
Abstract
The specificity of the Simons' staining technique for determining the specific surface area (SSA) of cellulose—the amount of cellulose in a pretreated substrate that is accessible to cellulases—was assessed over a range of cellulosic substrates. Both the Direct Blue (DB) dye and the high molecular weight (HMW) fraction (>100 kda) of the Direct Orange (DO) dye were shown to bind with high specificity to the cellulose rather than hemicellulose and/or lignin. The use of the DO dye by itself could also indicate differences in the accessibility/SSA of various types of cellulose resulting in an R2 value of 0.87 when plotted against enzymatic hydrolysis yields. The previously reported fluctuations in the ratio of DO:DB when the dyes were adsorbed to similar cellulosic substrates were shown to be primarily due to differences in the method used for the isolation of the high molecular weight fraction of the DO dye. A procedure for the enrichment of the >100 kDa HMW fraction of the DO using a minimum of two rounds of ultrafiltration is described. The ability of the modified assays to assess the specific surface area of pretreated and enzymatically hydrolyzed substrates showed good reproducibility.
Introduction
Due to the recalcitrance of biomass toward carbohydrate-degrading enzymes, biochemical processes that hope to hydrolyze the cellulose to sugars for subsequent utilization for biofuels and bioproducts must employ a pretreatment step to increase the accessibility of the cellulose to the carbohydrate degrading enzymes. However, despite a considerable amount of research there continues to be a lack of fundamental understanding of the substrate characteristics of pretreated lignocellulosic substrates that restrict the effective enzymatic hydrolysis of the cellulose. Characteristics such as cellulose crystallinity, degree of polymerization (DP) and specific surface area (SSA)—the amount of cellulose in a pretreated substrate that is accessible to cellulases—are all properties that have been shown to influence the susceptibility of pretreated substrates to enzymatic hydrolysis. 3 The presence of lignin and hemicellulose and the multiple roles these components play in influencing pretreatment, fractionation, and subsequent hydrolysis of the cellulose add another layer of complexity that strongly affects the ease of enzymatic hydrolysis of cellulose.
Variations in pretreatment and fractionation methods, biomass substrate, enzyme types, and loadings and measurement techniques have limited direct comparisons between the many studies that have looked at the predictability of pretreatment processes to enhance the subsequent hydrolysis of the cellulosic component. When groups have tried to look at the influence of a specific substrate characteristic, such as cellulose DP, it is difficult, if not impossible, to change one characteristic in isolation without changing other substrate properties, such as the overall accessibility of the cellulose. To complicate matters further, there is a great deal of contradictory information in the literature, with characteristics such as cellulose DP, crystallinity, hemicellulose and lignin removal, etc., shown to have either an important or negligible influence on the ease of cellulose hydrolysis. 4 –8
In contrast to many of these other substrate characteristics, cellulose accessibility, defined by its SSA and measured by techniques such as Simons' staining, pore volume, and enzyme adsorption, have shown good predictability in estimating the likely effectiveness of enzymatic hydrolysis of pretreated cellulosic substrates. 9 –13 However, due to the complex nature of lignocellulosic substrates, some of the techniques that have been used to estimate the SSA of cellulose have shown some limitations due to the time taken to carry out the assay or for various other reasons. 3 For example, any of the methods that require prior drying of the substrate, such as electron microscopy, gas adsorption, mercury porosimetry, etc., are typically less effective in determining the SSA of the cellulose due to effects such as the irreversible collapse of substrate pores that typically occurs upon substrate drying. 14 Other techniques that can measure the substrate in the wet state are limited, as they only measure a certain fraction of the accessible cellulose in the substrate. 3 For example, techniques such as 1 H NMR thermoporosimetry and solute exclusion only measure the interior surface of the cellulose. Other high-throughput methods, such as the use of a fiber quality analyzer (FQA), can assess the exterior surface area or particle size, but tend to be rather crude in their resolution. 15
As will be reported in more detail below, we have found that the Simons' staining method is relatively fast, simple, and accurate and provides a good estimation of the SSA of a cellulosic substrate and a good indication of how well a substrate might be hydrolysed by a typical cellulase enzyme mixture. 9,16 –22 One of the attractions of the Simons' staining technique lies in its ability to measure the interior and exterior surface area and its sensitivity to small changes in cellulose accessibility. Simons' staining also does not require tedious measurements or an investment in expensive spectroscopic equipment such as NMR.
