intraperitoneal injection of 30 μg r-FVII plus 50 μL IMMACCEL and 80 μL 1.3% Alhydrogel (Alum, Accurate Chemical & Scientific Corp., Westbury, NY). One day before performing fusion, 20 μg r-FVII in 50 μL of phosphate buffered saline (PBS) was intravenously injected.
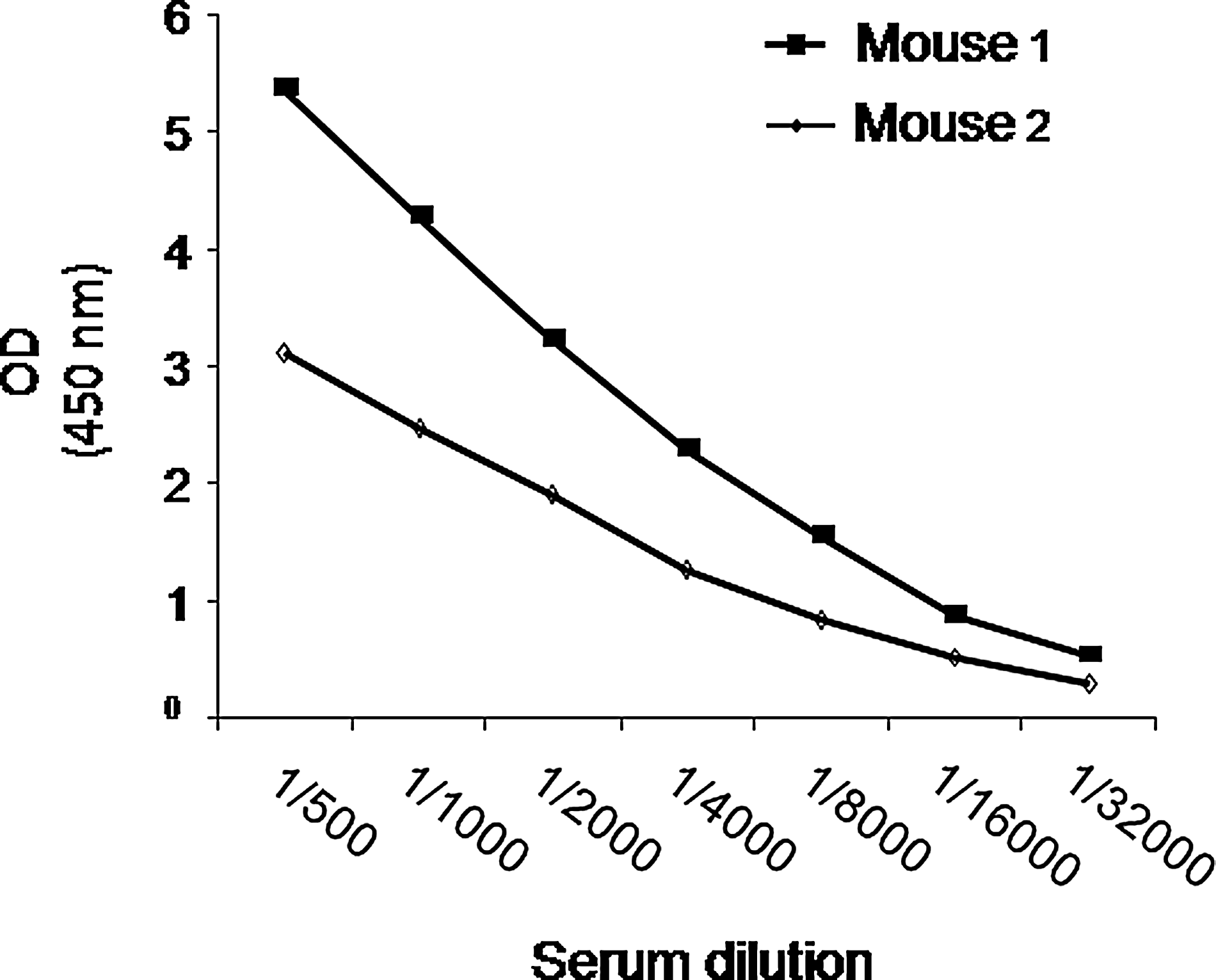
Serum titrations of two immunized mice. Mouse 1 was selected.
