
Abstract
A group of proteins including expansins, swollenins, and polysaccharide monooxygenases are non-hydrolytic cellulose active proteins (NHCAPs), which have been shown to enhance significantly cellulase-driven plant cell wall hydrolysis at reduced protein loadings. NHCAP-mediated enhancement of cellulase cocktails makes them important accessories in the molecular toolbox for producing fuels and chemicals from lignocellulose. This review presents an overview of the fundamental research advances involving these groups of proteins and considers their applicability to processes in the ongoing pursuit of an economically viable biorefinery.
Introduction
The search for sustainable, cleaner fuels and chemicals has intensified efforts to produce them from lignocellulose—a quest that began more than five decades ago. 1 Lignocellulose is comprised of cellulose (40–50%), hemicellulose (25–35%), and lignin (15–20%); the variation in relative abundance of these constituent polymers depends on the type of biomass. Within lignocellulose, linear chains of β-1,4-linked D-glucose units are held tightly by intramolecular and intermolecular hydrogen bonding between hydroxyl groups and oxygen atoms present in the macromolecule to form compact, crystalline cellulose microfibrils. 2
Efficient enzymatic hydrolysis of lignocellulose to monomeric building blocks is essential in the commercialization of second-generation liquid transportation biofuels and high-value chemicals. In nature, fungal saprotrophs and phytoparasites secrete complex cocktails of lignocellulose-degrading enzymes that function synergistically in plant cell wall deconstruction.
3
–5
These include the carbohydrate active enzymes (CAZymes;
The crystallinity of cellulose and its association with hemicellulose and lignin severely impair its susceptibility to enzymatic hydrolysis (saccharification). 12 Other key factors that contribute to inefficient hydrolysis include non-productive adsorption of enzymes by lignin and other structural components and the presence of naturally occurring enzyme inhibitors. 12 These factors increase processing costs. 13 Recent cost estimates for enzymes range between $0.30 and $0.50 per gallon of ethanol, making enzymes the second largest contributor (behind feedstock) to operating costs in the lignocellulose to ethanol bioprocess. 14,15
Research efforts to improve the efficiency of enzymatic saccharification of lignocellulosic biomass are broadly based and include the following: development of energy crops that are less resistant to enzymatic hydrolysis; development of a broad spectrum of lignocellulosic biomass pretreatment methods that enhance enzymatic accessibility to cellulose; engineering enzymes with improved catalytic efficiency (site-directed mutagenesis, directed evolution, construction of chimera or cellulosomes, and synergy engineering); searching nature for more efficient enzymes and accessory proteins; and designing synthetic enzyme cocktails with improved cellulose hydrolyzing capabilities. 16 –25
Significant improvements in lignocellulose hydrolysis have been obtained by the inclusion of auxiliary hydrolytic enzymes such as acetylxylan esterases, arabinofuranosidases, xylosidases, mannanases, and pectinases in core lignocellulolytic enzyme cocktails. 22,24,25 However, the resultant catalytic improvements are limited, possibly due to physical barriers imposed by insoluble or highly ordered polymer packaging in lignocellulosic biomass. High-level synergism may be possible in the presence of auxiliary proteins that can access insoluble biomass and disrupt highly ordered polymer packaging, thereby facilitating attack by hydrolytic enzymes.
A group of non-hydrolytic cellulose active proteins (NHCAPs) including expansins, swollenins and polysaccharide monooxygenases—previously known as glycoside hydrolase family 61 (GH61) proteins—have been shown to act synergistically with cellulases in lignocellulose hydrolysis. 26 –29 Unlike the core cellulases, for which synergistic mechanisms have been extensively studied, NHCAPs work concertedly with cell wall hydrolases via mechanisms that are currently poorly understood. 30 Due to their potential ability to enhance lignocellulolysis, NHCAPs are attracting interest as prospective catalytic components in biorefineries. This article presents an overview of the fundamental research advances on NHCAPs and their potential impact on the commercialization of lignocellulose-based fuels and chemicals.
Classification and Structure of Non-Hydrolytic Cellulose Active Proteins
Classification of Expansins
Expansins are thought to be present in almost all terrestrial plants ranging from bryophytes to angiosperms. 31 Two expansins and their encoding genes were originally isolated from cucumber cell walls. 32,33 Since then, other plant expansins and expansin-like proteins from organisms such as bacteria, fungi, and nematodes have been identified and characterized.
Expansins and expansin-like proteins are classified into families using the systematic nomenclature shown in Table 1. 34,35 The four main plant expansin families are designated EXPA (α-expansins), EXPB (β-expansins), EXLA (Expansin-like A) and EXLB (Expansin-like B). In this classification, expansin-like family X (EXLX) is the fifth and newest expansin family and represents the sub-family of expansin comprising non-plant proteins with distant homology to EXPAs and EXPBs. 34 These include the swollenins discussed below.