The Simons' staining method has also been shown to be effective in estimating the interior and exterior SSA of cellulose for a range of pretreated lignocellulosic substrates. The Simons' staining method involves the competitive adsorption between a larger DO 15 dye and a smaller DB 1 dye. The larger DO dye (>100 kDa) has been shown to out-compete the DB dye (998 Da) for adsorption sites in the larger substrate pores due to its higher affinity for hydroxyl groups on cellulose, while the DB dye populates the smaller substrate sites that the large DO dye cannot access. 23 The adsorption and distribution of the DO dye on the cellulosic substrates has been shown to be especially relevant as its molecular size is in the range of 4–7 nm, which is similar to the reported diameter of many cellulase enzymes. 23,24 Previous work has shown that substrates that adsorb a greater amount of the DO dye in comparison to the DB dye are usually more accessible to cellulases. 25,26
In earlier work, we had modified the Simons' staining method, which had originally been used to assess pulp and paper characteristics to see if it could be adapted and refined to give an indication of cellulose accessibility to enzymes after various wood substrates had been pretreated. 9,23 Although subsequent work has successfully utilized the modified Simons' staining method to assess the SSA of the cellulose in pretreated substrates, the specificity of the dyes for cellulose, when compared to lignin and hemicellulose, still needed to be more fully resolved. 9,18 –21,27 –30 In addition, due to the DO dye's approximate molecular weight similarity to cellulases and its high affinity for cellulose, we also wanted to assess if the adsorption of the DO dye alone, in the absence of the DB dye, might provide an even simpler method of determining the SSA of cellulose. However, as the DO dye is not a unique molecule but rather a condensation polymer of 5-nitro-o-toluenesulfonic acid, without a defined molecular weight, one of the main preparative steps in setting up the Simons' stain assay is the isolation of the >100 kDa molecular weight fraction of the DO dye mixture. 31 Although the Simons' stain assay has been successfully used to estimate the SSA of various cellulosic substrates, fluctuations in the absolute values of the ratio of DO:DB adsorption have been observed among earlier studies. 16 –22,30 It was likely that the observed differences in the absolute values for dye adsorption on similar substrates were due to lot-to-lot variations in the DO dye and differences in the procedure used to isolate the >100 kDa DO dye fraction. In the work below we describe a protocol to refine more clearly the preparation of the dyes so that more reproducible and accurate results can be obtained when using the Simons' stain assay method to estimate the SSA of the cellulosic component of a pretreated and enzymatically hydrolyzed lignocellulosic substrate.
Materials and Methods
DO (Pontamine Fast Sky Blue 6BX, lot no.327081) and DO (Pontamine Fast Orange 6RN, lot no. 814071) dyes were obtained from Pylam Products Co. Inc. (Garden City, NY). The cellulase preparation used in this study was Spezyme CP (Genencor International, San Francisco, CA), which had an activity of 83.5 FPU/mL, and a protein content of 119.1 mg/mL. The β-glucosidase preparation used was Novozyme 188 (Novozymes, Franklinton, NC), which had an activity of 350 IU/mL. Avicel PH101 (cat no. 11365, lot no. 1094627), a microcrystalline cellulose, was purchased from Fluka (Buch, Switzerland). Microgranular cellulose (cat no. C6413, lot no. 010M0147), medium cellulose (cat no. C6288, lot no. 020M0190), long cellulose (cat no. C6663, lot no.050M0164), Birch xylan (cat no. XO502, lot no. 010M0169), and beech xylan (cat no. X4252, lot no. BCBC0895) were purchased from Sigma (Madison, WI) and sieved manually through a 180 mesh screen prior to use, as described below. The length-weighted average particle size of each of the celluloses, measured using an FQA, were 0.067, 0.13, and 0.17 mm for the micro-granular, medium, and long celluloses, respectively. Ethanol-organosolv pretreated lodgepole pine (Pinus contorta) was prepared by acid-catalyzed ethanol treatment of wood chips (4×4 cm) in a 2-L rotating digester manufactured by Aurora Products Ltd (Savona, BC, Canada). The cooking conditions were 170°C, 1.1% H2SO4 (w/v) in 65% ethanol (v/v) for 60 min, as described previously. 