
Abstract

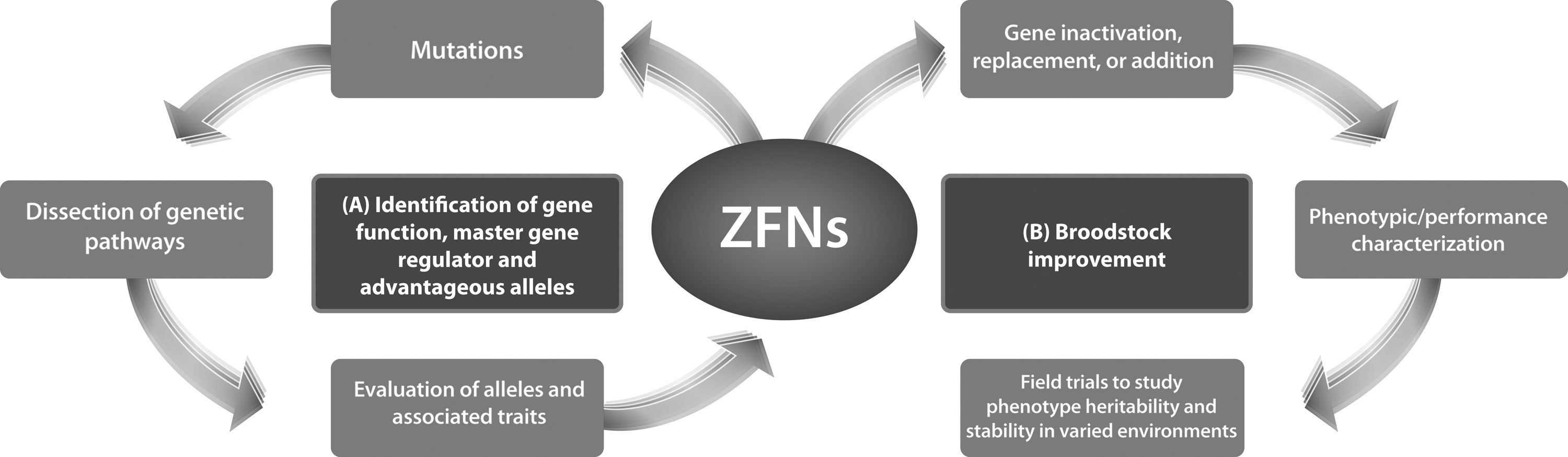
ZFN-induced genetic engineering can selectively modify the genome of commercial finfish, which until recently could only be altered using conventional breeding techniques. The technology enables better dissection of the genetic pathways responsible for accelerated growth rates, nutrient digestion and metabolism, reproduction, immune responses and host pathogen interactions (A) and will help build the foundation for rational design of genetic optimization that will help achieve sustainability in aquaculture (B).
ZFN-Tailored Targeted Genome Editing
ZFNs are engineered proteins that enable genomic editing exclusively at a desired locus, in any species and practically any cell type in which ZFN-encoding DNA or mRNA can be efficiently delivered. A variety of genomic edits can be introduced: gene knockout (elimination of a protein by disrupting its encoding gene, including genes with multiple alleles); gene modification (introduction or correction of a single nucleotide polymorphism, SNP); targeted gene integration (insertion of a transgene at a user-specified site without a transgene landing pad). Targeted integration eliminates the risk of insertional mutagenesis or phenotypic heterogeneity due to position-effect variation. The high efficiency of ZFNs enables trait-stacking of multiple targeted modifications in the same cell line.
ZFN-mediated genome editing requires a single transfection experiment. The ZFNs are then transiently expressed, and editing typically occurs within three days. Experimental data have demonstrated the desired genomic edit often occurs in up to 20% of the cell population, which eliminates the need for selection markers. ZFN-treated cells are dilution cloned, and successfully modified clones identified using routine molecular biology techniques. Cell lines that are difficult to transfect, such as primary cells, can be genetically modified via transduction of ZFNs.
