
Abstract
Xylan is a principal structural component in plant cell walls and could interfere in many industries and biological activities. Biosynthesis of the xylan-degrading enzyme xylanase by fungal isolates was investigated, using agricultural wastes as substrates for production. Aspergillus fumigatus was shown to be the superior isolate to produce xylanase, and wheat bran proved to be the most appropriate substrate for enzyme production and activity. Media composition and growth conditions were optimized for the production of fungal xylanase using Plackett-Burman multifactorial mixture design and Box-Behnken experimental design. Characterization of the purified xylanase revealed that the most favorable conditions for the maximum enzyme activity (57 IU/mL) were temperature of 55-60°C with thermal stability for 30 min and pH of 9 with enzyme stability for 20 min. Application of xylanase for biodegradation of sugarcane pulp fibers was examined using scanning electron micrographs, which exhibited a vigorous alteration in the topography of fiber surface and structure with a prolonged enzymatic treatment period.
Introduction
Fungal biotechnology is one of the effective means for the conversion of agro-based wastes into certain valuable products. 7 In this context, microorganisms are being used extensively to produce different enzymes, using such materials as substrates. An enzyme such as xylanase, can effectively be used in the processing of various foods. 8
Xylanases are reported to be produced mainly by microorganisms. However, there are reports of xylanases originating from plants, e.g., Japanese pear fruit and European wheat, and from some higher animals, including fresh water mollusk. 9,10 The production of microbial xylanases is generally preferred over plant and animal sources because of their convenience, structural stability, and easy genetic manipulation. 11 Aspergillus spp. have been designated as the most potent organisms for xylanase biosynthesis, including A. niger, A. nidulans, A. kawachii, A. tamarii, and A. fumigatus. 12 –18
The potential value of xylanase for various applications can be achieved through the development of cost-effective enzyme production methods. Fermentation of inexpensive agricultural by-products for xylanase production has been reported by many investigators. 19 The present study was designed for the production, purification, and characterization of fungal xylanase and its application for partial degradation of sugarcane bagasse.
Materials and Methods
Fungal Isolation and Selection
Twenty two soil samples were collected from different agricultural regions in Egypt—Sadat City, Matrouh, and Assiout. From each sample, 25 g was suspended in 250 mL of sterile distilled water, and 0.1 mL from each dilution was spread onto potato dextrose agar media (PDA) adjusted to pH 9.0. The plates were then incubated at 47°C for 5 days. The formed fungal colonies were picked and transferred to fresh PDA plates marked with a stainless steel replicator. Using a replica plating technique, copies from mold colonies grown on master plates were transferred to basal media containing either birch wood xylan (Sigma Chemical Co., St. Louis, MO) or carboxy-methyl cellulose (CMC) as a sole carbon source, and then incubated at 47°C for 5 days (Fig. 1). Colonies grown on xylan and not on CMC media were selected for further examination of their xylanase productivity.
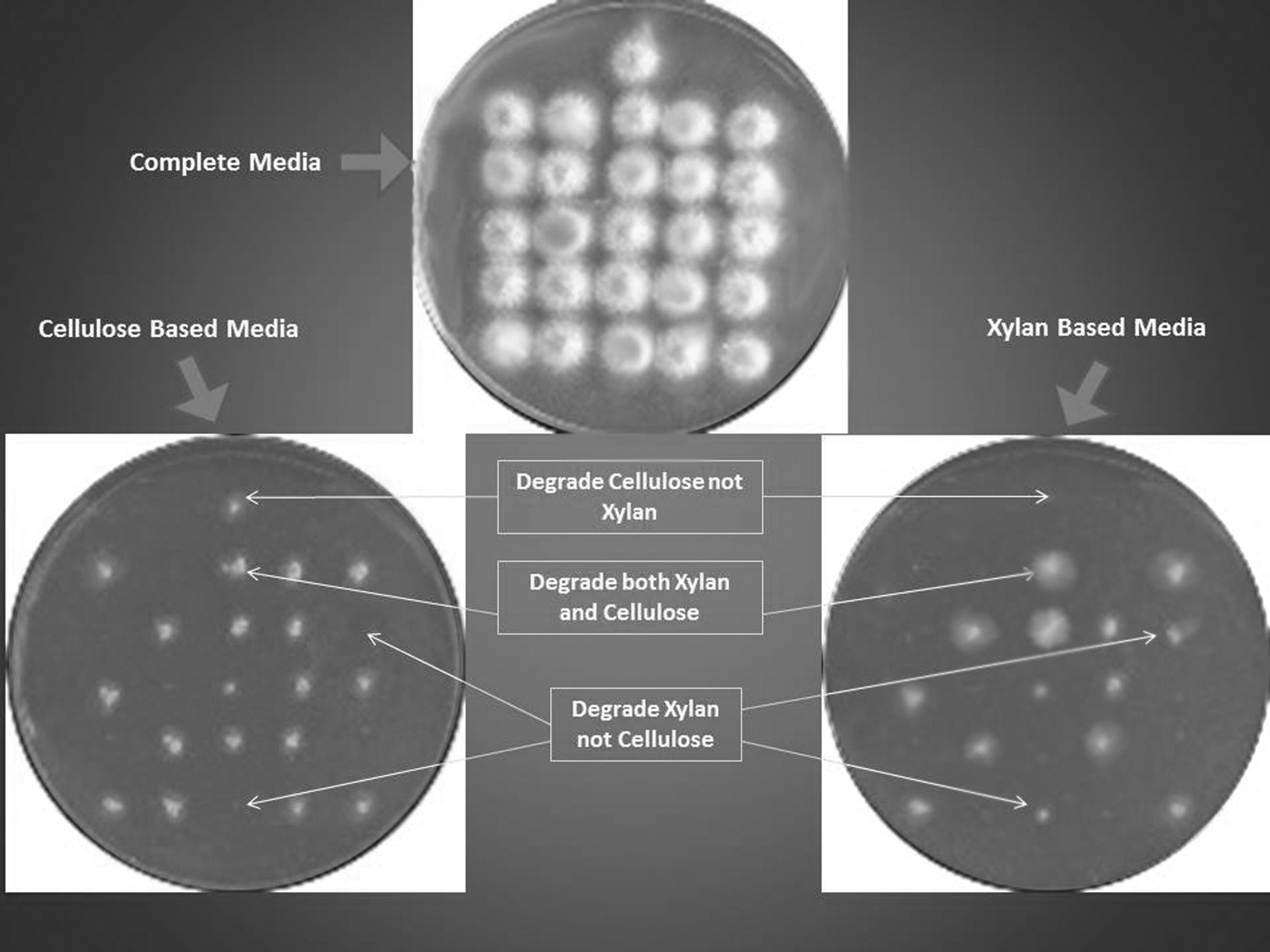
Replica plating technique used for the selection of xylanase, non-cellulase producing isolates.
Preparation of Agricultural Wastes for Enzyme Production