Reprinted with permission from Springer Science+Business Media B.V.
As listed in reference 31.
Structural Properties of Expansins
Expansins contain between 250–275 amino acids, with limited sequence conservation between expansins of different families (20–40%). 34 –36 A secretory signal peptide directs export to the cell exterior. With the exception of loosenins, described below, expansins comprise two distinct domains: an N-terminal catalytic domain (domain I) and a C-terminal tryptophan-rich putative polysaccharide binding domain (domain II) interconnected by a short linker, according to a review by Kende et al. 34,37 The crystal structures of two expansins have been solved. 38 –40 Domain I, which is organized into a double psi beta barrel (DPBB) fold, is structurally related to the catalytic domain of GH45 proteins (mainly fungal β-1,4-D-endoglucanases), whereas the C-terminal domain II is related to group II pollen allergen of grasses. 38,39,41 Domain II of an expansin-like protein (BsEXLX1) lacks any significant sequence similarity with members of the 62 currently known carbohydrate-binding module (CBM) families in the CAZy database, and was proposed as the founding member of CBM family 63. 38
Classification and Structural Properties of Swollenins
Swollenins are classified as members of the EXLX family. The first swollenin characterized was isolated from the filamentous fungus Trichoderma reesei. 42 Like expansins, swollenins have a catalytic domain I and cellulose-binding domain interconnected by a linker region. 42,43 However, a single domain EXLX (loosenin) from the white-rot basidiomycete Bjerkandera adusta, BaLOOS1, lacks the carbohydrate-binding C-terminal domain II present in plant expansins, with both cell-disrupting and polysaccharide-binding activities bundled in domain I. 37
Structure of Polysaccharide Monooxygenases
Polysaccharide monooxygenases (PMO) constitute a group of small (22–45 kDa) proteins previously classified as family 61 glycosyl hydrolases (GH61). It was recently shown that PMOs are Cu-dependent metalloenzymes that oxidatively depolymerize crystalline cellulose in the presence of small molecule redox active agents such as gallate and ascorbate or the enzyme cellobiose dehydrogenase. 28,44 –46 These proteins are structurally similar to chitin-binding proteins, such as CBP21 from Serratia marcescens and CelS2 from Streptomyces coelicolor, belonging to the carbohydrate-binding module family 33 (CBM33), and which can oxidatively cleave polysaccharides. 28,47 –51
