
Abstract
β-Glucosidases have great potential to be used in various biotechnological processes, such as those used by the food and feed industries, biomass hydrolysis for bioethanol production, and enhancing the flavor of wine, tea, and fruit juice. This work describes β-glucosidase production by Penicillium miczynskii. The effects of pH, time, and condition of cultivation, temperature, different carbon sources, and concentration of the selected agro-industrial waste on extracellular β-glucosidase production were studied in submerged fermentation. Additionally, the enzyme produced was biochemically characterized. The best conditions for P. miczynskii β-glucosidase production were 3% pineapple peel as carbon source, under stationary conditions, for 9 days, at pH 5.5 and temperature of 20°C, which yielded 2.82 U/mL. The enzyme showed best activity at pH 4.5–5.0, and 65°C. It was stable at 55°C and 60°C, with a half-life of 50 min and 40 min, respectively. The β-glucosidase was slightly activated in the presence of β-mercaptoethanol, Ca2+, and Co2+, and strongly inhibited by sodium dodecylsulfate (SDS), Zn2+, and Hg2+. Also, sodium citrate, NH4 +, and Ba2+ were able to inhibit this enzyme, especially at 10 mM concentration. The enzyme presented high glucose tolerance, with a Ki of 760 mM. This study describes a novel β-glucosidase that presents favorable industrial properties, such as low inhibition rate by glucose, and production on pineapple peel, an inexpensive and abundant agro-industrial waste.
Introduction
Biofuel technology is now globally considered a promising technology to replace fossil fuels. Lignocellulosic biomass from forestry, agricultural, and municipal sources are abundant and inexpensive potential feedstock for bioenergy production. To initiate lignocellulosic energy production, saccharification of cellulosic biomass is essential. 1,2 Bioconversion of lignocellulosic residues is initiated primarily by microorganisms such as fungi and bacteria, which are able to degrade these wastes. In filamentous fungi, cellulases include endoglucanases (Enzyme Commission number 3.2.1.4), cellobiohydrolases (3.2.1.91), and β-glucosidases (3.2.1.21), which efficiently act on cellulolytic wastes in a synergistic manner. 3
β-glucosidases hydrolyze soluble cellobiose into two glucose molecules. This enzyme has been isolated from many fungal species including ascomycetes and basidiomycetes. 3,4 Industrial applications of β-glucosidases include the paper, food, and feed industries; bioethanol production; and oligosaccharide synthesis for enhancing the flavor of wine, tea, and fruit juice. 5,6
Frequently, β-glucosidase is a rate-limiting factor during enzymatic hydrolysis of cellulose and is very sensitive to D-glucose inhibition. 7,8 Recently, the search for glucose-tolerant β-glucosidases has increased significantly, since these enzymes can improve the process of saccharification of lignocellulosic materials. To date, a few microbial β-glucosidases have been reported to tolerate glucose. 9 –12
The major limitation for broader applicability of industrial cellulases is their high cost. A successful strategy for producing these enzymes can be achieved through microorganism selection, since different fungi produce a variety of enzymes. Other strategies include screening using various lignocellulosic materials such as agro-industrial wastes and the improvement of fermentation process conditions. 6,13,14
Pineapple waste is a by-product of the processing industry and consists of residual pulp and peels. This lignocellulosic material still retains a considerable amount of soluble sugars, as well as high fiber and low protein content. 15,16 A high volume of agro-industrial waste is produced in Brazil due to the country's abundant agricultural activity. Brazil is the largest pineapple producer in the world, reaching 2,205,590 tons in 2010. As a consequence, a large quantity of pineapple waste is generated. Like many other agro-industrial by-products, pineapple waste has low commercial value, and its deposition on a large scale is a serious environmental problem. In many research centers, efforts to find alternative uses for organic material generated by the agro-industrial sector are ongoing. 15 –18
Penicillium miczynskii is a widely distributed fungus isolated from a variety of foodstuffs as well as from soil. 19 Although β-glucosidase from different fungal species has already been characterized, there are no reports about the β-glucosidase produced by this species. The aims of this study were to investigate the influence of the chemical-physical parameters on β-glucosidase production by P. miczynskii and to characterize biochemically the enzymes produced.