Several indigenous agricultural byproducts and wastes were collected and utilized as substrates to supply production media with carbon sources for the biosynthesis of xylanase enzyme (Table 1). The substrates, collected from local markets, were initially dried with hot air (65°C) in an electric oven until constant weight was obtained. The materials were then ground using a mixing grinder and sieved to obtain approximately 40-mesh size particles. The ground substrates were then used to form production media containing 5% agricultural substrate, 0.1% KH2PO4, and 0.05% MgSO4·7H2O, with pH 9.0. The media were then autoclaved for enzyme production. 20
Plant Substrates with Scientific Names, Cultivar, and Part Used
NCI: No cultivar identified.
Preparation of the Inoculum
Microorganisms were propagated on PDA at 30°C and maintained at 4°C. The conidiospores, grown for 72 h on PDA medium, were harvested in sterile Tween-80 (1%) for use as the inoculum, added to the production media, and incubated at 47°C for 7 days on a rotary shaker at 170 rpm.
Identification of the Microorganism
Identification of the superior fungal strain to produce xylanase was first carried out according to the macromorphology of the strains by observing culture shape and color on PDA plates. Micromorphological characteristics were recognized using a slide culture technique after staining with lacto-phenol cotton blue. Fungal cultures were then grown and maintained on PDA media for further examination.
Enzyme Assay
Because the purpose of the xylanase was to hydrolyze the polymer xylan into xylose monomers, 0.9 mL of 1% birch wood xylan solution in Tris/Boric acid/EDTA buffer (Bio-Rad Laboratories, Hercules, CA), pH 8.5, was treated with 0.1 mL of cell-free broth containing the enzyme for 10 min at 50°C. Xylose was measured using phloroglucinol as a chromogenic reagent; color absorbance was read at 554 nm. 21 Released xylose concentration was calculated using a standard curve of absorbance readings. According to the International Union of Biochemistry, one international unit of xylanase (1 IU) was calculated as the amount of enzyme required to release 1 micromole of reducing sugar (xylose) in 1 minute.
Optimization of the Xylanase Enzyme Produced Through Multifactorial Design of Production Media
Plackett-Burman multifactorial mixture design was applied to evaluate the effect of four nutritional component concentrations (agricultural waste, magnesium sulfate, corn steep liquor, and calcium chloride) and five non-nutritional conditions (pH, incubation temperature, aeration, incubation time, and inoculation level) on fungal xylanase production. 22 Two levels were used for each factor, then parameters having the most influence on enzyme production were examined again using Box-Behnken experimental design to describe the nature of the response surface in the experimental region. 23 Other factors having negative effects were kept constant at their lower levels.
Enzyme Purification
The enzyme was precipitated from cell-free broth using ammonium sulphate (50%) and dialyzed against Tris/Boric acid/EDTA buffer pH 8.5 for 48 h. The precipitate was recovered by centrifugation (15,000 x g for 10 min at 3°C) then applied to a column of Sephadex G-100 (1.8 cm×40 cm) already equilibrated with Tris/Boric acid/EDTA buffer (pH 8.5). Elutions of 10 mL were collected and subjected to xylanase assay procedure and protein estimation measured by monitoring the optical density at 280 nm and at 546 nm. 24
Enzyme Characterization
Optimum temperature for the enzyme was determined by measuring the enzyme activity at different temperatures, ranging from 50-80°C, for 10 min at pH 8.5, according to the method of xylanase assay described previously. The temperature stability was then measured by incubating the pure enzyme at the optimum temperature for 1 h.