7 After pretreatment, the solubilized lignin component was precipitated, dried, and sieved through a 180 mesh screen according to Berlin et al. and was designated organosolv soluble lignin (OSL). 32 To obtain a high yield kraft pulp (55% pulp yield), a mixture of pine softwood chips were cooked in a 14% (Na2O) active alkali charge and 15% sulfidity using the same digester as mentioned above at a liquor-to-wood ratio of 4:1 to an H-factor of 750. The kraft pulp, steam, and organsolv substrates were washed extensively and stored at 4°C prior to analysis. Steam-pretreated Douglas-fir (Pseudotsuga menzeisi) and lodgepole pine were prepared at a temperature of 200°C for a residence time of 5 min using 4% SO2, according to Ewanick et al. 33 Steam pretreatment of hybrid poplar at three conditions was carried out the same as for the softwood samples, but using temperatures of 175°C, 185°C, and 195°C for a residence of 5 min using 3% SO2 as an acid catalyst. Protease treated lignin was isolated from the steam-pretreated Douglas-fir substrate according to Nakagame et al. and was designated steam-pretreated lignin (SPL). 34 A portion of the water-insoluble substrates from both the organosolv and steam pretreatments of lodgepole pine were bleached to remove lignin (according to Pulp and Paper Technical Association of Canada standard method G10.U). The fully bleached organosolv substrate (FBOS) substrate was subjected to treatment with 80% phosphoric acid according to Hall et al. to obtain phosphoric acid swollen cellulose (PAS). l5
Chemical Composition of Substrates for Simons' Staining Tests
The chemical compositions of cellulose and lignin samples were tested according to Sluiter et al. 35
Simons' Staining
Unless otherwise specified in the text, the isolation of the 100 kDa fraction of the DO dye and the Simons' staining method was performed according to Chandra et al. 17 All of the filtrations used to isolate the >100 kDa dye started with 10 mg/mL stock solution of the initial unfiltered DO as received from the manufacturer. Filter membranes were changed after each round of filtration. To conserve both dyes and substrates, the procedure was scaled down by a factor of 10 to a total volume of 1.0 mL and performed in 2.0 mL screw cap polypropylene microcentrifuge tubes (Fisher Scientific cat no. 02-681-343). Substrates were weighed (10 mg oven dry basis) into tubes, and the required amount of water and phosphate buffered saline was added. Substrates were allowed to sit overnight in the water/buffer mixture to hydrate prior to the addition of the dye. A coefficient of variance (COV) value was obtained for Simons' stain measurements by using the standard deviation of the DO and DB dye adsorptions to the organosolv pulp, repeated five times. The COVs for the adsorption of the DO and DB dyes were 10.9% and 12.4%, respectively.
Enzymatic Hydrolysis
All of the enzymatic hydrolysis experiments were performed in 50 mL of 50 mM acetate buffer (pH 4.8) at a 2% (w/v) solids loading. The hydrolysis reaction was incubated at 50°C, with shaking at 150 rpm for 72 h. The cellulase loading was 5 FPU/g of substrate cellulose, and the β-glucosidase (Novozym 188) loading was 10 IU per g cellulose (2:1 ratio to cellulase loading). The reducing sugar content was measured by high performance liquid chromatography (HPLC). 7 The hydrolysis yield of the substrate was calculated from the measured reducing sugar content as a percentage of the theoretical reducing sugar available in each substrate.
Results and Discussion
Our findings, and that of other researchers, have found that the Simons' stain assay can be successfully utilized for the rapid estimation of the accessibility of cellulases to the cellulose present in pretreated lignocellulosic substrates. 15 –22,27 –30 While the Simons' stain assay has been shown to be one of the most useful tools for assessing the SSA of cellulose, the relative specificity of the DO and DB dyes for cellulose while in the presence of hemicellulose and lignin has not been fully resolved. Recent studies have also indicated that the DO dye in the absence of the DB dye may be sufficient for estimating the SSA of pretreated lignocellulosic substrates. In addition, due to variations in the absolute values of the DO to DB dye adsorption ratio, further refinements were required to ensure consistency in the procedure used for the isolation of the HMW DO dye.