Animals can be genetically modified by microinjection of embryos with ZFNs, transfection of embryonic stem (ES) cells or somatic cell nuclear transfer of ZFN-modified cells. Since the initial design of ZFNs more than 15 years ago, they have been utilized to modify the genomes of a variety of industrially relevant organisms and standard research models, including zebrafish, rats, mice, pigs, cattle, rabbits, and catfish. 1 –9
Strategies for Finfish Aquaculture
A primary research focus at the Center for Aquaculture Technologies is blocking sexual maturation differentiation, and ultimately controlling reproduction in farmed fish. 10 A prerequisite for the commercial deployment of genetically engineered fish will likely be reliable sterilization, which would eliminate the risk of impacting wild populations and prevent the establishment of feral populations. Sterile fish would likely also demonstrate greater performance in culture by conserving energy for growth rather than gonad development and sexual differentiation. 11,12 Monosex populations are advantageous in species displaying sexual dimorphism in economically important traits such as growth rate, body shape, coloration and carcass composition. 12,13
While strategies for gender manipulation and sterilization currently exist, they are often only applicable to a few fish species, incur significant cost in labor and equipment, or otherwise remain impractical at the commercial scale. 12,14,15 A preferred approach would be to apply targeted genome editing with ZFNs to inactivate key hormones involved in these physiological processes. Fish are an ideal host for ZFN genome editing because of their relatively short generation time, external fertilization, and the potential for efficient micro-injection procedures for germplasm delivery.
In collaboration with Sigma Life Science using CompoZr™ ZFNs, we tested multiple ZFNs for gene knockout in finfish embryos (zebrafish and tilapia). These ZFN-tailored gene knockouts should enable the production of sterile fish and monosex populations. These fish will be bred in captivity with corrective hormone supplementation, ensuring reversible control over sterility or sex specification. The successful deployment of such reproductive containment strategies will facilitate development of other ZFN-engineered improvements such as those listed in Table 1.
Commercially Valuable Applications of Targeted Gene Editing in Finfish Aquaculture