Material and Methods
Fungal Strain and Culture Conditions
The P. miczynskii used in this investigation was isolated from the Atlantic forest on Ecologic Station Juréia-Itatins, located in São Paulo State, Brazil. The strain belongs to the Culture Collection of the Environmental Studies Center at Universidade Estadual Paulista, Brazil. The fungus was grown on Vogel's solid medium containing 1.5% (m/v) glucose and 1.5% (m/v) agar, at 28°C, for 7 days for conidia production. 20 Submerged fermentation was prepared in 125-mL flasks containing 25 mL of the Vogel liquid medium at pH 6.5 and inoculated with 1.0 mL spore suspension (1.0×107 spores/mL). The cultures were incubated at different conditions described below. After incubation, cultures were filtered by vacuum. The filtrate was assayed for extracellular activity and protein. The mycelium was frozen and macerated with sand in McIlvaine buffer pH 5.0. The slurry was centrifuged at 3.900×g, for 15 minutes, and the supernatant was used as an intracellular protein source. All experiments were carried out in duplicate to verify their reproducibility, and the results are presented through mean values.
Enzyme and Protein Assays
β-glucosidase activity was determined by measuring the amount of p-nitrophenol released from p-nitrophenyl-β-D-glucopyranoside (pNPG) at 405 nm. A 0.2 mL solution of 5 mM pNPG was preincubated for 5 min in McIlvaine buffer pH 5.0 at 50°C. For this solution, 0.3 mL of the properly diluted enzyme sample was added. After 5 min of incubation, the enzymatic activity was stopped by adding 2 mL of 2 M Na2CO3 solution. One unit of enzyme activity was defined as the enzyme amount that releases 1 mM of p-nitrophenol per mL per minute of reaction. Specific activity was expressed as unit per mg of protein. All enzyme assays were performed at least three times and results are presented as mean values.
Protein concentrations were determined using the modified Bradford method with reference to a standard calibration curve for bovine serum albumin. 21
Culture Conditions for β-Glucosidase Production
Enzyme production on different agro-industrial wastes
Enzyme induction was studied with different substrates as sole carbon source, 1% (w/v) in Vogel's medium, incubated for 10 days, at 28°C under stationary conditions. The humid wastes were washed and dried in an oven at 60°C for 3 days. After the agro-industrial waste that induced the highest enzyme level was selected, some concentrations of this carbon source were evaluated from 0.5% to 3.5% (w/v).
Effect of culture conditions, initial pH, and temperature on β-glucosidase production
The influence of incubation period on β-glucosidase production was evaluated in standing culture for 12 days and under shaking culture (120 rpm) for 8 days. The effect of initial pH on the enzyme production was evaluated from 3.0 to 9.0, while the influence of temperature was verified from 20°C to 35°C.
Enzyme Characterization
Exclusion molecular chromatography
To verify the presence or absence of isoforms, the enzyme preparation was dialyzed against 50 mM ammonium acetate buffer (pH 6.8) and then lyophilized and dissolved in a small volume of this buffer. This sample was applied to a Sephadex G-100 column (Sigma-Aldrich, St. Louis, MO), equilibrated, and eluted with the same buffer flowing at 18 mL/h. Fractions (3 mL) were collected and tested for protein and β-glucosidase activity as previously described.
Optimal pH and temperature
The optimal temperature was determined by monitoring β-glucosidase activity at several temperatures from 35°C to 75°C, in McIlvaine buffer at pH 5.0. To determine the optimal pH, the β-glucosidase activity was assayed in different pH values using McIlvaine buffer from pH 3.0 to 7.0, at the optimal temperature.
β-glucosidase stability at different temperature and pH
Temperature stability was measured by incubating the enzyme at the predetermined optimal pH value for different periods, at temperatures ranging from 55°C to 70°C. Following incubation, the enzyme solution was frozen and the remaining activity determined. For pH stability assays, the crude filtrate was diluted (1:2 v/v) in McIlvaine buffer at a pH range 3.0 to 7.0. The samples were incubated at 4°C for 24 h. After this period, the β-glucosidase activity was assayed under optimal conditions and the residual activity was determined.
Effect of metals and substances
β-glucosidase activity was investigated against some metallic ions and other substances at final concentrations of 2 mM and 10 mM. The enzyme assay was performed at optimal conditions, and the relative activities were expressed as a percentage of the control. The data obtained were submitted to statistical analysis by Student's t-test at a significance level of 5% (p<0.05).
Effect of glucose on β-glucosidase activity
The extent of glucose inhibition was determined by incubating 10 μL enzyme preparation, 250 μL of 5 mM pNPG dissolved in glycine-HCl buffer at pH 2.5, and 240 μL of varying amounts of buffer and glucose with a final glucose concentration of 0–1,000 mM, at 65°C for 5 min. The Ki value was defined as the amount of glucose required to inhibit 50% of β-glucosidase activity and was calculated using GraFit software (Erithacus Software, Surrey, UK).