The effect of pH on the activity of the purified enzyme was measured at pH values from 7.0 to 11.0 at 55°C for 10 min using phosphate buffer (for pH values 7.0-8.0), Tris/Boric acid/EDTA buffer (for pH value of 8.5) and 0.1 M glycine-NaOH buffer (for pH values 9.0-11.0). pH stability of the purified enzyme was then measured by incubating 0.1 mL of purified xylanase enzyme with 0.2 mL of each buffer at different pH at 55°C, without substrate, for 1 h. The residual activity of the enzyme was then assayed by the standard assay method.
The effects of metal ions on xylanase activity were investigated by incubation with 1 mM Ca2+, Mg2+, Ba2+, Zn2+, Mn2+, Cd2+, Al3+, K+, Cu2+, and Fe2+ in 0.1 M glycine-NaOH buffer, pH 9.0, for 30 min at 55°C. The relative activity was measured at the optimum conditions and compared with that of the control without any metal ions, taking the control as 100% positive.
Application of Xylanase for Bagasse Biodegradation
To determine fungal xylanase's capacity to degrade agricultural lignocellulosic wastes, untreated sugarcane bagasse pulp (Saccharum officinarum) was recurrently washed with tap water to remove the fines and water-soluble compounds. For enzymatic biodegradation, pulp samples (10 g) were pretreated with 57 IU/mL of crude xylanase at 50°C (pH 8.5) for 3 h and 12 h. The reaction was terminated by heating the enzyme-treated pulp samples at 100°C for 20 min. For scanning electron microscope (SEM) examination, xylanase was treated, and control samples mounted on stubs sputter-coated with gold palladium and examined under SEM (JSM-6490LA, JEOL-USA Inc., Peabody, MA) at 10 kV.
Results
The selective isolation of xylan-degrading fungi resulted in a total of 28 morphologically different strains; only five exhibited the ability to grow at 47°C, pH 9, and use xylan and not CMC as the sole carbon source. The most superior strain to produce CMCase-free xylanase was A. fumigatus, which was identified using its macro- and micromorphological features (Fig. 2).

Morphological features of the xylanase producer Aspergillus fumigatus isolate as grown on agar media (
Xylanase Production on Different Substrates
The production and activity of A. fumigatus xylanase were highly influenced by the type of agricultural by-products used as substrate for enzyme production (Fig. 3). Wheat bran proved to be the most appropriate substrate for enzyme production and activity, followed by corn cobs and wheat straw. Alfalfa-based media gave the lowest production and activity of A. fumigatus xylanase, followed by cotton stalk then rice bran-based media.

Activity of A. fumigatus xylanase produced from agricultural by-products.
Plackett-Burman Optimization Design
As wheat bran was the optimal substrate for producing A. fumigatus xylanase enzyme, it was selected to be included in the Plackett-Burman multifactorial mixture design. Regarding xylanase production from formulated media, it could be observed that formula 8 exhibited optimal enzyme activity (Table 2). The evaluation of factors influencing xylanase enzyme activity determined that the concentration of wheat bran had the most influence, followed by pH value.
Influence of Growth Factors on Aspergillus fumigatus Xylanase Activity Using Plackett-Burman Multifactorial Mixture Design
Box-Behnken Factorial Design
To identify the optimum response region for the activity of the xylanase enzyme, the most significant factors—i.e., time of fermentation, substrate concentration, and pH—were further explored at three levels (Table 3). The best formula, giving high activity of the xylanase enzyme (74 IU/mL), was that containing the highest wheat bran concentration (130 g/L) at pH 8 for 11 days.
Effect of Fermentation Time, Substrate Concentration, and pH on Aspergillus fumigatus Xylanase Activity Using Box-Behnken Factorial Design
Purification of Xylanases
Crude filtrates of cultures from A. fumigatus grown on wheat bran exhibited high levels of xylanolytic activity (about 74 IU/mL), but less than (1 IU/mL) of cellulolytic activities (data not shown). The profile of elution from this culture filtrate in Sephadex G-100 chromatography exhibited a peak showing xylanase activity (61 IU/mL) (Fig. 4). A summary of a representative purification protocol is shown in Table 4.