Significant fluctuations occurred in comparing the DO:DB dye adsorption ratios when the Simons' stain method was used to characterize the same substrates (Table 1). When assessing the various steps involved in the overall Simons' stain assay it appeared that one of the main reasons for the deviations in the DO:DB ratio was differences in the homogeneity of the DO dye. When the DO dye is obtained from the manufacturer it is in the form of a condensation polymer that does not have a defined molecular weight. There are also lot-to-lot variations in the DO dye. It was evident that there were fluctuations in the DO:DB ratios even when using DO dye from the same lot (Table 1). Earlier work had demonstrated the importance of fractionating out the HMW fraction of the DO dye, and since the DB dye had a defined molecular weight, the observed differences in the DO:DB ratio were likely due to inconsistencies in the ultrafiltration step used to isolate the >100 kDa dye fraction from the DO dye mixture (Table 1). 23
Variations in the Ratio of Adsorption of DO to DB on Organosolv- and Steam-Pretreated Lignocellulosic Substrates
Dye lot number 1 was a sample from Pylam products from January 2007 (lot number 2 was 814071).
Corresponds to reference numbers.
As mentioned earlier, the lower molecular weight (LMW) fractions of the DO dye have a lower affinity for cellulose and, unlike the HMW fraction, compete poorly with the DB dye for binding sites on cellulose. 9 Therefore, if a substantial amount of the LMW fraction of the DO dye is present, the ratio of DO:DB dye absorption will decrease, since a greater amount of the DB dye will successfully compete with the LMW DO fraction and preferentially absorb to the substrate. 23 As the HMW fraction of the DO dye can constitute as little as 20% of the overall dye composition, careful purification of the HMW fraction from the dye mixture is essential. 23 In earlier work by Yu and Atalla, this intermediate molecular weight (IMW) fraction was also referred to as HMW DO dye, while the LMW DO dye fractions had molecular weights closer to that of the DB dye. 23 Although the DB dye can normally out-compete the smaller molecular weight fraction of the DO dye, the adsorption of the IMW (just below <100 kDa) fraction of the poorly filtered DO dye will increase the relative amount of the DO dye adsorbed, because the IMW fraction can access substrate pores/cracks that are normally inaccessible to the HMW DO dye (>100 kDa).
To resolve this possible interference from the lower and intermediate weight DO dye fractions, the DO dye was ultra-filtered multiple times and, after each round of filtration, the relative adsorption of the DO and DB dyes were assessed on a fully bleached organosolv pulp. This pulp was utilized because, unlike other model substrates such as Avicel, the organosolv-pretreated substrate retains the biomass fiber structure, which should be more representative of pretreated biomass. 15 In addition, the lignin and hemicellulose removed during organosolv pretreatment helps facilitate subsequent further delignification and provides a fibrous substrate with cellulose content greater than 90%.
The greater total amount of the unfiltered DO dye that was adsorbed to the pulp indicated the much wider range of molecular weights that were present in this mixture, with this wider range allowing more of the DO dye to adsorb to the substrate (Fig. 1). It was also apparent that a greater amount of the DB dye was also adsorbed onto the pulp as, due to the relatively high amount of the lower molecular weight DO fraction in the DO mixture, the smaller DO dye fractions in the unfiltered DO dye could not out-compete the blue dye. As the higher molecular weight DO fraction was increasingly purified after sequential filtration, the relative amount of the DO dye adsorbed to the substrate gradually increased because of its higher proportion in the mixture and its greater affinity for cellulose compared to the lower molecular weight components present in the crude DO dye mixture (Fig. 1). Consequently, the amount of DB adsorption decreased due to the higher proportion of >100 kDa DO molecules that were able to out-compete the DB dye.