Presented are many of the value-added traits for commercial finfish aquaculture and the corresponding physiological pathways amenable to ZFN-tailored genetic engineering. Genetic modifications may include gene knockout, modified expression of existing genes, expression of modified alleles, or expression of new genes (transgenic).
These new opportunities are welcome in light of increasing demand for seafood, global challenges associated with overexploitation of most wild fish stocks, limited resources of coastal habitats, high competition for water between crop, livestock, and fisheries, and the need to advance sustainable finfish aquaculture. 16,17
ZFN Design and Application
ZFNs contain two distinct domains, a zinc finger DNA binding domain and the FokI restriction endonuclease domain (Fig. 2). The zinc finger DNA binding domain is comprised of zinc finger proteins that each recognize and bind to three DNA nucleotides. The modularity of the proteins enables linkage of 4 to 6 zinc finger proteins together, creating a protein that specifically binds 12 to 18 nucleotides within a genome.
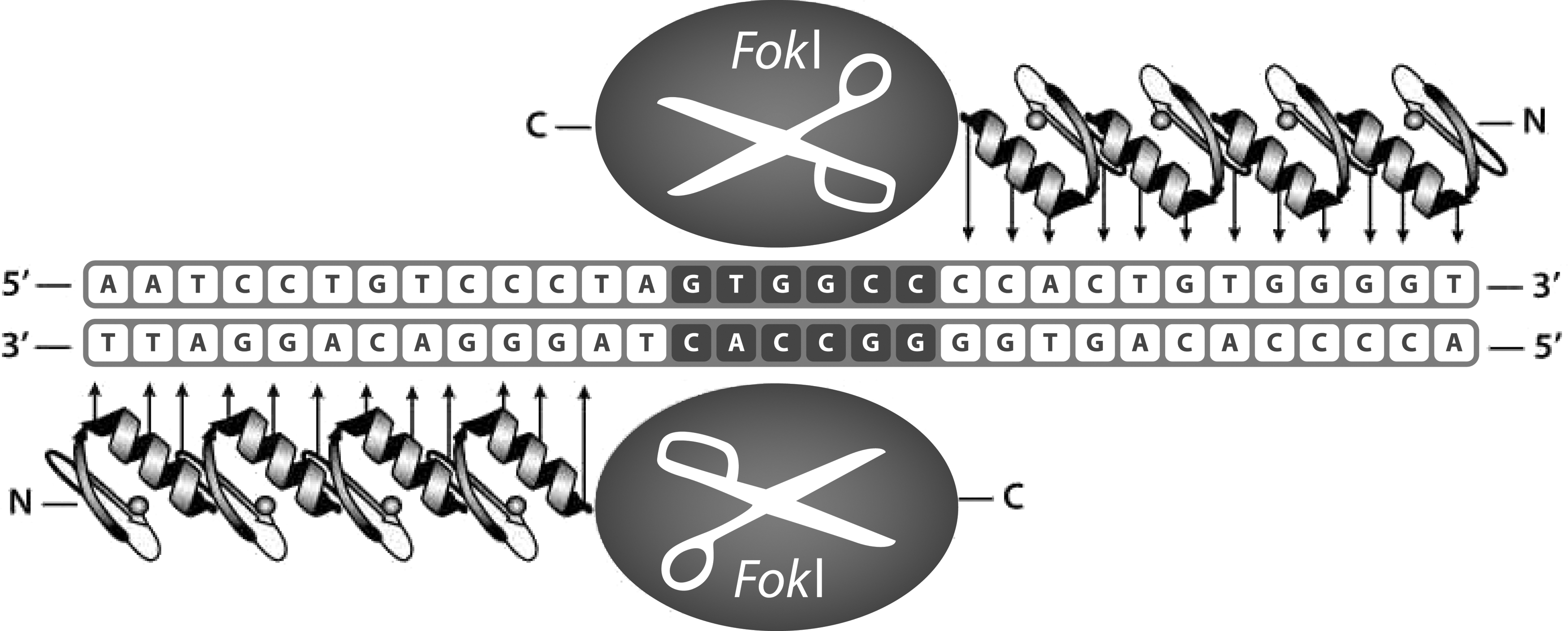
ZFNs are comprised of two functional domains: the FokI cleavage domain and the user-specified DNA binding domain.
The FokI endonuclease domain cleaves DNA to create a DSB. As the FokI endonuclease operates as a dimer, ZFNs are designed in “left-” and “right-handed” pairs that bind upstream and downstream of the DNA cleavage site. The first-generation “wildtype” FokI endonuclease allows two left- or right-handed ZFNs to homodimerize, which can cleave genomic DNA at off-target locations and generate undesired phenotypic outcomes. CompoZr ZFNs utilize an engineered “eHiFi” FokI domain that mandates heterodimerization between the left- and right-handed ZFN pairs. This creates a highly specific 24 to 36 nucleotide long DNA recognition and cleavage site that reduces the potential for undesirable off-target effects.
ZFNs are delivered to a cell in either DNA or mRNA format. Once translated, the ZFN translocates to the nucleus, binds to the appropriate target site and creates the desired DSB. Living cells use two pathways to repair DSBs: non-homologous end joining (NHEJ) and homology directed repair (HDR) (Fig. 3). In the absence of a repair template, the cell utilizes NHEJ. In a percentage of cases, NHEJ can be an imperfect repair process, leading to nucleotide insertions or deletions (indels) at the target site. Introduction of an indel can create a frameshift within the coding region of the gene, disrupting gene translation and knocking out gene function.