ELISA assay
A 96-well plate (Nunc, Roskilde, Denmark) was coated with 50 μL of recombinant FVII (1μg/mL) in PBS and was incubated at 4°C overnight. After incubation, unbound antigens were removed by washing three times with PBS/Tween (PBS-T) buffer for 3 min. Blocking was performed by adding 100 μL of blocking buffer (PBS-T and 2.5% skim milk) in each well. Wells were then washed three times with PBS-T buffer for 3 min. Mouse sera were serially diluted in PBS and then added to each well. After incubation for 1.5 h, the plate was washed, 50 μL HRP-conjugated sheep anti-mouse (Avicenna Research Institute, Tehran, Iran) was added to each well, and the plate was incubated at 37°C for 1 h. After three washings, 50 μL of 3,3′,5,5′-tetramethylbenzidine (TMB) substrate (PishtazTeb, Tehran, Iran) was added and incubated at room temperature for 15 min in the dark. The reaction was stopped by the addition of 15 μL/well stopping solution (20% sulfuric acid). The optical density was measured at 450 nm using a microplate reader (Bio-Tek, Winooski, VT). A mouse with a better antibody response was selected for hybridoma generation.
Hybridoma production
Spleen from the immunized mouse was removed under sterile conditions and cells were isolated and fused to a previously prepared mouse myeloma SP2/0 cell line (ATCC, Manassas, VA) at a ratio of 1:5 using PEG1500 (Sigma-Aldrich, Saint Louis, MO), as described previously.(16) Fusion hybrids were selected using HAT medium and the reactivity of the secreted antibody was tested by ELISA after 5–10 days. Positive hybridomas were then cloned (four times) by limiting dilution until a stable hybridoma clone 1F1-B11 was obtained.
Preparation of ascitic fluid
About 5×106 hybridoma cells from stable clone 1F1-B11 were intraperitoneally inoculated to a BALB/c mouse for producing high-titer monoclonal antibody, as described previously.(17) Briefly, 0.5 mL of Pristane (Sigma-Aldrich) was injected into the peritoneum the mouse 1 week prior to inoculation of cells. Hybridoma cells were washed by resuspension in 50 mL sterile PBS without FBS, centrifuged at 500g for 5 min at room temperature. Cells were counted and adjusted to a concentration of 5×106 cells/mL with PBS prior to intraperitoneal injection. The ascitic fluid was collected after 2 weeks.
Antibody isotype determination
Isostrip mouse monoclonal antibody isotyping kit (Roche, Mannheim, Germany) was used to determine the heavy and light chain isotypes of the monoclonal antibody. Each strip contains IgG1, IgG2a, IgG2b, IgG3, IgM, and IgA as well as kappa and lambda light chains.
Antibody purification
The ascitic fluid was centrifuged; after 1:5 dilution by PBS buffer, it was filtered through 0.45 μm filters (Orange Scientific, Braine-l'Alleud, Belgium). The MAb was purified using a 5 mL Hi-Trap protein G column (GE Healthcare, Uppsala, Sweden). Elution was performed using 0.1 M glycine (pH 2.7). The pH of eluted antibody was adjusted to 7.0 with 1 M Tris buffer (pH 9.0). The eluted antibodies were dialyzed by dialysis tubing (Sigma-Aldrich) against PBS (pH 7.5) overnight, and the reactivity of the antibody was measured by ELISA as described above.