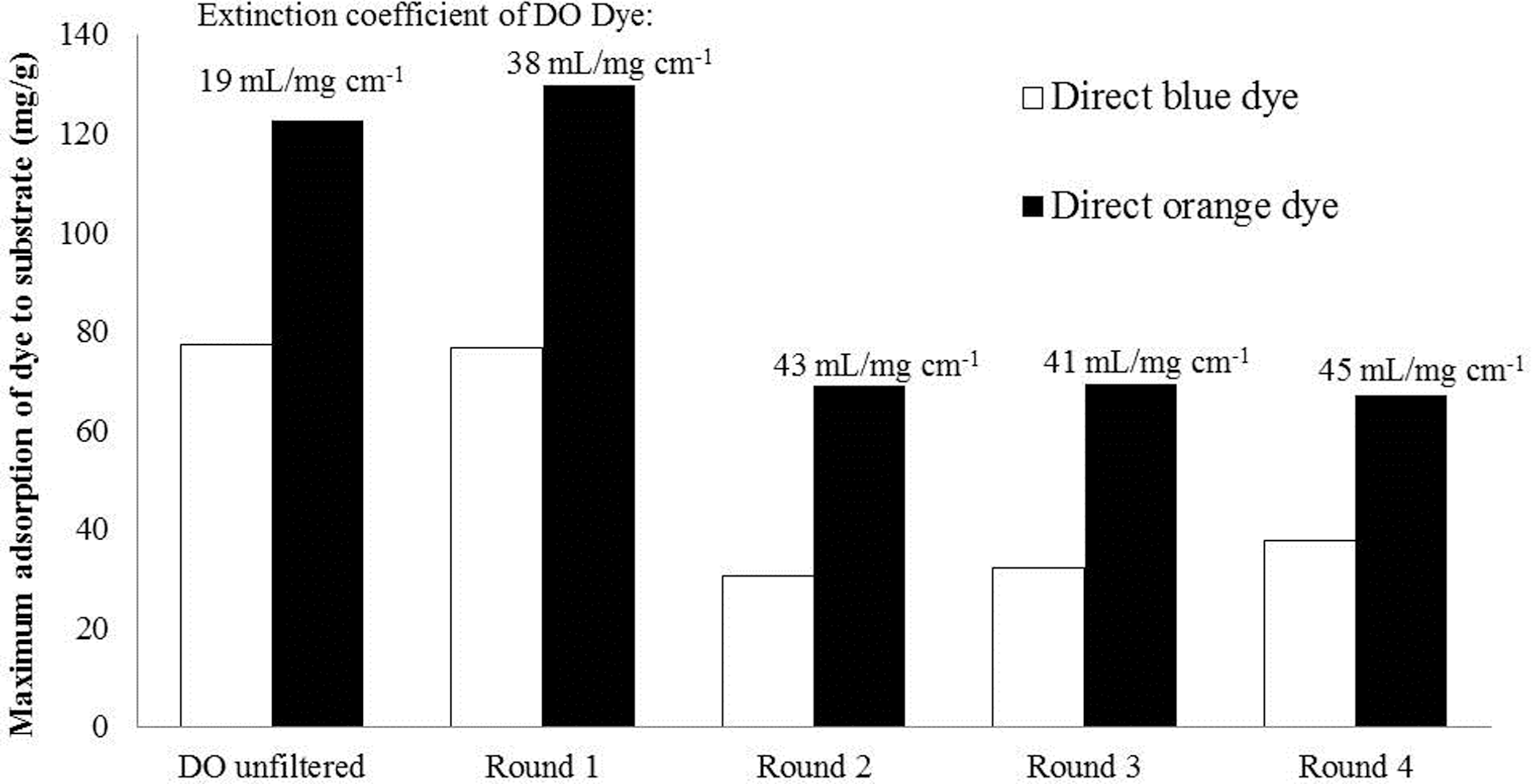
Influence of sequential rounds of ultrafiltration of DO dye (to isolate the >100 kDa molecular weight fraction for use in the Simons' staining test) on the adsorption to a fully bleached organosolv pulp with a consequent increase in the extinction coefficient of the resulting dye mixture. The COV for the adsorption of the DO and DB dyes were 10.9% and 12.4%, respectively, as determined by adsorption of the dyes to the fully bleached organosolv pulp in five replicates.