Results and Discussion
Influence of the Carbon Source and its Concentration on β-Glucosidase Production
The use of lignocellulosic wastes as carbon source in the growth medium would reduce the costs of enzyme production. β-Glucosidase production by P. miczynskii using different agro-industrial wastes is shown in Table 1. The carbon source that induced the highest enzymatic activity and fungal growth was pineapple peel, corresponding to 0.98 U/mL and 0.70 mg protein, respectively. To date, only two studies employed pineapple peel as substrate for microbial enzyme production. 22,23 However, these works did not evaluate β-glucosidase activity.
Influence of Agro-Industrial Wastes on β-Glucosidase Production by P. miczynskii
Average and standard deviation of two cultures.
ND, not detectable.
P. miczynskii was also able to produce β -glucosidase in the presence of apple peel and orange peel, yielding 0.27 U/mL and 0.25 U/mL, respectively. According to Matos and Reinhardt, the increase in fruit pulp consumption results in higher fruit processing wastes that can be processed into higher value-added products. 24 The use of pineapple peel to produce β-glucosidase has been shown to be promising in Brazil, since the country is a major pineapple producer. 16,24
In the presence of wheat bran, passion bagasse, and soybean peel, low levels of enzyme activity were verified, when compared to the cultures with pineapple peel. Enzyme activity was not detected when malt bagasse, sugar-cane bagasse, corn cob, rice peel, sugar-cane straw, or corn straw were used as carbon sources. Wastes such as wheat bran, sugar cane bagasse, rice husk, and maize straw have been shown to be appropriate for β-glucosidase production in other filamentous fungi. 25 –29 Gao et al., reported the use of corn stover in addition to wheat bran for β-glucosidase production by Fusarium proliferatum. 30 Differences in composition, as well differences in accessibility of the substrates by P. miczynskii, have resulted in distinct production levels among the evaluated agro-industrial residues. 30
The most efficient concentration of pineapple peel to induce P. miczynskii β-glucosidase production and fungal growth was 3% (1.99 U/mL) and 2.5% (1.04 mg protein), respectively (Table 2). Folakemi et al., verified that 5% of the same substrate induced high levels of cellulose production by Trichoderma longibrachiatum and Saccharomyces cereviseae. 22 On the other hand, Azzaz et al., reported the use of wheat straw at a concentration of 20% (w/v) to induce cellulose production by an Aspergillus niger strain (0.076 U/mL). 31
Effect of Pineapple Peel Concentration on β-Glucosidase Production by P. miczynskii
Average and standard deviation of two cultures.
Effects of Culture Conditions on β-Glucosidase Production
The capacity of microorganisms to produce enzymes is influenced by environmental conditions such as temperature, pH, agitation, and incubation period. For this reason, optimization of these parameters is important for developing the process. In standing culture, with pineapple peel as the carbon source, the highest extracellular β-glucosidase production, 1.07 U/mL, was obtained in 9-day-old cultures (Fig. 1a). The maximal fungal growth was observed on the 9th and 10th days, corresponding to 1.01 and 1.09 mg of protein, respectively (not shown). In shaking condition (Fig. 1b), the maximal activity in units per volume was achieved in 6-day-old cultures (0.68 U/mL). P. miczynskii showed maximum growth, measured by intracellular protein, on the 9th and 10th day in stationary condition and on the 5th and 6th day in shaking culture (not shown). According to Papagianni and Mattey, the mycelial morphology control in fermentation is often a prerequisite for industrial application. 32

Time-course of β-glucosidase production by P. miczynskii in
Macroscopic morphology examination of the P. miczynskii in stationary and shaking conditions revealed that, in the first case, the hyphae formed a freely dispersed mycelium, whereas in the second case, pellets were produced. According to Braun and Vecht-Lifshitz, for producing fungal metabolites the morphology varies from one product to another. 33 In some processes, free mycelia are required for increased productivities, as in the current work. 33,34 For this reason, the subsequent experiments were carried out under stationary condition.