Monoclonal antibody affinity constant determination
To determine the affinity constant (Kaff) of 1F1-B11, a non-competitive enzyme immunoassay was performed, as described previously.(18) Briefly, several concentrations of recombinant factor VII, including [Ag]=0.25, [Ag′]=0.125, and [Ag″]=0.0625 μg/mL were coated in a 96-well plate (Nunc, Roskilde, Denmark) in duplicate. After overnight incubation at 4°C, the antibody was serially diluted in concentrations of 6, 3, 1.5, 0.75, 0.375, 0.187, 0.093, 0.047, 0.023, 0.011, 0.006, 0.003, and 0.0015 μg/mL and added to the plate. The rest of the process was performed as previously described for the ELISA assay. Sigmoid curves were plotted to represent the relationship of OD450 value versus logarithmic MAb concentration in three different antigen concentrations (Fig. 2). The half maximum OD (OD-50) was assigned for all selected curves from which the corresponding antibody concentration ([Ab], [Ab′], and [Ab″]) was extrapolated. Accordingly, [Ab] and [Ab′] are the measurable total antibody concentrations at OD-50 and OD′-50 for plates coated with [Ag] and [Ag′], respectively. The affinity constant was finally determined using the following equation:
Representative binding curves employed for extrapolation of affinity constant of 1F1-B11 MAb.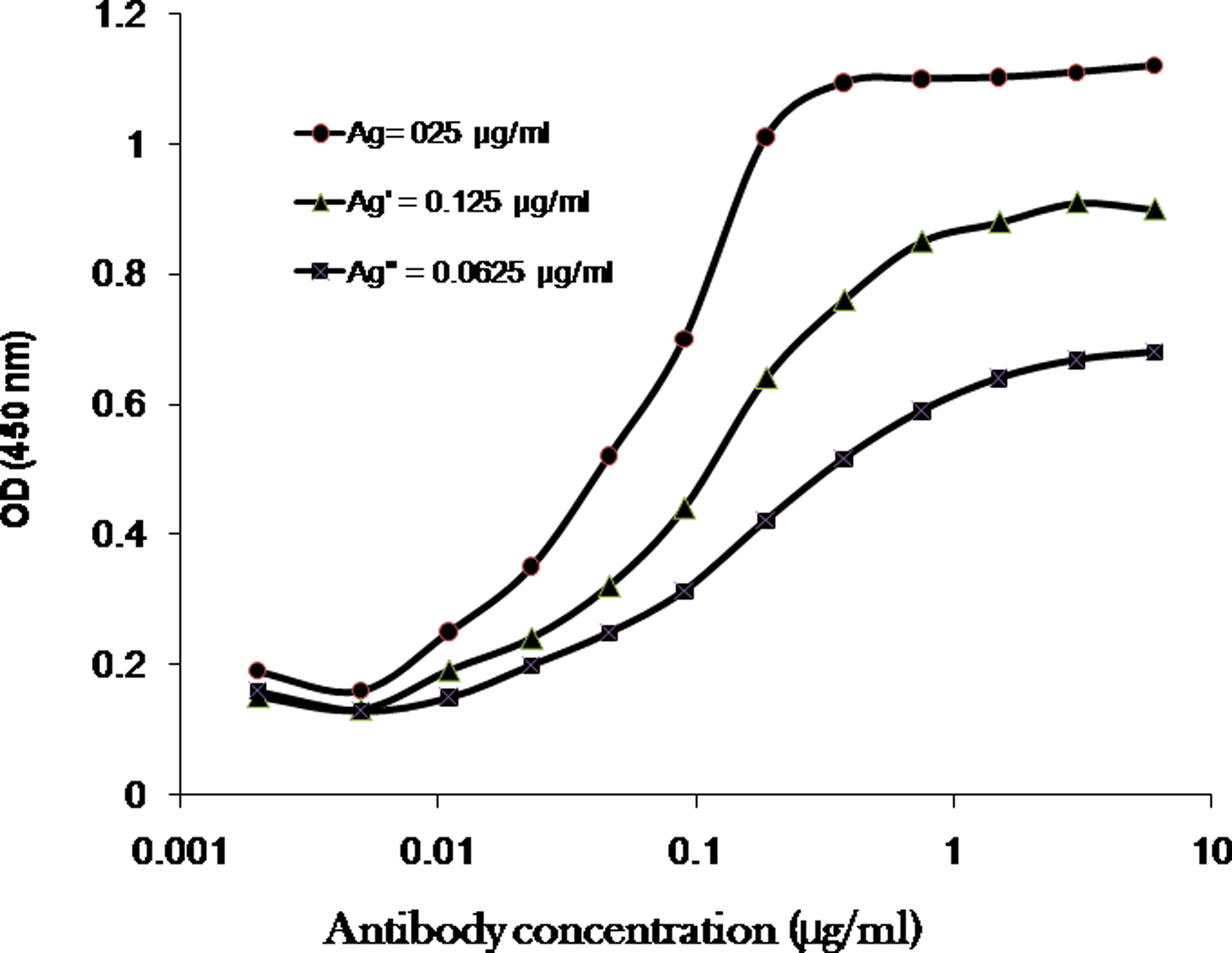
As above, the affinity constant for [Ag′] and [Ag″] was measured and the average of these two Kaff was calculated (Table 1).