Despite the structural and functional similarity of PMOs with CBM33 proteins, there is little evidence that these enzyme families share phylogenetic ancestry. PMOs appear to be restricted to fungi, while CBM33 proteins are found mainly in bacteria and viruses. 52 –54 Within the fungal kingdom, PMOs are widespread and occur in organisms with different ecological lifestyles and nutritional modes. Most cellulolytic fungal species harbor multiple PMO genes. For instance, the T. reesei genome contains six PMO genes whereas that of Thielavia terrestris harbors 18; the highest number so far (>40) has been detected in Chaetomium globosum. 55 –57 White-rot fungi tend to contain more copies of PMO genes than brown-rot fungi. 58
A recent analysis of 143 PMO genes indicated that 26% of the encoded PMOs are associated with CBM1—a domain that is involved mainly in cellulose binding—and two are associated with the chitin-binding CBM18, according to a review by Lo Leggio et al. 59,60 The Cel2S protein from Streptomyces coelicolor contains both a CBM33 domain and a CBM2 domain and is active in cellulose cleavage. 51,60
As of February 2013, there were 249 GH61 (PMO) entries in the CAZy database. 7 Of these, five structures have been solved: polysaccharide monooxygenases PMO–2 and PMO–3 from Neurospora crassa, GH61A from Thermoascus aurantiacus, GH61E from T. terrestris, and GH61B from T. reesei. 60 None of these five PMO structures is associated with a CBM1 and none shows any evidence of a binding pocket for soluble polysaccharides, ie, large surface clefts, crevices, or holes that are commonly found in or proximal to the active sites of conventional glycoside hydrolases. Instead, PMOs (and CBM33 proteins) present planar surfaces that appear to be suitable for binding to crystalline polysaccharide surfaces, according to Lo Leggio et al. 60 In the known PMO structures, a divalent metal ion binding site that preferentially binds Cu2+ is formed by an essential methylated N-terminal histidine residue, together with a second essential histidine residue and a tyrosine side chain. 28,46,60 For more information, see the review on the structure of PMOs by Lo Leggio et al. 60
Classification of Polysaccharide Monooxygenases
At least two types of PMOs have been functionally distinguished thus far. Type-I PMOs incorporate one oxygen atom from O2 into C-H bonds adjacent to glycosidic linkages generating products oxidized on C1 (reducing end), while Type-II PMOs yield products oxidized at C4 (non-reducing end) (Fig. 1). 60,61
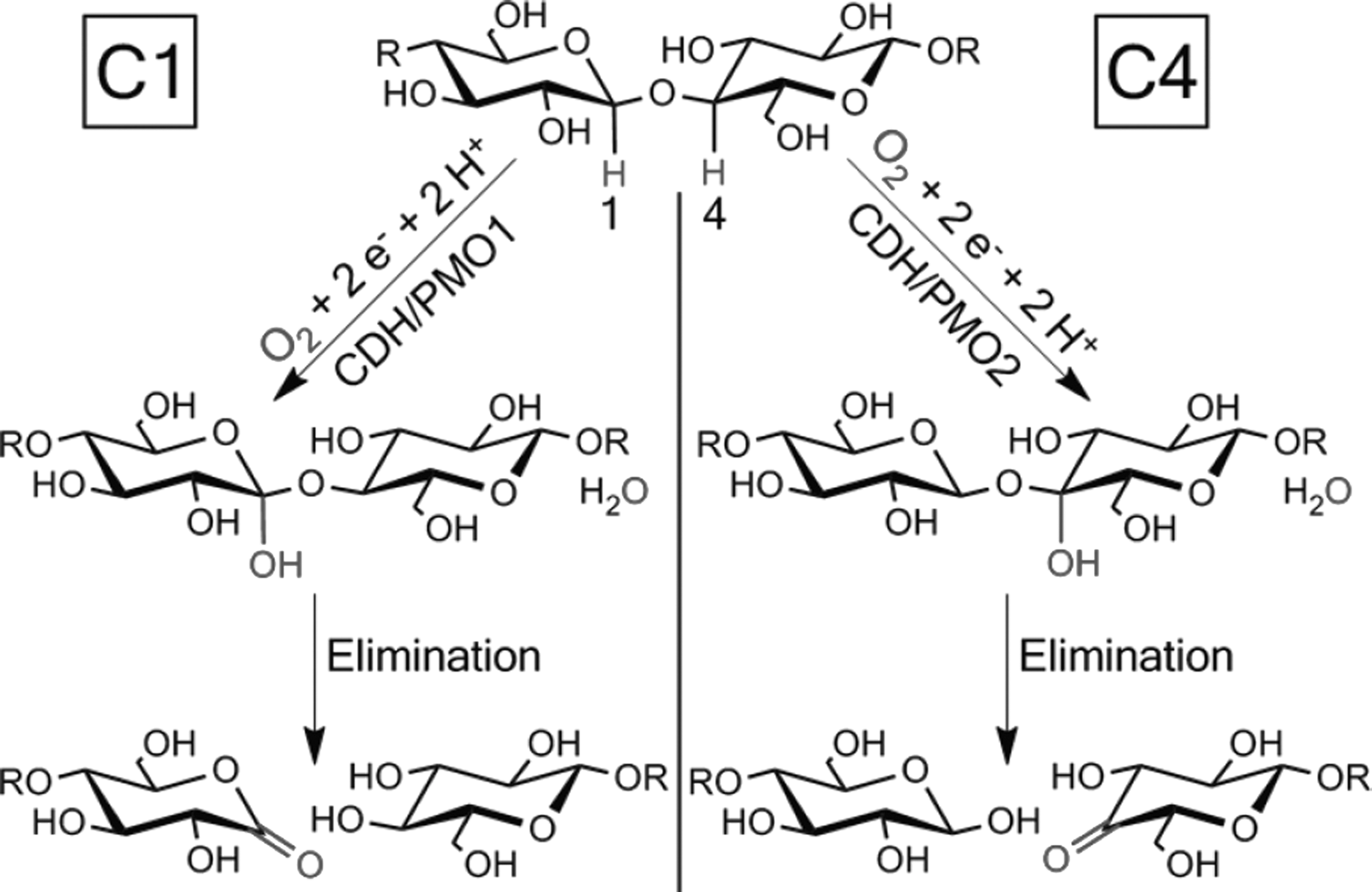
Proposed reaction pathway for CDH-assisted oxidative cleavage of cellulose by GH61 (PMO). Note that the oxidation of carbon C1 or C4 is spontaneously followed by a non-reversible elimination reaction, which produce aldonic acid and 4-ketoaldose, respectively. 61 (Reprinted with permission; copyright 2012 American Chemical Society.)