In addition to interfering with the ratio of DO and DB dye absorption it was also possible that the lower and IMW dye fractions in the unfiltered DO dye lowered the overall extinction coefficient of the DO dye mixture. To assess this possible effect, the LMW 10−30 kDa and intermediate IMW 30−100kDa fractions were isolated and shown to have extinction coefficients of 21.6 and 39.7 mL/mg cm−1 respectively. When the crude DO dye mixture was sequentially filtered, although the extinction coefficient doubled after the first round of ultrafiltration, (resulting from the preferential removal of the DO dye fraction with a MW of less than 30 kDa), the DO dye mixture behaved quite similarly to the unfiltered dye (Fig. 1). Staining the pulp with the IMW fraction of the DO dye indicated that slightly more of this IMW DO dye fraction was adsorbed to the pulp (75 mg/g). As mentioned earlier, this IMW range of dye likely populates the substrate pores that were inaccessible to the HMW DO dye, consequently increasing the overall absorption of the DO dye to the pulp after the first round of filtration. Since the intermediate fraction has a lower molecular weight than the HMW DO dye fraction, its adsorption to the substrate is likely not indicative of the cellulase enzyme accessibility to the pulp. After the second round of filtration, which predominantly removed the IMW DO dye fraction, the DB dye adsorption decreased due to the increased proportion of the >100 kDa DO dye fraction (Fig. 1). After four successive rounds of ultrafiltration, the ratio of adsorption of DO:DB dyes to the pulp increased from 1.5 to a maximum of 2.3. Thus, to obtain good reproducibility and to minimize interference, the crude unfiltered DO dye should always be ultrafiltered for a minimum of two rounds of filtration prior to its use in the Simons' stain assay.
It is likely that in some of the earlier work using the Simons' stain assay, the DO dye contained significant amounts of the LMW DO dye fraction, thus reducing the DO:DB ratio. It is also likely that in this earlier work most of the available cellulose was stained by the range of molecular weights in the crude DO dye and the DB dye, potentially indicating the total cellulose content. 18 Although it might be anticipated that a lignocellulosic substrate enriched in cellulose would contain lower amounts of hemicellulose and lignin, and thus would be more accessible to the DO dye, it has been shown that swollen cellulosic substrates (such as phosphoric acid treated cellulose) can have a similar cellulose content to substrates such as Avicel, but vary significantly in their cellulose accessibility or the SSA of the celluose. 5
As the HMW fraction of the DO dye is about the same size as many cellulases, we next assessed the ability of the DO dye in the absence of the DB dye, and vice versa, to estimate the SSA of various cellulose samples that contained a similar cellulose content but likely differed in their accessibility. 23,24
It was apparent that the DO dye was able to resolve differences in the SSA of the different cellulosic samples with an R 2 value of 0.87 (Fig. 2; Table 2). When the >100 kDa fraction was enriched and purified, the adsorption of the DO dye was shown to be independent of the cellulose content of the substrate (Fig. 2). It was also apparent that the DB dye was far less effective in detecting changes in the SSA of the cellulosic substrates, as plotting the enzymatic hydrolysis yields vs. the DB adsorption gave an R2 value of 0.60. For example, the long, medium, microgranular, and Avicel cellulosic samples adsorbed similar amounts of the DB dye, while their enzymatic hydrolysis yields varied from 19–31% (Table 2). In contrast, for the same samples, the DO adsorption varied from 15–52 mg/g. Although the DO dye was able to provide a good estimate of the SSA of these cellulose-rich substrates, the possibility that the dye might also adsorb to the lignin and hemicellulose fractions in a pretreated substrate still needed to be resolved.

The adsorption of the enriched (>100 kDa) DO dye to various cellulose samples compared to their ease of enzymatic hydrolysis. Cellulose hydrolysis experiments performed at a 2% substrate consistency using a cellulase loading of 5 FPU/g substrate cellulose and a β-glucosidase loading of 10 IU/g substrate cellulose. The COV for the adsorption of the DO and DB dyes were 10.9% and 12.4%, respectively, as determined by adsorption of the dyes to the fully bleached organosolv pulp in five replicates.
Chemical Composition and Adsorbance of DB and DO Dyes Separately to Celluloses, Lignin, and Xylan
SP: steam-pretreated; EO: ethanol organosolv; PAS: phosphoric acid swollen; Bdl: below detection limit.