Comparison between anti-FVII clone 1F1-B11 and commercial polyclonal antibody
Western blot was performed to obtain the minimum amount of FVII that could be recognized by the 1F1-B11 antibody. To do this, different concentrations of human recombinant FVII (64, 32, 16, and 8 ng) were loaded onto a 12% SDS-PAGE in duplicate to compare 1F1-B11 with a commercial sheep anti FVII polyclonal antibody (Abcam, Cambridge, UK). Electrophoresis was run at 100 volts and 150 mA for 1.5 h. The gel was electroblotted (100 volts, 67 min) onto a PVDF membrane (Millipore, Bedford, MA) and blocked by 5% skim milk at 4°C overnight. Eight μg/mL of 1F1-B11 and polyclonal antibodies were used separately as primary antibody at room temperature while shaking for 1.5 h. PVDF membrane was washed six times for 10 min with washing buffer (0.05% Tween-20, 1×PBS). For 1F1-B11, HRP-conjugated sheep anti-mouse (1:2000) and, for the polyclonal antibody, HRP-rabbit anti-sheep (1:2500) were used as secondary antibody while shaking at room temperature for 1 h. Again, washing was done four times and bands were visualized using advanced ECL (GE Healthcare, Uppsala, Sweden).
Analysis of 1F1-B11 specificity
Fresh citrated blood samples were collected from human and several other species, mouse, rabbit, rat, sheep, bovine, and goat. Samples were rapidly centrifuged and plasma was separated. Diluted plasma (1:50) was prepared and mixed with 0.01 molar benzamidine (Sigma-Aldrich). Equal volumes of samples were loaded on a 12% SDS-PAGE and after blotting to PVDF; 1F1-B11 (8 μg/mL) was used as primary antibody. The Western blot analysis was continued as described above.
Detection of recombinant factor VII in supernatant of FVII-transfected cell line by 1F1-B11
To produce recombinant FVII, the construct containing whole FVII gene was obtained from Origene (OriGene Technologies, Rockville, MD) and transfected into BHK21 cell line using jet-PEI™ reagent (Polyplus, Illkirch, France), according to the manufacturer's instructions. After 48 h, lysate and supernatant were analyzed by Western blot and sandwich ELISA.
Sandwich ELISA for detection of r-FVII by 1F1-B11
ELISA plate wells were coated with 1F1-B11 (3 μg/mL in PBS) and incubated at 4°C overnight. The plate was washed with PBS-T three times and wells were blocked with 2.5% skim milk. After washing, 50 μL of transfected and untransfected BHK21 cell culture supernatants, PBS (as well as recombinant FVII as a positive control) were added and titration was done and incubated at 37°C for 1.5 h. The plate was then washed and 50 μL of HRP-conjugated rabbit anti FVII polyclonal antibody (1:1500; Tehran University of Medical Sciences, Tehran, Iran) were added. Following addition of TMB substrate, the reaction was stopped by adding 20% H2SO4. Color development was assessed at 450 nm in a microplate reader.