Activities of Non-Hydrolytic Cellulose Active Proteins
Expansin Activity
Expansins loosen plant cell wall compactness and disrupt cellulose crystallinity without themselves having cellulolytic or xyloglucan endotransglycosylase activities. 62 –64 Several studies have shown that expansins (BsEXLX1 and maize β-expansin) bind to cell wall polysaccharides. 38,65,66 Expansins are thought to loosen cell wall matrix polysaccharides by a mechanism involving the disruption of non-covalent bonds between cellulose microfibrils or cellulose and other matrix polymers. This allows cellulose microfibrils to slip or “creep” past each other within cell walls subjected to mechanical stress. For instance, decrystallization of cellulose by expansin involves binding of the protein onto H-bonded cellulose coated with other polysaccharides. As the protein drifts along the chain, disruption of intermolecular and intramolecular H-bonding occurs within the interacting matrix carbohydrates to facilitate slippage. 67,68
A recent report on the structure-function relationship of recombinant BsEXLX1 revealed that although both domains work together, domain I is the main catalytic domain responsible for "creep activity" (cell wall loosening), while domain II is responsible for binding to cellulose and whole cell wall matrix polysaccharides, indicative of a CBM. 38 Using site-directed mutagenesis, the same study pinpointed various residues involved in polysaccharide binding and wall loosening activities.
Regardless of their limited sequence identity, both EXPA- and EXPB-type expansins show similar plant cell wall loosening effects, are activated by reducing agents, and are active under mildly acidic (below pH 6) conditions, but not at neutral pH. 32,69 Evidence of the non-hydrolytic nature of expansins includes the fact that their cell wall loosening action is not time-dependent and their effects on the mechanical properties of the cell wall are fully reversible upon protein denaturation. 67 Some studies have shown that expansins are co-expressed with endoglucanase and xyloglucan endotransglycosylase in response to auxin, suggesting that these proteins may act synergistically in cell expansion. 70 –72 Differences in substrate specificity have been observed between EXPAs and EXPBs, with EXPA showing preference for dicot cell walls, whereas the EXPBs prefer those of grasses; grass cell walls are composed of unusual hemicelluloses (eg, mixed-link glucan and glucuronoarabinoxylan) in addition to the usual matrix polymers present in dicot cell walls (ie, xyloglucan, pectin, etc.). 73 Although the cell wall loosening activity of many members of the EXPA and EXPB families has been demonstrated, there is a dearth of experimental evidence for this activity in EXLA and EXLB family proteins.
Swollenin Activity
Swollenins are expansin-like proteins that cause loosening, partial disruption, and swelling of plant cell walls. 42 As is the case for expansin, the catalytic domain of swollenin harbors cell-wall polysaccharide-loosening activity that partially disrupts, loosens, and reduces cellulose integrity (decrystallization) without release of reducing sugars. 42,43 The original work on T. reesei swollenin characterization is complemented by binding studies of swollenins from other fungal species. Aspergillus fumigatus swollenin, AfSwo1, binds to microcrystalline cellulose (avicel) and chitin, shows negligible activity toward carboxymethyl cellulose (CMC) and avicel, and does not bind to xylan. 26 Swollenin 2 (Swo2) from Trichoderma pseudokoningii S38 shows slight hydrolytic activity on xylan and yeast cell wall glycan, but not on CMC, cotton fiber, filter paper, or cellulose powder CF11. 74
Polysaccharide Monooxygenase Activity
In the presence of molecular oxygen and reducing agents, PMOs are involved in the oxidative disruption of crystalline cellulose and chitin, tying their function to the structural homology they share with CBM33. 28,47 –49,60,75 Their mechanism of action and the nature of carbohydrate oligomers released (ie, oxidized cello- or chito-oligomers) are markedly different from those of canonical hydrolytic cellulases (ie, non-oxidized oligomers). 29,75 Although C1, C4, or, to a lesser degree, C6 in the glucose ring may be oxidized by PMOs, aldonic cellodextrins resulting from C1 oxidation are the most abundant products. 61,76,77 This oxidative cleavage of cellulose into a variety of native and oxidized cellodextrins with varying degrees of polymerization renders lignocellulosic biomass more susceptible to attack by classical endo- and exo-cellulases. 28,46,75
Synergy between PMO and the secreted fungal flavocytochrome cellobiose dehydrogenase (CDH) appears to promote cellulose degradation by canonical hydrolases. 54,75,78,79 Among other suggested roles, CDHs are thought to bind cellulose; oxidize cellodextrins, maltodextrins, and lactose to their corresponding lactones; and reduce Cu(II) to Cu(I) in the PMO catalytic cycle. 44,60,80 –82 Many fungal genomes contain PMO- and CDH-encoding genes that may be expressed and up-regulated together; a few others (eg, the brown-rot fungus, Postia placenta) have multiple PMO-encoding genes but none encoding CDH. 56,83,84
Cellulose Hydrolysis with NHCAP Additives
Expansins as Additives to Lignocellulolytic Enzyme Cocktails
The expansins most commonly used in research as additives to enzyme mixtures for lignocellulose hydrolysis are the EXLX family proteins. Unlike the plant expansin families, which have not been successfully expressed in non-plant host systems, most EXLX members are readily expressed heterologously in a diverse number of host systems including bacteria, fungi, and plants. 26,39,42 Consequently, the EXLX proteins are promising candidates for improvement of industrial-scale lignocellulose hydrolysis.