The individual adsorptions of DO (>100 kDa) and DB dyes to each sample.
According to manufacturer.
Both lignin and hemicellulose are known to limit enzyme access to the cellulose component of pretreated substrates. 36 –38 It is inevitable that some residual lignin and hemicellulose will be associated with the cellulosic component of pretreated and fractionated biomass; even pulping methods that have a goal of producing very pure cellulose pulps still contain some lignin and hemicellulose. 39 Both the lignin and xylan samples adsorbed a small amount of the DO and DB dyes when compared to the total adsorption of the dyes to the pulp sample (Table 2). For example, 70 mg/g of the DB dye adsorbed to the pulp, as compared to only 2.1 mg/g on the corresponding OSL that was isolated from the organosolv pulp. In general, a greater amount of the DO dye was adsorbed to the lignin and xylan samples, compared to the DB dye, which was likely indicative of the higher affinity of the HMW DO dye for the hydroxyl groups present in these samples.
Although a greater mass of the DO dye was adsorbed to the lignin and hemicellulose substrates, the molecular weight of the DO dye is at least 100 times greater than that of the DB dye. Therefore, the absorption of far fewer DO dye molecules to the lignin and hemicellulose samples compared to the DB dye molecules should be measured as a higher total mass of DO dye. Direct dyes such as the DO dye are large planar molecules that become fixed to cellulose via hydrogen bonding between the dye's polar groups and cellulose's hydroxyl groups. 40,41 It was apparent that the OSL adsorbed a greater amount of the DO dye when compared to SPL (Table 2). Since the OSL was isolated from the organosolv liquor fraction, it is likely that it had undergone a greater amount of fragmentation of β-O-4 and α-O-4 linkages during the pretreatment reaction that facilitated the solubilization of the lignin into the ethanol solvent. Consequently, lignin fragmentation will result in an increase in the number of phenolic groups that could mediate the hydrogen bonding of the DO dye to the OSL. 41,42 The adsorption of the DO dye to the OSL was also in the same range as that observed in the case of Brilliant Red HE-3B dye adsorption to industrial lignin (Table 2). 43 This is in line with previous work that demonstrated that anionic direct dyes such as the DO and DB dyes have a greater affinity for cellulose, while cationic and cationic phthalocyanine dyes had a greater affinity for the lignin and hemicellulose components, respectively. 44
Although the lignin and hemicellulose samples were quite pure (>90%), the adsorption of both the DO and DB dyes to these samples was low when compared to the highly accessible, cellulose-rich, organosolv pulp. However, due to its limited accessibility, the long cellulose sample, which was composed of 92% cellulose, adsorbed only 15 mg/g of the DO dye—in the same range as that observed for the lignin and hemicellulose samples. In contrast to the DO dye, and due to its smaller size, 62 mg/g of the DB dye was adsorbed to the long cellulose sample, compared to 2–5 mg/g adsorption to each of the lignin and hemicellulose samples.
To try and gain further insights into the possible effects of the lignin and hemicellulose components on the adsorption of the DO dye, the adsorption of the DO dye on a range of pretreated substrates varying in their lignin and hemicellulose contents was also assessed (Table 3). Softwood (lodgepole pine) pulp chips were steam- and organosolv-pretreated to generate substrates with lignin contents of 43% and 18%, respectively, while each substrate contained less than 3% hemicellulose. Subsequent enzymatic hydrolysis, at a loading of 5 FPU/g cellulose, resulted in only 20% of the original cellulose in the steam-pretreated softwood substrate being hydrolyzed after 72 hours, compared to 66% in the case of the organosolv-pretreated substrate, likely due to the higher lignin content of the steam-pretreated substrate. The high lignin content of steam-pretreated softwood has been shown to limit the accessibility of the cellulose to cellulases. 20 Adsorption of the dye paralleled this recalcitrance as the steam-pretreated softwood adsorbed only 22 mg/g of the DO dye compared to 60 mg/g DO dye in the case of the organosolv-pretreated softwood (Table 3). To determine the potential influence of lignin removal on the ease of hydrolysis and adsorption of the DO dye to the steam and organosolv-pretreated substrates, we next delignified each of the substrates using sodium chlorite. Similar hydrolysis yields were obtained with the delignified steam and organosolv pulp substrates, and this was reflected in the similar amounts of DO dye adsorption by each substrate.