Results
Five stable clones—3C7-F11, 1B2-B9, 3A1-B5, 1F1-B11, and 1B11-C5—were generated against human factor VII. 1F1-B11 clone was selected based on the best reactivity at Western blot (Fig. 3). 1F1-B11 is an IgG1-kappa monoclonal antibody that recognizes an epitope in the light chain of human factor VII (Fig. 4).

Western blot analysis of different hybridoma clones using 20 ng of r-FVII on each well. Only 1F1-B11 reacted with FVII.
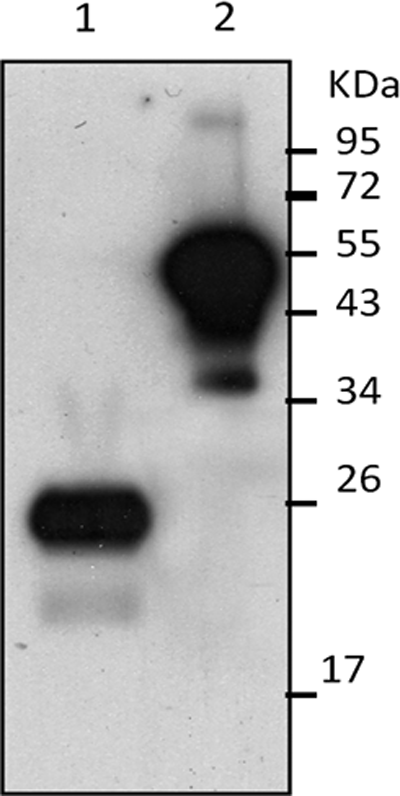
Western blot analysis of 20 ng NovoSeven under reducing (lane 1) and non-reducing (lane 2) conditions with 1F1-B11 (5 μg/mL) showing the recognition of FVII light chain (20 kDa) by 1F1-B11.
According to the Beatty method,(18) the affinity constant of 1F1-B11 monoclonal antibody was calculated to be 6.4×1010 Mol –1 (Table 1).
Sensitivity comparison between 1F1-B11 and commercial polyclonal antibody
Sensitivity of 1F1-B11 MAb was compared to sheep polyclonal anti-factor VII commercial antibody (Abcam) under equal conditions in Western blot analysis. The minimum amount of factor VII recognized by the commercial polyclonal antibody was 32 ng, while 1F1-B11 showed a higher degree of sensitivity, recognizing 8 ng of FVII (Fig. 5).
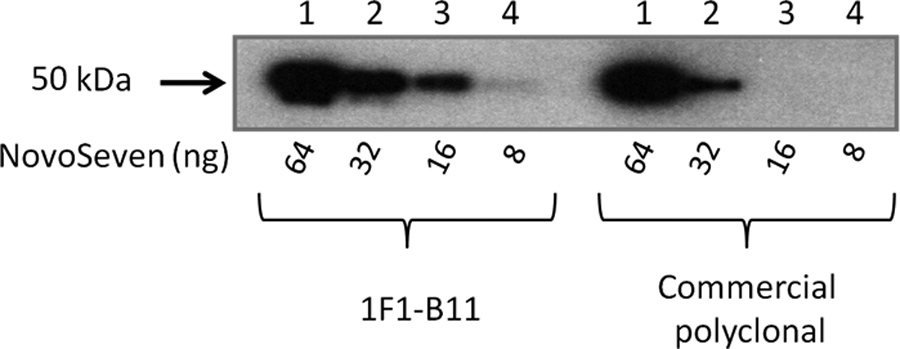
Western blot analysis comparing 1F1-B11 and commercial sheep polyclonal antibody (Abcam) to determine the reactivity to different concentrations of FVII (under non-reducing conditions).
Specificity of 1F1-B11 to human FVII
Western blot assay was done using 1F1-B11 monoclonal antibody on plasma samples of several species, including mouse, rabbit, rat, sheep, bovine, goat, and human. Despite high amino sequence similarity between human factor VII and the FVII of above-mentioned animals, our data revealed that 1F1-B11 exclusively recognized human factor VII, but not factor VII of mouse, rabbit, sheep, bovine, and goat (Fig. 6).