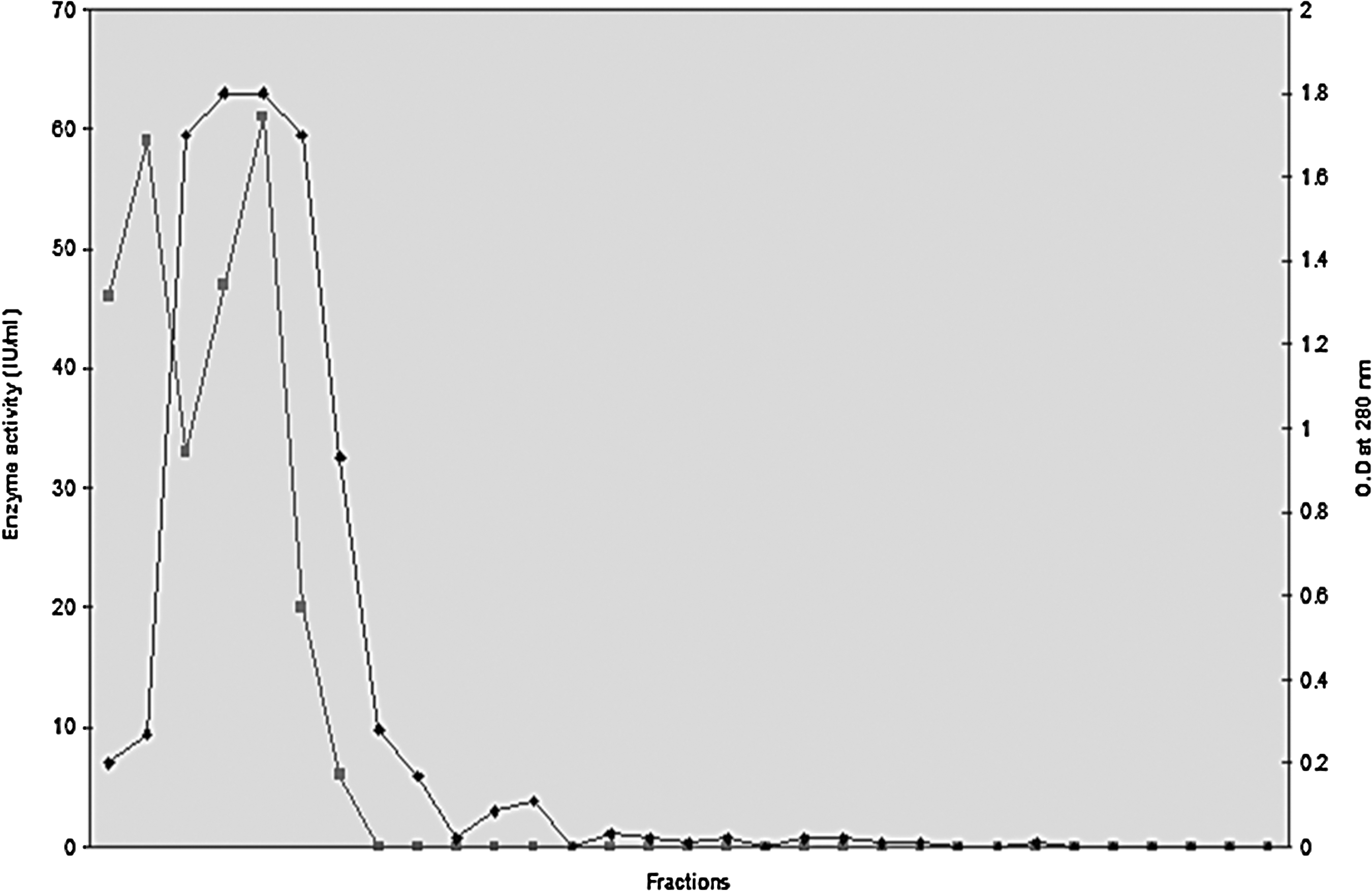
Elution profile of xylanase activity from the culture filtrate and absorbance at 280 nm in a Sephadex G-100 column.
Summary of Purification of Xylanases From Aspergillus fumigatus
Gel Electrophoresis
The purified enzyme showed a single protein band on SDS-PAGE (Fig. 5). The molecular mass of the purified xylanase, estimated from the relative mobility of proteins on SDS-PAGE was ∼22 kDa (second lane).
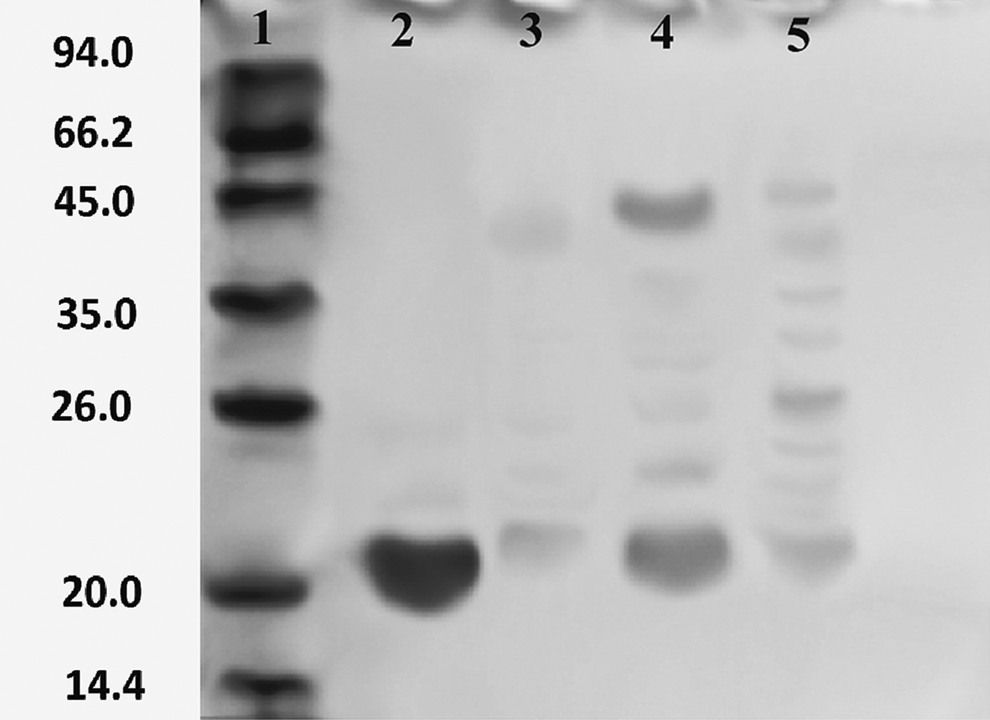
SDS-PAGE analysis of purified xylanases from A. fumigatus. Lane 1: molecular markers; Lane 2, 3: after chromatography column; Lane 4: precipitate of 50% amm. sulphate; Lane 5: precipitate of 70% amm. sulphate.