Temperature and pH are directly related to growth behavior and development of the fungal mycelium, as well as to β-glucosidase production. The influence of pH on β-glucosidase production is shown in Fig. 2a. β-glucosidase production was verified in all initial pH values evaluated. The highest activity in units per volume was observed in two major peaks at initial pH from 4.5 to 5.5 and from 6.5 to 7.0. Dhake and Patil verified an optimum initial pH value of 5.5 for Penicillium purpurogenum β-glucosidase production, while Ang et al., reported high levels of Paecilomyces variotii β-glucosidase production within the pH range of 5.0 to 9.0. 35–36 P. miczynskii could grow in media with initial pH between 3.0 and 9.0, with maximal growth at 7.5 (0.73 mg of protein) and 8.5 (0.64 mg of protein; not shown). This result indicates the alkaline nature of this fungus.

Effect of initial ) specific β-glucosidase activity (U/mg of protein). Culture conditions: Vogel medium with 3% pineapple peel (w/v) under stationary condition for 9 days at
The effect of temperature on β-glucosidase production by P. miczynskii is presented in Fig. 2b. The maximal value of β-glucosidase production was verified at 20°C, corresponding to 2.82 U/mL, while the highest specific activity was obtained at 25°C, corresponding to 39.02 U/mg of protein. However, other authors determined the best temperature range for filamentous fungi β-glucosidase production between 30°C and 45°C. 22,37 The highest growth was verified at 20°C (0.42 mg of protein), while at 35°C P. miczynskii growth was not detected (not shown). The optimal temperature for β-glucosidase production was similar to the optimal temperature for the growth of the fungus, corresponding to the environmental temperature from which it was initially isolated. This observation was in accordance with those reported by Knob and Carmona studying β-xylosidase production by Penicillium sclerotiorum. 38
Biochemical Properties of P. Miczynskii β-Glucosidase
Chromatographic analysis on Sephadex G-100 indicated the presence of only one form of β-glucosidase (data not shown). β-glucosidase from P. miczynskii showed maximal activity at 65°C (Fig. 3a). Similar β-glucosidases with growth at optimal temperatures from 50°C to 65°C have been reported from several fungi, such as Penicillium purpurogenum and Aspergillus sp. 35,39 Besides, other fungal β-glucosidases presented maximal temperature activity at 70°C. 40,41 This study showed that the best pH for β-glucosidase activity was in the range of pH 4.5–5.0 (Fig. 3b), which is similar to that reported for the β-glucosidase from Diaporthe (Phomopsis) helianthi, Aspergillus awamori, and T. aurantiacus. 28,42,43