Western blot analysis of plasma (1:50 dilution) from different species. Lane 1, mouse; lane 2, rabbit; lane 3, rat; lane 4, sheep; lane 5, bovine; lane 6, goat; lane 7, r-FVII (30 ng); lane 8, human.
In addition, to test the ability of 1F1-B11 to discriminate between human FVII and other proteins, BHK-21 cell line was transfected with human FVII gene and the lysate and supernatant of the cell culture were tested in Western blot analysis. Interestingly, the antibody exclusively recognized a 50 kDa band in the lysate and supernatant of FVII transfected BHK21 cells, but not in the untransfected cellular counterpart (Fig. 7). This is despite the fact that the culture medium contained bovine FVII from fetal bovine serum.

Western blot analysis of lysates and supernatants from transfected and untransfected BHK-21 cell cultures after 48 h run under non-reducing conditions. Lane 1, commercial r-FVII (20 ng); lanes 2 and 3, concentrated supernatant and lysate (20 μg) of FVII transfected BHK-21 cell culture; lanes 4 and 5, concentrated supernatant and lysate (20 μg) of non-transfected BHK-21, respectively.
The results of Western blot were also confirmed by sandwich ELISA where titrations of commercial factor VII (positive control) and the culture supernatants of FVII-transfected BHK21 cells clearly showed reactivity of the antibody with FVII in contrast to no reactivity with FVII-untransfected BHK-21 cell supernatant (Fig. 8).
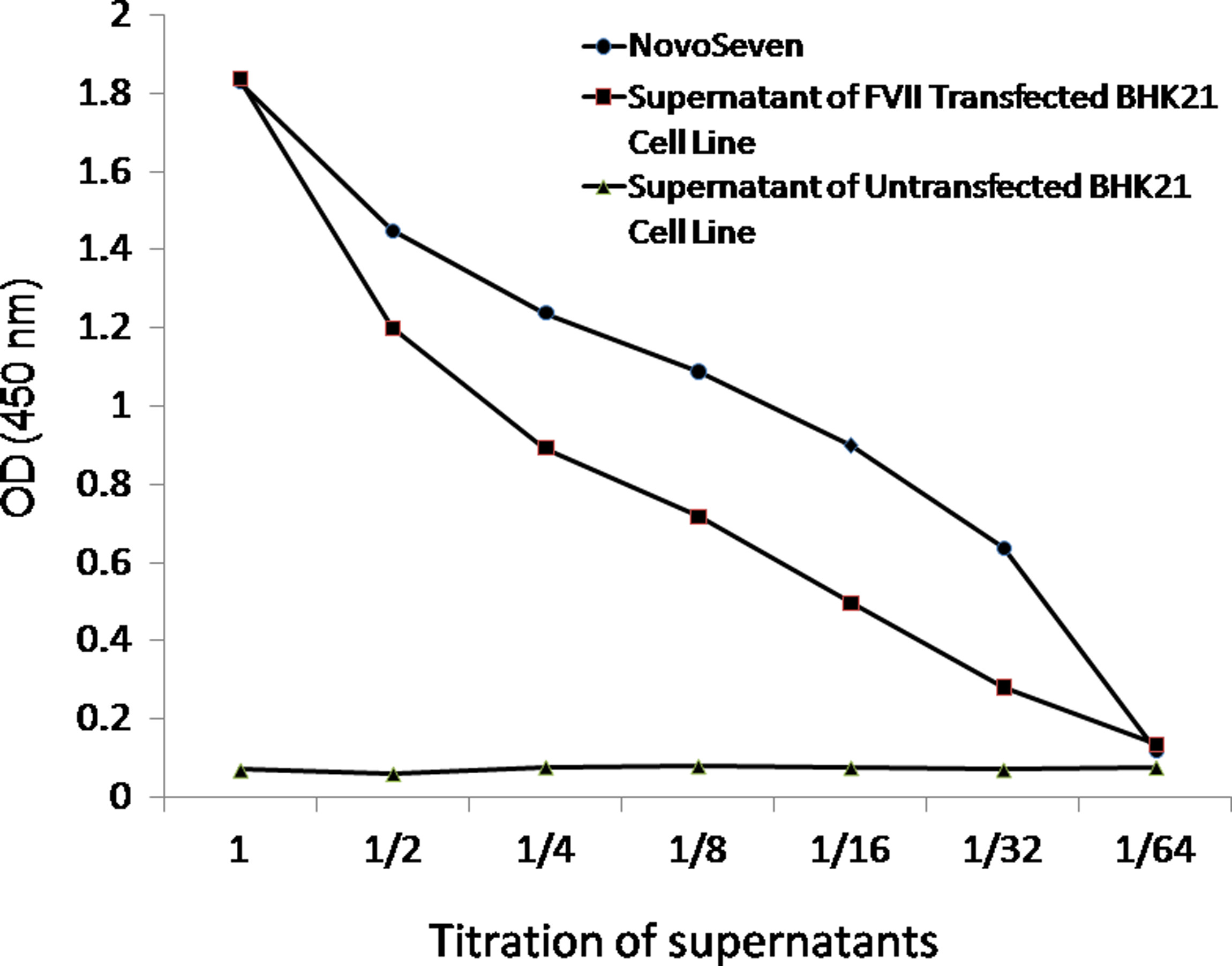
Titration of transfected and untransfected BHK21 cell line supernatants by sandwich ELISA using 1F1-B11.
Discussion
Considering the important role of affinity in the antigen-antibody interaction, an antibody with a higher affinity would be a superior tool in different applications such as immunotherapy,(19,20) tumor targeting,(21,22) immunotoxins,(23,24) immunoaffinity chromatography,(25) and diagnostic assays.(26) In this study we have developed a highly sensitive and specific monoclonal antibody against human factor VII with Kaff=6.4×1010 Mol–1.