Effect of Temperature on Xylanase Activity
Figure 6 shows the effect of different temperatures on the xylanase enzyme activity. Data show that the optimum temperature for the enzyme to exhibit its maximum activity was (55-60°C). Further increase in temperature caused a decrease in enzyme activity.
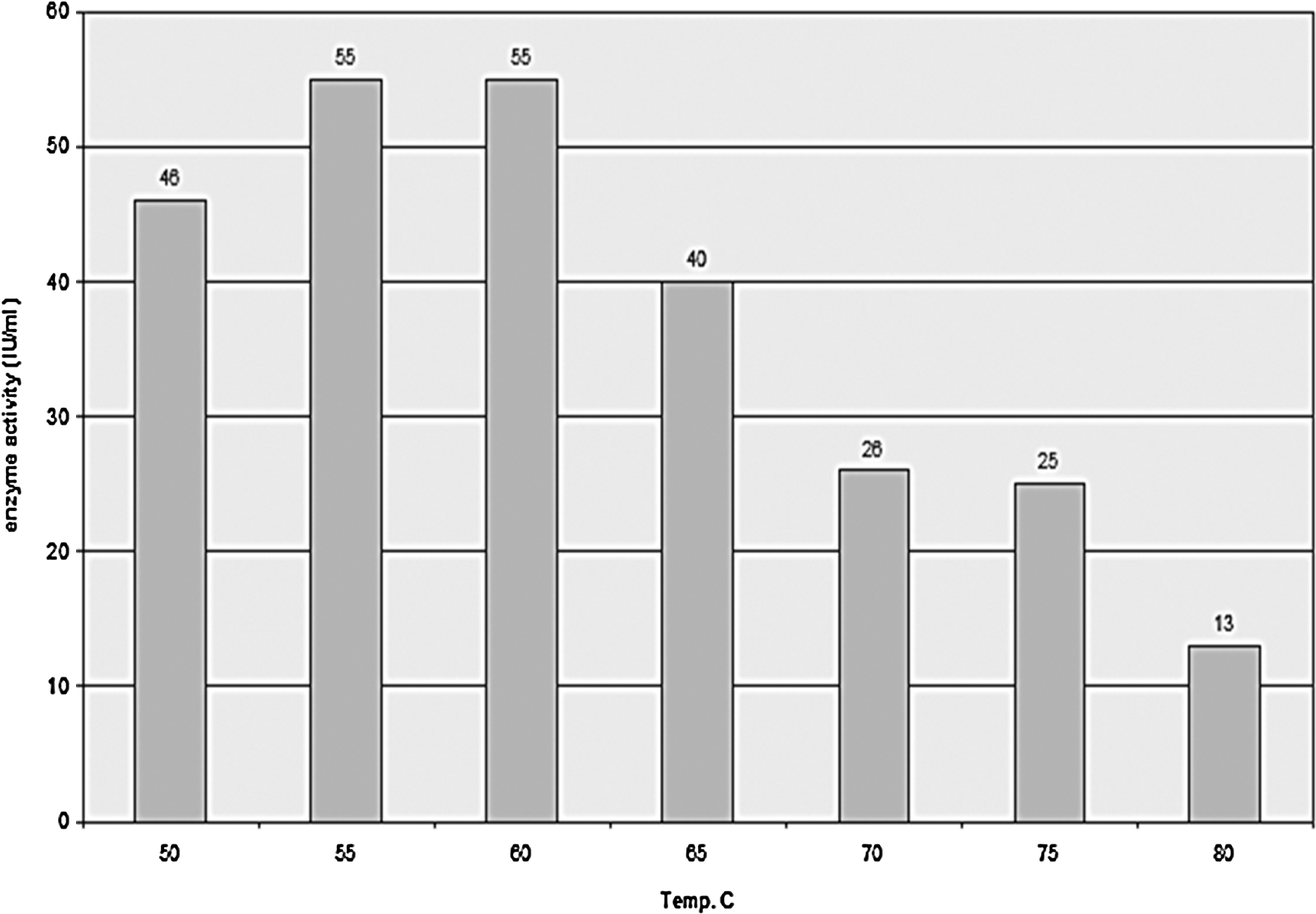
The effect of different incubation temperatures on fungal xylanase activity.