Influence of
The stability of enzymes remains a critical aspect for biotechnological applications. For this reason, thermal and pH stability of β-glucosidase produced by P. miczynskii were investigated. The enzymatic preparation was incubated without substrate at 55°C, 60°C, 65°C, and 70°C (Fig. 4a). P. miczynskii β-glucosidase was stable at 55°C and 60°C, showing half-life (T1/2) values of 50 min and 40 min, respectively. P. miczynskii β-glucosidase was more stable than that produced by Aspergillus awamori, which retained 60% of its activity after 10 min of incubation at 55°C. 43 Conversely, Chodi et al., verified a β-glucosidase from Humicola brevis that was stable at 65°C, with a T1/2 of 5.1 hours. 44
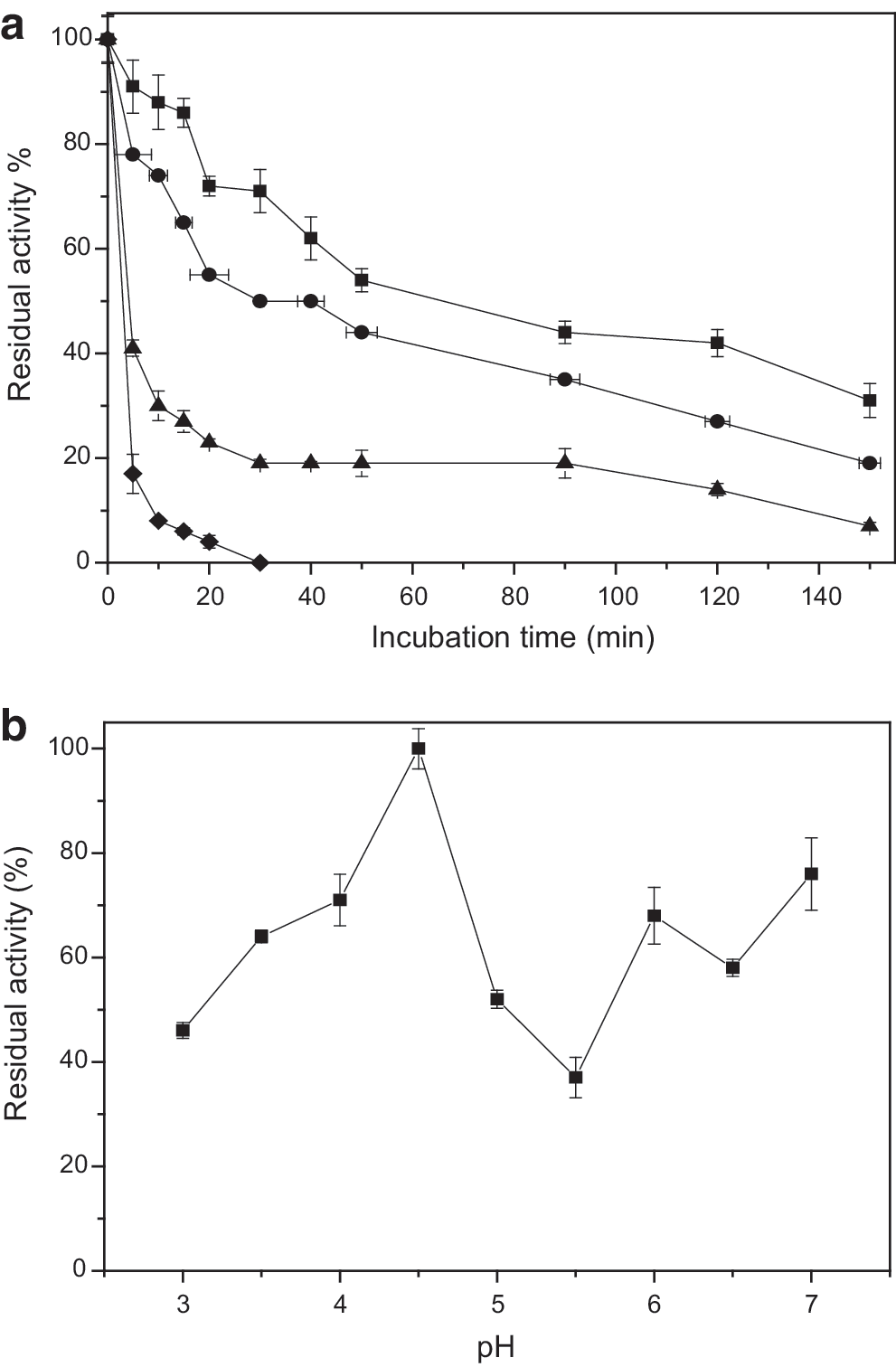
The β-glucosidase produced by P. miczynskii maintained its stability in acid and neutral conditions (Fig. 4b). High stability (around 80%) was observed at pH 7.0 and 4.5, while at pH 3.5, 4.0, 6.0, and 6.5 more than 60% of its residual activity was verified. Low residual activity (less than 40%) was observed at pH 3.0 and 5.0. Aspergillus fumigatus β-glucosidase remained stable at pH ranging from 4.0 to 7.0, while β-glucosidase from Melanocarpus sp. exhibited pH stability between 5.0 and 6.0. 45,46
To verify the effect of substances on β-glucosidase activity, the crude filtrate was incubated in the presence of several metallic ions, sodium dodecylsufate (SDS), tetrasodium ethylenediaminetetraacetate (EDTA), and β-mercaptoethanol, at 2 mM and 10 mM concentrations (Table 3). The chelating agent EDTA inhibited β-glucosidase activity, indicating that this enzyme is a metalloprotein and requires metal ions for action. Hg2+, Mg2+, and Zn2+ were strong inhibitors of β-glucosidase activity, while sodium citrate, Cu2+, Mn2+, NH4 +, and Ba2+ had a moderate inhibitory effect on the enzyme. Likewise, Humicola grisea, Aspergillus oryzeae, and Monascus purpureus β-glucosidase were inhibited by some of these elements. 47 –49 The inhibition by Hg2+ indicates the presence of thiol groups of cysteine residues in β-glucosidase active sites or around them. β-Glucosidase activity retained less than 50% of its initial activity in the presence of SDS, indicating the relevance of hydrophobic interactions to maintenance of native structure.