Several studies of cellulose cocktail enhancement have used the β-expansin-like protein from Bacillus subtilis (BsEXLX1). This protein showed a degree of synergism of 5.7 with low cellulase loadings (<0.6FPU/g cellulose) on filter paper, but no improvement was observed at higher cellulase loading. 26 Glucose yields were, however, too low for BsEXLX1 to be considered applicable in industrial lignocellulolysis. In a second study, BsEXLX1 and crude cellulase mixtures from Aspergillus and Penicillium species increased glucose, xylose, arabinose, and total reducing sugar yields from alkali pretreated wheat straw by 45–112% compared to the cellulase preparation Celluclast 1.5L alone. 85 By replacing 32% of cellulase with recombinant BsEXLX1, the total cellulase loading was reduced by 21%, with a net gain in glucose, xylose, arabinose and total reducing sugar yield of 10–13% each. Another expansin-like protein from Hahella chejuensis (HcEXLX2) was reported to confer a 4.6-fold enhancement in reducing sugar released from filter paper by Celluclast 1.5L, but no tests were performed on natural lignocellulose. 86
Cellulase loading often used for assessing digestibility of pretreated lignocellulosic biomass is 5 filter paper unit (FPU)/g glucan or higher, which exceeds loadings at which enhancement is observed for BsEXLX1 and HcEXLX2. 13,26,39,86,87 Preferential binding of recombinant BsEXLX1 to lignin, rather than cellulose, has led to the suggestion that bacterial expansins could be used as lignin blocker in the enzymatic hydrolysis of lignocellulose. Other substrate binding studies on BsEXLX1 conclude that the success of expansin dosing in biomass conversion is dependent on the substrate pretreatment method. 88
The first report using a plant expansin to reduce doses of T. reesei cellulase in the hydrolysis of microcrystalline cellulose was in 1998. 26 A small amount (0.1 mg of enzyme/g cellulose) of β-expansin-D from corn pollen was reported to enhance hydrolysis of acid-pretreated yellow poplar sawdust by a T. reesei cellulase mixture. 89 The improved rate of saccharification was observed in the glucan conversion range of 55–80%. When tested on a synthetic binary cellulase mixture containing Acidothermus cellulolyticus endoglucanase and T. reesei CBHI (8.35 mg total protein/g biomass), β-expansin-D did not enhance saccharification, which suggests that the cell wall loosening corn β-expansin-D has to interact with one or more component(s) in the T. reesei cellulase complex mixture missing in the synthetic binary mixture. 89
Wei et al., studied synergism between an α-expansin from cucumber and a mix of purified fungal cellulases, hemicellulases, and pectinases. 90 Cucumber α-expansin shows concentration-dependent enhancement of hydrolysis of homogeneous model substrates (pectin and carboxymethylcellulose), but it is difficult to extrapolate these results to complex lignocellulosic biomass. 90 Moreover, all the previous studies that employed expansins as additives in lignocellulolytic enzyme cocktails used biomass concentrations lower than 2%, in contrast to the high loadings required in industrial bioprocesses. 91
Swollenins as Additives to Lignocellulolytic Enzyme Cocktails
Swollenins have also been reported to enhance cellulase activity synergistically. One study reported increased release of reducing sugar from endoglucanase hydrolysis of cotton fiber pretreated with BaLOOS1, the increase of which was proportional to the amount of BaLOOS1 used. 37 When tested with a cocktail of commercial cellulases and xylanases on lignocellulosic substrate (Agave tequilana bagasse), BaLOOS1 rendered the biomass 7.5 times more susceptible to enzymatic hydrolysis. Nonetheless, BaLOOS1 is inactive at temperatures above 40°C, limiting its use in lignocellulolytic bioprocesses designed for higher temperatures. 37