Thermal Stability of Xylanase Enzyme
The enzyme was subjected to an elevated temperature (60°C) for an hour to determine the stability of xylanase. Enzyme performance was maintained for 35 min with only a slight change in its activity. Generally, the increase in time showed a negative effect on enzyme performance (Fig. 7).

Thermal stability of xylanase during 1 h at 60°C.
Effect of p H Value on Xylanase Activity
Enzyme activity under the influence of different pH values (Fig. 8) indicated that the enzyme exhibited maximum relative activity at pH 9.0 (57 IU/mL). The graphical representation shows that further increases in pH resulted in a gradual decrease in xylanase activity.
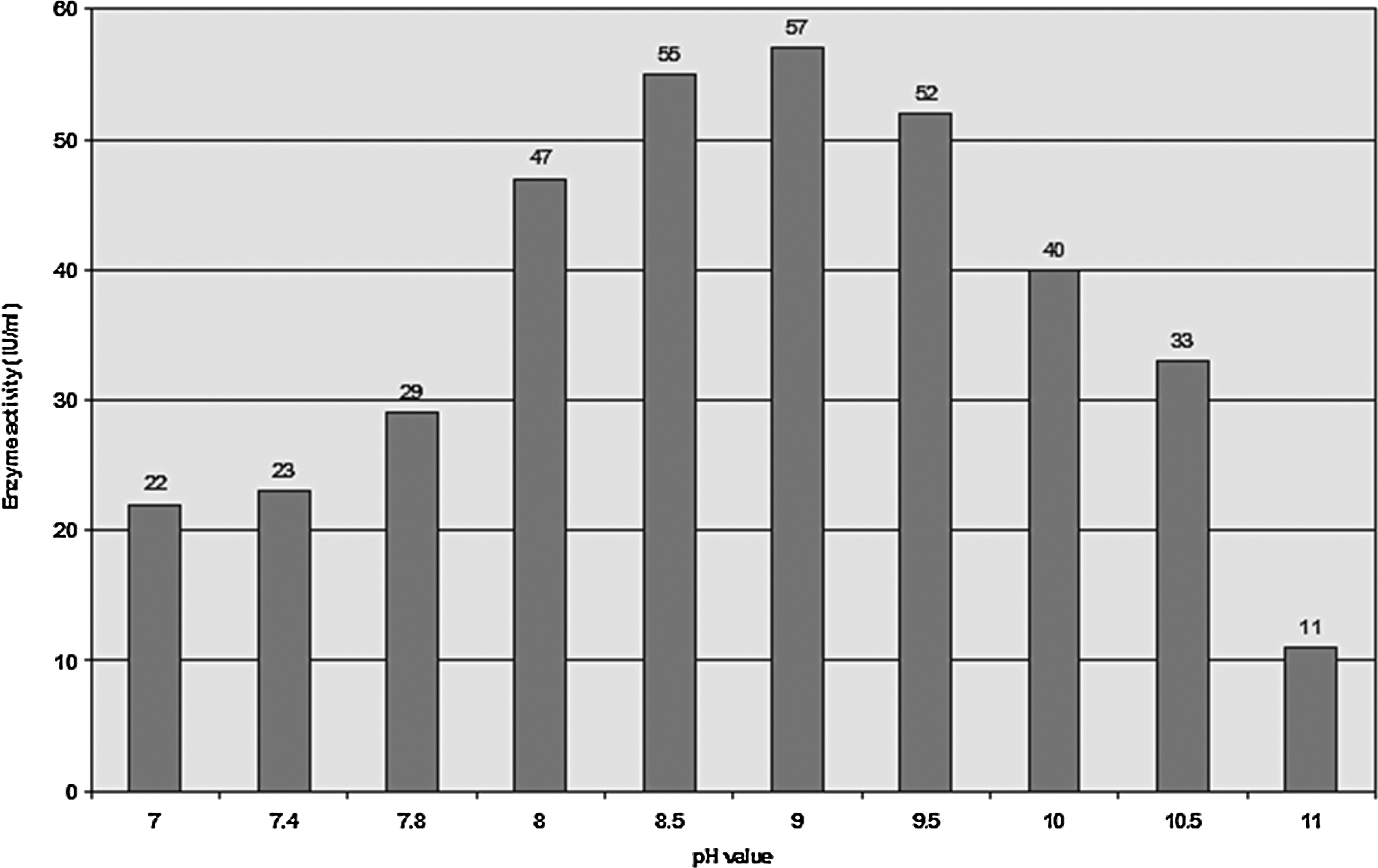
Effect of pH value on xylanase activity.
Effect of p H on Xylanase Stability
Figure 9 shows that the enzyme was almost fully stable at pH 9.0 for 20 min, and then the activity gradually decreased with time.

Effect of pH on xylanase stability.
Effect of Some Metal Ions on Xylanase Activity
The effects of metal ions on xylanase activity are shown in Figure 10. The results demonstrated that xylanase activity was strongly inhibited by the addition of 1 mM Fe2+, Cu2+, and Zn2+ ions, and notably elevated by the addition of 1 mM K+, Mg2+, Al3+ and Ba2+ ions.