The antibody clone 1F1-B11 showed superiority in recognizing minute amounts of human FVII compared to a commercial polyclonal anti-factor VII. In addition, its exclusive recognition of human FVII will ease many experimental problems such as background cross-reactivity with bovine FVII caused by its presence in fetal bovine serum, which is supplemented to most of cell culture media. Based on Western blot data, 1F1-B11 recognizes an epitope in the light chain (20 kDa) of human factor VII under reducing conditions (Fig. 4). Previously reported antibodies against FVII recognize different epitopes in different domains of this protein located on the light chain,(27) heavy chain,(9,28,29) or three-dimensional structures.(29)
Considering the fact that 1F1-B11 did not react with FVII from mouse, rat, rabbit, bovine, sheep, and goat, one may suggest that it recognizes a very restricted linear epitope on the light chain of human FVII. Moreover, with reference to the very high degree of similarity of FVII between human and animals,(30,31) pBlast of only light chains revealed over 70% similarity—mouse (75%), rat (73%), rabbit (79%), and bovine (79%) (sheep and goat FVII amino acid sequences did not exist in protein databases), and the exclusive reactivity of 1F1-B11 with human FVII—it may be predicted that the recognition site of the antibody must consist of a very limited number of amino acids on the light chain, as evidenced by multiple alignment and pBlast in NCBI (data not shown). No significant differences were found by homology searching between human factor VII and factor VII from other species (as mentioned above) to assist in characterization of the recognized epitope by our antibody. In this regard, further experiments on other species need to be done to map the exact antigenic epitope.
Conclusion
The high affinity, specificity, and sensitivity of the 1F1-B11 MAb convert this antibody to a superior tool for purification of recombinant human factor VII by its coupling to sepharose beads. Also, this column can be used for immunodepletion of FVII, producing factor VII deficient plasma from normal human plasma.
Footnotes
Acknowledgment
We extend special thanks to Ms. Shaghayegh Emami and Ms. Raheleh Farahani for technical assistance.
Author Disclosure Statement
The authors have no financial interests to disclose.