The effects of a number of swollenins on hydrolysis of model cellulose substrates have been reported. Pretreatment of filter paper and avicel by T. reesei swollenin, produced in Kluyveromyces lactis, was found to enhance cellulase hydrolysis of both substrates. 92 However, T. reesei swollenin is produced only at a low level by T. reesei (≤25 μg/L), limiting its usefulness for industrial applications. 42,43 Trichoderma asperellum swollenin enhances hydrolysis of avicel by endoglucanase from Fervidobacterium nodosum. 93 Recombinant Aspergillus oryzae swollenin promotes cellulase hydrolysis of filter paper by up to 80% in a concentration-dependent fashion. 43 A concentration-dependent increase (61–74%) in cellulase-mediated saccharification of crystalline cellulose was observed when 20μg AfSwo1/g cellulose was added to a commercial cellulase mixture from T. reesei and Aspergillus niger. 26
Some swollenins appear to exhibit low but significant hydrolytic activity toward polysaccharides, making it difficult to determine whether it is the wall loosening or the hydrolytic activity that interacts synergistically with cellulases in lignocellulosic biomass hydrolysis. 27
Polysaccharide Monooxygenases in Lignocellulose Deconstruction
The current generation of commercial cellulase cocktails are known to contain PMOs. Cellic CTec2, manufactured by Novozymes A/S (Bagsværd, Denmark), contains extra PMOs that have significantly contributed to the improved performance compared to its predecessors, notably Celluclast 1.5L. 77 Coexpression of T. terrestris or T. aurantiacus PMOs with cellulases in a commercial hypercellulolytic T. reesei strain leads to a 2-fold reduction in the total protein loading and cost of hydrolysis of acid-catalyzed steam-exploded corn stover. 28 Using the T. aurantiacus PMO as model protein, Harris et al. concluded that stimulatory effects of PMOs on cellulases are not due to interaction with any single specific enzymatic activity, but rather with the overall activity of the cellulase complex or substrate. 28
The cellulase enhancement activity of PMOs seems to be influenced by the presence of other cell-wall-derived material such as lignin or hemicellulose present in the substrate. 28 For instance, lignin boosted the cellulase-enhancing performance of a recombinant Myceliophthora thermophila PMO in the degradation of hydrothermally pretreated wheat straw, soda-pretreated spruce/pine pulps, and acid-catalyzed steam pretreated spruce. Up to 40% enhancement was achieved on the cellulase hydrolysis of pretreated spruce. 94 A mixture containing 10.5 mg of M. thermophila PMO and 16.5 mg mixture of purified cellulases (a combination of endoglucanase, cellobiohydrolase, and β-glucosidase) was found to have the same hydrolyzing activity on pretreated lignocellulosic biomass as 33 mg of purified cellulolytic enzymes—the equivalent of 20% reduction in enzyme loading. 94 Canella et al., used pretreated wheat straw as representative lignocellulosic biomass and filter paper as pure cellulosic substrate to demonstrate that T. reesei PMO additives in Cellic CTec2 does not require any reducing co-factor for activity in the presence of lignin, implying that lignin can function as electron donor for PMO activity. 77 This also suggests that PMOs may exploit adventitious copper and reductants present in natural lignocellulosic biomass for their activity, making such metal-induced lignocellulolytic enhancement feasible at low cost. The role of PMOs in the degradation of cellulose by fungal enzymes is represented in Fig. 2. 54,61
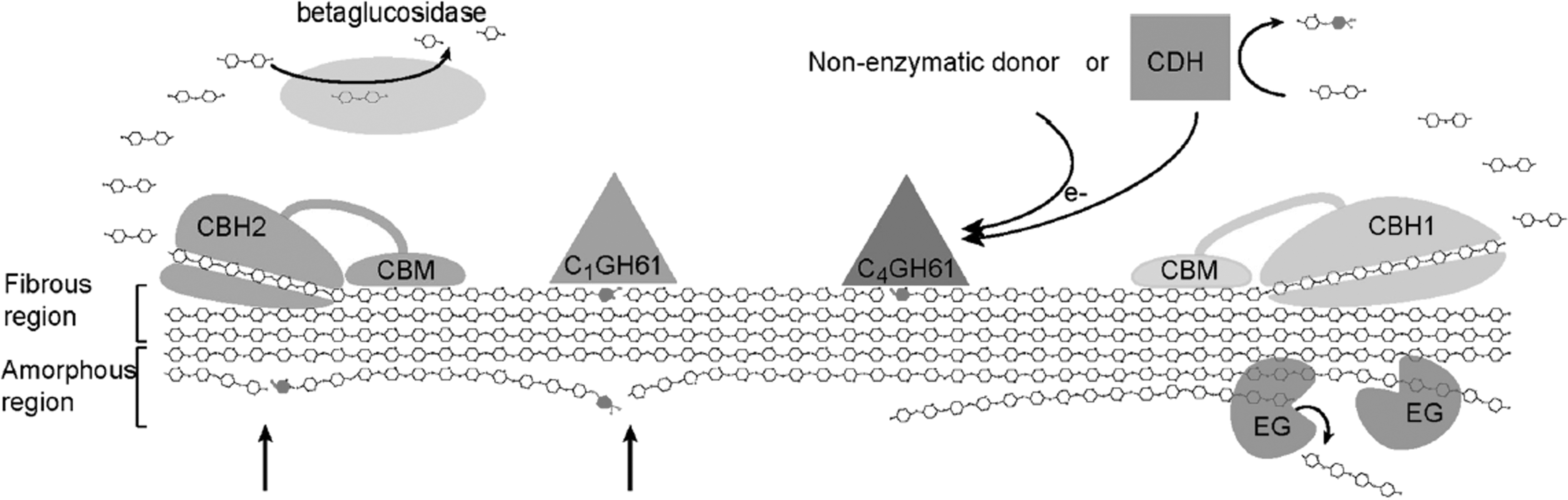
Illustration of the current understanding of fungal enzymatic degradation of cellulose, showing a C1 and a C4 oxidizing GH61, which generates non-oxidized ends that serve as new points of attack for cellulobiohydrolases I and II (arrows); oxidized sugars are shown in black. 54 Like non-enzymatic electron donors such as ascorbic acid, gallic acid, and reduced glutathione, CDH plays the role of reductant in GH61 activity. (EG, endoglucanase; CBH, cellobiohydrolase; CDH, cellobiose-dehydrogenase; CBM, carbohydrate-binding module). Reprinted with permission from BioMed Central.