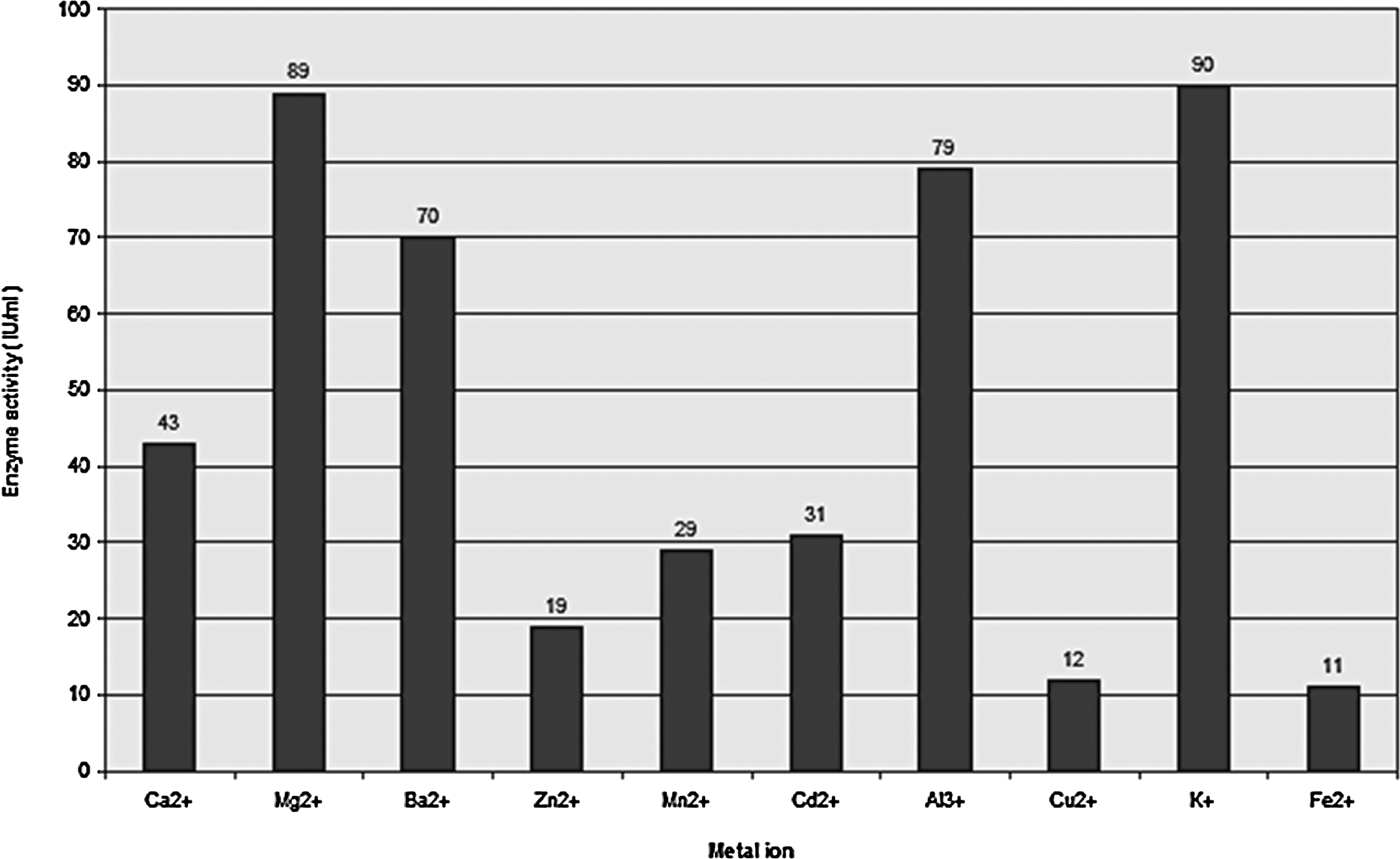
Effect of metal ions on xylanase activity.
Bagasse Treatment with Xylanase
SEM studies clearly showed that the application of crude xylanase caused notable morphological alteration in the bagasse pulp fiber. This indicates that the enzyme was not only involved in xylan hydrolysis but also helped in fiber modification. SEM micrographs indicated that the control bagasse pulp fibers were smooth and showed a uniform surface without protruding fiber formation, whereas bagasse pulp treated with xylanase for 3 h became irregular and heterogeneous with the appearance of swelled and unwrapped fibers on the surface (Fig. 11).

Scanning electron micrographs of treated sugarcane pulp with crude fungal xylanase,
After 12 h of pulp treatment with xylanase, the fibers softened; enlargement and obvious internal degradation could be observed (Fig. 11c). The topography of the fiber surface was vigorously altered with prolonged xylanase treatment. On the other hand, the brightness of treated fibers notably increased with the extension of the treatment period; the fibers' color intensity became lighter, especially after xylanase treatment for 12 h.