Adsorption of DO Dye to Pretreated Substrates
Bdl: below detection limit.
Cellulose hydrolysis experiments performed for 72 h at a 2% solids loading using a cellulase loading of 5 FPU/g substrate cellulose and a β-glucosidase loading of 10 IU/gram substrate cellulose.
Cellulose hydrolysis performed as described above, but reported yield after 24 h.
Similar to the approach taken with varying the lignin content of the softwood biomass, an abbreviated alkaline kraft pulping cooking condition (H-factor: 750) was also applied to the lodgepole pine chips to provide a substrate with comparable lignin content to the organosolv substrate (15%) but with a higher hemicellulose content (10%). Yields of 40% and 66% were obtained after subsequent hydrolysis of the kraft and organosolv softwood pulps, respectively, at an enzyme loading of 5 FPU/g. This suggested that the higher hemicellulose content may have inhibited the enzymatic hydrolysis of the substrate and, consequently, the adsorption of the DO dye (Table 3). As has been shown with lignin, hemicelluloses such as xylan have also been shown to limit the accessibility to cellulases during enzymatic hydrolysis, thus decreasing hydrolysis yields. 36 –38 Delignification of the kraft softwood pulp further enriched the hemicellulose content to nearly 15%, which limited the cellulose accessibility to both the DO dye and the cellulases compared to the fully bleached organosolv softwood substrate, which contained <3% hemicellulose (Tables 2,3). It was evident that the DO dye alone was capable of estimating the accessible cellulose of the softwood substrates, even when relatively high levels of either lignin or hemicellulose were present.
Previous work had shown that the combined DO and DB dyes were able to provide an estimate of the SSA of the cellulose present in steam- and organosolv-pretreated softwoods. 16 These pretreated softwoods substrates contained low levels of hemicellulose and differed primarily in their lignin content. 16 Therefore, to assess the use of the DO dye alone for selective estimation of cellulose accessibility in substrates that contained hemicellulose and lignin, steam-pretreated poplar samples were treated over a range of temperatures (175–195°C) and assessed for cellulose accessibility and ease of enzymatic hydrolysis. It was apparent that, by raising the temperature and thus the severity of the steam pretreatment, a corresponding increase in both the accessibility of the substrate to the DO dye and its enzymatic hydrolysis could be obtained (Table 3). 45 It should be noted that the presence of >30% lignin and 2–7% hemicellulose within the steam-pretreated poplar substrates did not adversely affect the ability of the DO dye to act as a probe for the detection of differences in cellulose accessibility as the temperature of the pretreatment was raised.
Conclusions
The Simons' stain assay is an effective method for estimating the accessibility of the cellulose present in pretreated and enzymatically hydrolyzed lignocellulosic substrates. However, it is crucial that the HMW fraction of the dye (>100 kDa) be enriched and purified as much as possible to improve the resolution of the assay. As a result, the DO dye can compete with the lower molecular weight DB dye when both dyes are used simultaneously to measure accessibility. The efficacy of the Simons' stain assay can be primarily attributed to the high specificity of adsorption of the HMW DO dye fraction to the cellulose and the similar average molecular size of the dye and cellulase enzymes, resulting in similar accessibility challenges. The work reported here supports the suggestion that accessibility of the enzymes to the substrate is a primary limitation in achieving effective enzymatic hydrolysis of pretreated substrates. Although the refined Simons' stain method cannot directly quantify the specific surface area of cellulose, it can provide a rapid, reproducible estimation of cellulose accessibility and give an indication of a substrate's probable ease of enzymatic hydrolysis.
Footnotes
Acknowledgments
The authors would like to thank the National Science and Engineering Research Council (NSERC), The NSERC Bioconversion Network, and Genome British Columbia for their financial support of this work. They also thank Genencor and Novozymes for their donation of enzymes. We also want to thank our colleagues, Pablo Chung, Jenya Dudley, Carolina Chanis, Kathy Au-Yeung, Linoj Kumar, and Dr. Teresita Marzialetti for their assistance with this work.
Author Disclosure Statement
No competing financial interests exist.