One challenge associated with the use of PMO additives in cellulase mixtures for the deconstruction of lignocellulose is the production of non-fermentable gluconic and cellobionic acids (Fig. 3). 77 Cannella et al., found a positive correlation between the gluconic acid yield and cellulose conversion when Cellic CTec2 was used to hydrolyze pretreated wheat straw at high (30%) solids loading. Unlike CTec2 which contains PMO additives, Celluclast 1.5L supplemented with Novozyme 188 does not produce any detectable gluconic acid from the same substrate. 77 Gluconic acid was found to be a more potent β-glucosidase inhibitor than glucose, while Cellic CTec2 was found to hydrolyse cellobionic acid at a much slower rate than cellobiose. In the same study, gluconic acid production was found to be lower at higher temperatures without compromising the overall cellulose conversion. 77 This suggests that temperature could be used, to a certain extent, to control aldonic acid levels and its inhibitory effect on β-glucosidase.
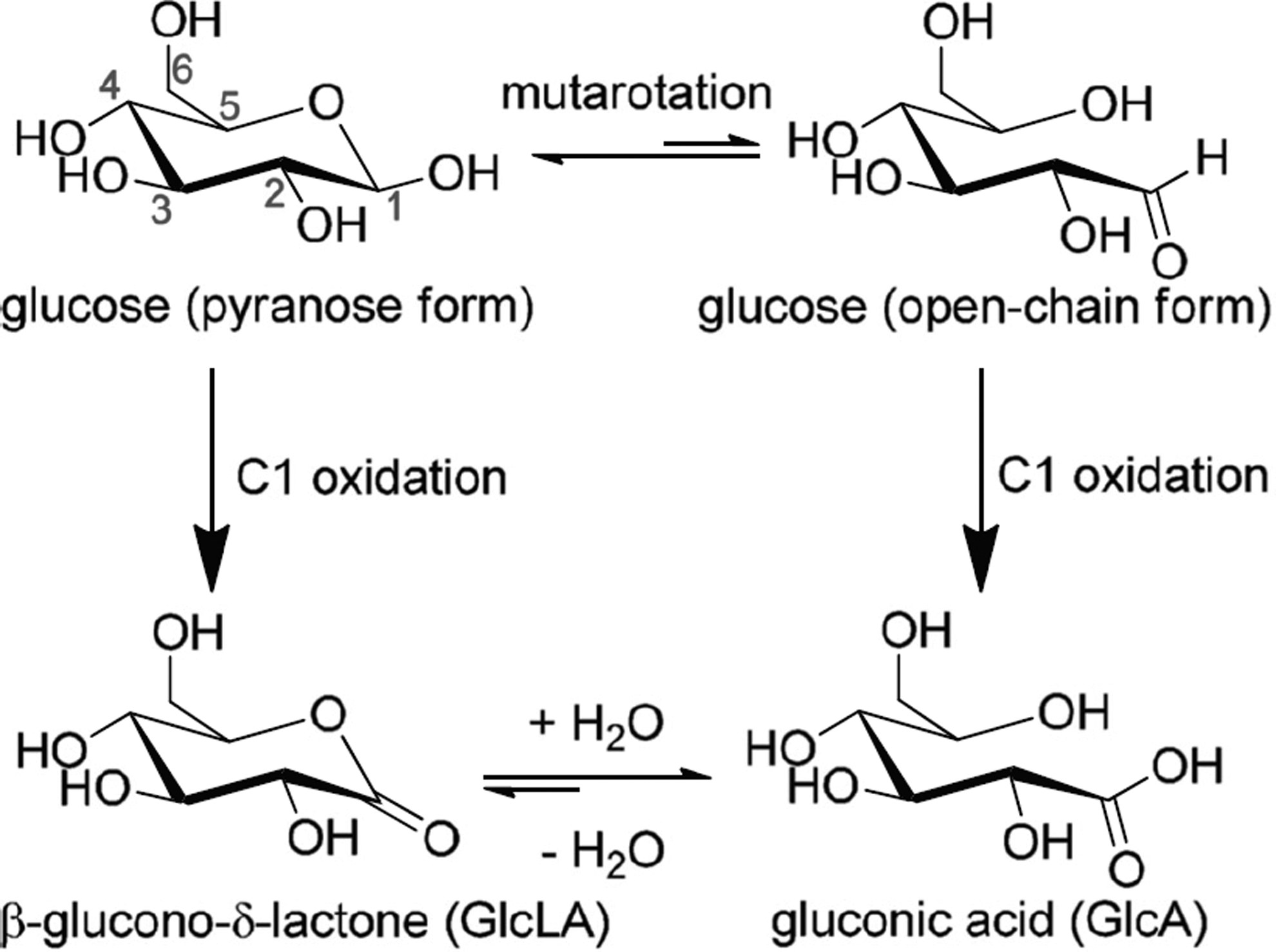
Gluconic acid formed from C1 oxidation of glucose.7 Reprinted with permission from BioMed Central.
Other Non-Hydrolytic Cell Wall Active Proteins
Other non-hydrolytic cell-wall active proteins have been reported to cause expansion or degradation of plant cell wall polysaccharides. Two non-hydrolytic proteins that regulate cell wall expansion in cowpea hypocotyl were reported in 1995. 95 However, details of the description and characterization of these proteins are not available. Another non-hydrolytic protein termed “acid wall protein” was found to stimulate activity of exo- and endoglucanases in maize coleoptile walls. 96 It is not clear, however, whether the action of the “acidic wall protein” renders cell wall polymers more accessible to hydrolysis by exo- and endoglucanases. A cellulose-induced protein Cip1, was identified in T. reesei. 97 The fact that the sequence of Cip1 does not fit into any of the known GH families, that it is co-expressed with canonical cellulases, and that it contains a cellulose-binding module suggests that it may play a non-hydrolytic role in biomass degradation. 97
Conclusions