Discussion
Egypt is an agricultural country with abundant agro-industrial wastes and by-products. Wheat bran, corn cobs, and rice bran are some of the most widely available waste materials from food industries. If not handled properly, these waste materials can be a source of environmental pollution. This study evaluated xylanase production by Aspergillus fumigatus in submerged fermentation using low-cost substrates as an example of the potential for converting agricultural wastes into value-added products.
There is an increasing push to utilize such neglected materials in the production of enzymes that can be employed further in many applications. 25,26 The inclusion of agricultural wastes and by-products in the production of medium composition for the attainment of high xylanase activity has an extremely important economic significance in xylanase production. 27
Notable differences in xylanase production during fungal growth on examined substrates were observed; this could be attributed either to the combination of other substrate components combined with xylan or to variations in xylan structures. It has been reported that the ratio of cellulose to xylan in the growth substrate is important for production of xylanase. 28 The agricultural substrates used in the current study could be categorized according to their ability to induce xylanase production and activity from A. fumigates: wheat bran, corn cobs, and wheat straw, for example, were strong inducers; banana stalk and rice straw were moderate inducers; and alfalfa, cotton stalk, and rice bran were weak inducers.
The utilization of wheat bran as a substrate for the production of xylanase from many Aspergilli species was recommended by multiple investigators, supporting our results, which found it to be an optimal substrate for production from A. fumigatus. 16,29
The structure of xylans found in plant cell walls can also differ greatly depending on their origin and the structures attached to the xylan backbone. Although most xylans have branched structures, some linear polysaccharides have been isolated. 30 Therefore, the variation in xylanase production from the various lignocellulosic materials used as substrates in the current study could be explained by their dissimilar cellulose contents and/or xylan structures.
It has been suggested that cellulose is protected from cellulases by xylan and mannan. 31 When xylan or mannan is selectively removed from delignified fiber, the residual cellulose becomes accessible to cellulase hydrolysis. However, a similar prehydrolysis of cellulose or mannan does not improve accessibility of xylan to xylanases. On the other hand, xylanases employed in paper technology must not contain any cellulolytic enzymes in order to preserve the cellulose fibers. Selective removal of xylans increases the accessibility of other polysaccharides due to increased fiber porosity that is positively correlated with cellulose hydrolysis in pretreated fibers. 32,33
In the first phase of this study, a Plackett-Burman experimental design was applied to reflect the relative importance of various fermentation factors. The resulting enzyme activity showed wide variation, from 25 to 66 IU/mL, reflecting the importance of studying the medium composition to attain higher productivity. To improve the formula for the subsequent optimization step, the most significant variables—namely wheat bran concentration, pH, and incubation time—were explored at three levels, while the other variables that had negative effects were fixed at their minimum levels.
After the purification steps, the total activity of xylanase enzyme produced by A. fumigatus reached 57 IU/mL, elevated by the application of the optimization steps described above, compared to other reported findings in which the total activity was 10 IU/g or 3.21 IU. 34,35 Another study reported that the maximum xylanase activity at optimum conditions was 0.29 IU/mL. 36 There are many citations in the literature related to xylanase production by fungi using agriculture residues. However, data on Aspergillus are not common, and data specifically on A. fumigatus are rare.
The data derived from this study also showed that the optimum temperature for the enzyme to exhibit its maximum activity was (55-60°C). These results are in line with previous findings, which reported that the purified xylanase from the fungus Aspergillus nidulans KK-99 exhibited highest activity at 55°C. 37 Likewise, optimum activity of xylanase was calculated to be at a temperature of 55°C. 38,39 However, an optimal temperature of 75°C has also been reported. 40 Current results also indicated that the enzyme exhibited maximum relative activity at pH 9.0, and that is in agreement with the findings of Kavita et al., which demonstrated maximum enzyme activity at a pH range 4.0 to 9.5. 37 However, our findings do not agree with other results that reported maximum xylanase activity at pH levels 6.0 to 7.0 39 ; whereas Huang claimed that pH 5.5 and temperature 60°C was the optimum for highest xylanase activities. 41 Goulart et al. cultivated Rhizopus stolonifer on wheat bran to produce cellulase free xylanase, and the purified xylanase exhibited optimum pH and temperature at 6.0 and 45°C, respectively. 42
Our findings regarding the effect of metal ions on enzyme activity are in close agreement with other findings using Aspergillus caespitosus, in which Cu2+, Zn2+, and Ca2+ inhibited the activity of xylanase and Mg2+, Al3+, and Ba2+ greatly enhanced enzyme activity. Using Aspergillus fumigatus FBSPE-05, it was reported that Mn2+ and Zn2+ inhibited enzyme activity, with slight inhibition in the presence of Ca2+ and K+. 43,44
SEM studies showed that the application of crude xylanase caused notable morphological alteration in the bagasse pulp fiber. This clearly indicated that the enzyme was not only involved in xylan hydrolysis but also helped in fiber modification. Results on crude xylanase-aided biodegradation of bagasse pulp showed that the enzyme might have played a role either in releasing the reducing sugars from bagasse or hydrolyzing the xylans precipitated onto the surface of cellulosic fibers. Regarding treated pulp color and brightness, harmonized observations on pulp fiber bioleaching after xylanase treatment have been reported. 45,46
Footnotes
Author Disclosure Statement
No competing financial interests exist.