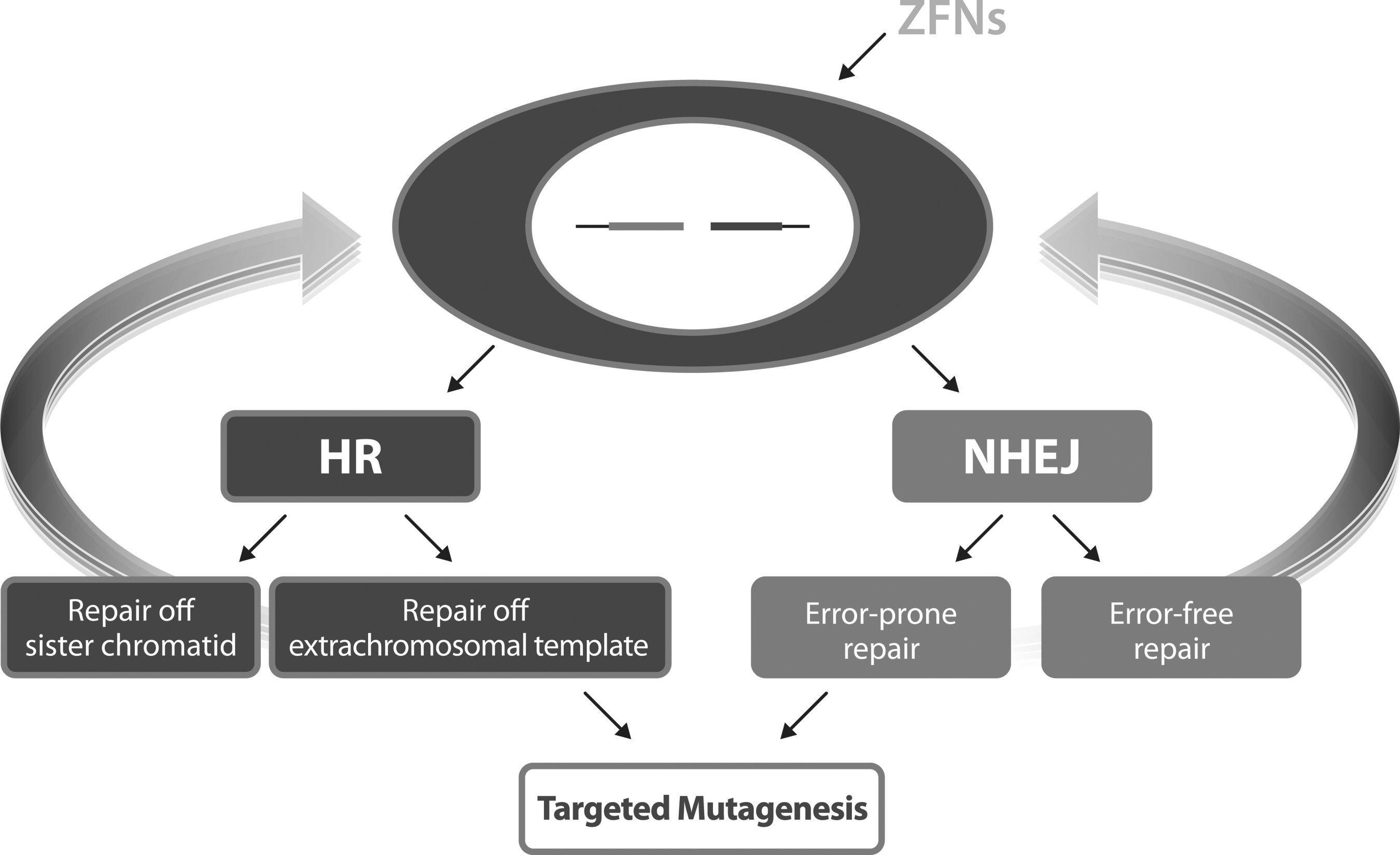
A ZFN-induced double-stranded break is repaired by NHEJ or HDR. NHEJ is used for gene knockouts while HDR is used for gene integration and/or modification.
If a donor plasmid with homology arms that immediately flank the ZFN target site is delivered with the ZFNs, the cell will likely utilize HDR as the repair process. The donor plasmid can be designed to include a variety of genetic elements that allow gene modification or gene integration. Potential modifications include integration of a transgene, tagging of the endogenous gene, introduction or correction of a SNP or any other modification of interest.
To assure success with CompoZr ZFNs, a genome-wide bioinformatics analysis is conducted prior to assembly of the ZFNs. This analysis prevents the manufacturing of ZFNs that could target areas of a genome with sequence homology, SNPs, splice variants, or repeat elements. The overwhelming majority of CompoZr ZFNs contain target sites of at least 30 nucleotides and a minimum criterion that each target site differs from any other genomic locus by at least four nucleotides to prevent off-target DNA cleavage.
Conclusions
The abundance of and easy access to fish embryos makes early genotyping possible, allowing rapid optimization of ZFN design and injection protocols, and early selection of ZFN-treated animals for phenotypic characterization. ZFN induced gene knockout has proven to be very successful across fish species. 3,9,18,19 Efforts to achieve ZFN directed site-specific DNA integration in the finfish genome continue. Nonetheless, the ease of use and effectiveness we and others have observed are convincing and supportive of a movement toward commercially relevant applications.
Genome editing with ZFNs has utility across industrial applications that use cell culture or animal models for research and development and/or manufacturing. In addition to fish, efficient genome editing of large domestic animals, including cattle and pigs, has been demonstrated with ZFNs. The increase in efficiency with ZFN-mediated genome editing in such animals is significant due to the time required and associated costs of animal husbandry. Improved production of high-strength biopolymers can also benefit from genome editing, exemplified by advances in the production of spider silk in silkworms. These results indicate that implementation of ZFN-mediated genome editing will increase efficiency and enable technological advances across a variety of industrial biotechnology applications.