Effect of Different Substances on β-glucosidase Production by P. miczynskii
β-Glucosidase showed enhanced activity in the presence of the reducing agent β-mercaptoethanol, which can be explained through its ability to prevent the oxidation of sulfidryl groups. The observed slight activation by Ca2+ and Co2+ may be explained by the ions' effect on enzyme structure stabilization. Similarly, Daldinia eschscholzii β-glucosidase was activated in the presence of Ca2+ and Co2+. 8
Glucose is an end-product inhibitor of β-glucosidases. 50 For practical purposes, it is essential that β-glucosidase be glucose tolerant in order to make enzymatic saccharification of cellulolytic substrates an efficient process. So, to determine the extent of glucose inhibition, the enzyme preparation was incubated with pNPG, at varying glucose concentrations (Fig. 5). The enzyme was inhibited with a Ki of 760 mM. The majority of β-glucosidases reported to date present Ki values for glucose ranging from 0.2 mM to no more than 100 mM. 7,8,51 –53 However, some fungal β-glucosidases show high glucose tolerance with Ki values of more than 100 mM. 48 Recently, Karnaouri et al., purified a glucose tolerant β-glucosidase from Myceliophthora thermophila, that had a Ki of 282 mM. 54 The Thermoanaerobacterium thermosaccharolyticum β-glucosidase is also a glucose-tolerant enzyme, with a Ki of 600 mM. 12
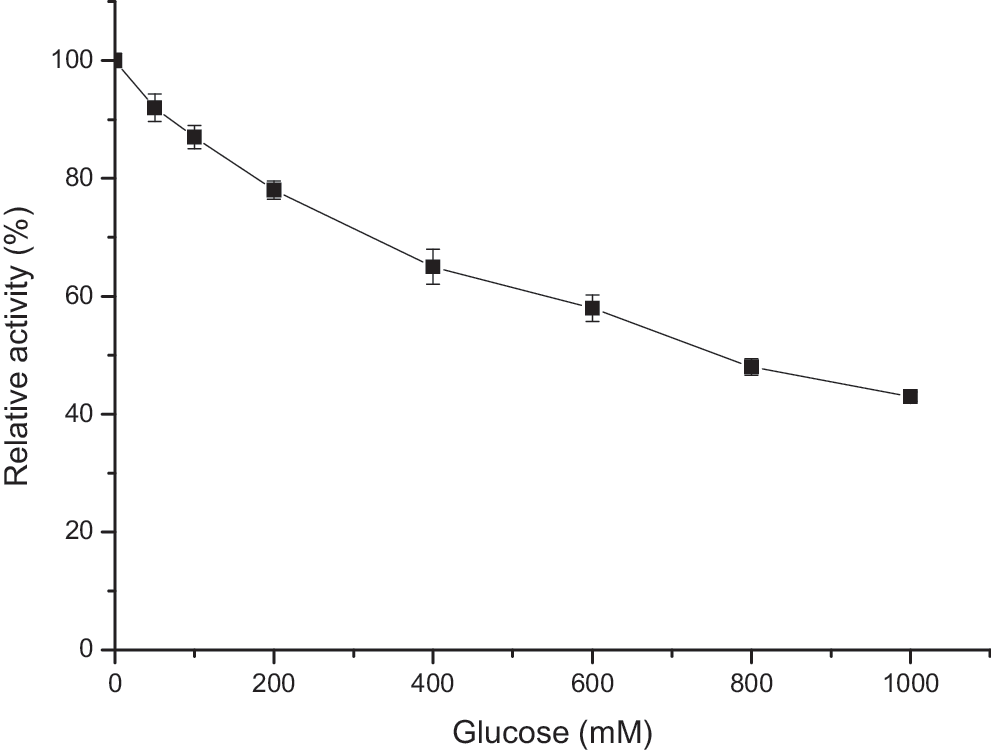
Inhibition of β-glucosidase activity by glucose. The enzyme preparation was incubated with pNPG, varying glucose concentrations (0–1000 mM), at 65°C for 5 min. The data are presented as means for duplicate measurements.
Conclusions
In this study, a P. miczynskii strain was able to produce high levels of β-glucosidase using pineapple peel as substrate. Large amounts of this agro-industrial waste—a by-product of pulp industries—are generated and accumulated annually. The use of pineapple peel for β-glucosidase production can decrease the impact of deposition of this waste on the environment and add to the value of the residual by-product. Additionally, β-glucosidase production cost can be reduced. A new β-glucosidase produced by P. miczynskii was able to function in high temperatures and at acidic pH, and it was stable in a wide pH range. These characteristics and the low inhibition rate by glucose allow its application in a wide field of biotechnological processes such as additives in cellulose-based feeds; release of aromatic compounds from glycosidic precursors present in fruit juices, musts, and wines; and bioethanol production.
Footnotes
Acknowledgments
The authors acknowledge the Coordination for the Improvement of Higher Level Personnel (CAPES-Brazil) for the scholarship awarded to SM Beitel.
Author Disclosure Statement
No competing financial interests exist.