This review highlights the growing research interest on NHCAPs, particularly regarding their potential application in commercial-scale enzymatic biomass conversion. Fundamental research progress on NHCAPs has been significant in recent years, but much remains to be learned. For instance, due to the large superfamily size of expansins, the expression, regulation, and functional roles of many expansin genes have yet to be explored, which might unlock vital information toward the use of these proteins in biomass degradation. While the mechanistic reactivity of a number of PMOs has been clearly established as copper- and reductant-dependent polysaccharide (cellulose and chitin) monooxygenases, sequence diversity in the family is large, and how similar the as-yet uncharacterized PMOs are remains to be seen. Furthermore, many of their catalytic properties (eg, pH and temperature optima and specific activity) have not yet been extensively studied. Finally, it is still unclear on which specific polymers expansins and swollenins act to loosen the cell wall, and what molecular mechanisms are involved.
Very few studies have focused on the direct use of NHCAPs as booster additives in enzyme cocktails for lignocellulosic biomass degradation. With the exception of studies leading to the development of the Cellic CTec enzyme family, the small number of studies that have tested NHCAPs for lignocellulosic biomass hydrolysis have used relatively homogeneous model substrates (such as avicel, phosphoric acid swollen cellulose, and filter paper), which do not match the complexity of natural lignocellulosic biomass. In addition, these studies usually employed solids loadings of 5% or less, which is far below the loadings in excess of 15% required in an industrial setting, or have used thermolabile NHCAPs (eg, BaLOOS1, tested by Quiroz-Castañeda et al.). 37,91 To overcome the challenges related to enzyme inefficiency in lignocellulose degradation, research on NHCAPs could be focused on thermostable NHCAPs with tolerance to high solids loading. High temperatures facilitate biomass solubilization, while high solids loadings ensure high product yields at low operational cost. Thermophilic fungal saprotrophs represent an interesting, rich gene reservoir from which such proteins could be sourced. Several research groups are now focusing on such thermophilic fungal species for the development of improved lignocellulolytic enzyme cocktails.
Footnotes
Acknowledgments
Research on lignocellulose-active proteins at Concordia University is supported by funds from Genome Canada, Génome Québec, and the Bioconversion Network of the Natural Sciences and Engineering Research Council of Canada.
Author Disclosure Statement
The authors collaborate on related research with DSM and Iogen Corporation but do not have financial interests in these companies.