
Abstract
D-ribose is a commercially important functional sugar used as a nutritional supplement and as a starting compound in the synthesis of antiviral drugs. In this study, we report engineered Escherichia coli mutants that can produce D-ribose from glucose and xylose. Two endogenous haloacid dehalogenase-like (HAD) phosphatases from E. coli, HAD12 and HAD13, encoded by the genes ybiV and yidA, respectively, were expressed in E. coli wild type and the glucose-xylose co-utilizing mutant LMSE2. All the mutants constructed in this study produced D-ribose. The mutant RB-006 (LMSE2 expressing ybiV) showed the highest D-ribose titer of 1.16 g/L from 5 g/L each of glucose and xylose. Additionally, using xylose feeding, D-ribose titer was improved to 3.36 g/L. Xylulose and acetate were formed as the major byproducts in the fed-batch study. This study represents the first example of engineered E. coli for production of D-ribose. This study also demonstrates reengineering of a glucose-xylose co-utilizing mutant of E. coli for production of a valuable chemical.
Introduction
D-ribose is a naturally produced pentose sugar with several commercial applications. It is used as a sweetener, as well as a starting compound for synthesis of riboflavin, inosine monophosphate, and several antiviral drugs. 1,2 D-ribose is also used as a nutritional supplement for muscle recovery among athletes, and aids in functional recovery of patients with ischemic cardiovascular disease. 3 D-ribose can potentially be used as a starting compound for production of its stereoisomer L-ribose, which has a market price of about $1,000/kg as compared to $20–30 /kg for D-ribose. 4
The earliest methods of synthesizing D-ribose involved enzymatic hydrolysis of yeast RNA or chemical synthesis from glucose, arabinose, gluconic acid, and xylose. 5 However, these methods suffered from poor conversion yields and thus did not achieve much commercial success. Fermentation-based processes were then developed for replacing the chemical- and biochemical-based methods for commercial production of D-ribose. 1 Several organisms, including Penicillium brevicompactum, Pseudomonas reptilivora, and Candida pelliculose, are known to produce D-ribose naturally. However, due to low yields and the difficulties in culturing these microorganisms, their use in fermentation processes has found limited applications.
Current industrial fermentation processes for D-ribose production employ transketolase-deficient Bacillus spp. isolated using random mutagenesis. 1 Absence of transketolase, and/or other pentose phosphate pathway (PPP) enzyme D-ribulose-5-phosphate-3-epimerase, results in the accumulation of ribose-5-phosphate (R5P), which is converted to D-ribose by the endogenous phosphatases in Bacillus spp. Similar transketolase mutants of other industrially relevant organisms such as E. coli and Corynebacterium glutamicum do not show accumulation of D-ribose despite supplementation of aromatic amino acids and vitamins (pyridoxine). 6,7 A Saccharomyces cerevisiae strain with deletion of transketolase (TKL1) and phosphoglucose isomerase (PGI1), over-expressing the genes GHD2 and DOG1, was recently shown to produce D-ribose and ribitol. 2 The most preferred Bacillus strain for commercial production of D-ribose is B. subtilis. 1,8 Many different variants of B. subtilis with transketolase deletion have been reported to produce D-ribose from glucose and other carbon sources at high yields and titers. A summary of D-ribose-producing strains with their culture conditions is presented in Table 1. 1,2,5,8,9
D-Ribose-Producing Strains Previously Described in Literature
Feed: 100 g/L xyl.
Feed: 200 g/L xyl+50 g/L glu.
YNB, yeast nitrogen base.
Includes concentration of both D-ribose and ribitol.
Apart from Bacillus spp., E. coli is a strong candidate for D-ribose production, especially from glucose-xylose mixtures obtained from lignocellulosic biomass. E. coli has been extensively studied for growth on lignocellulosic hydrolysates as feedstocks. 10 –12 Additionally, there exists a possibility for further engineering of D-ribose-producing E. coli for production of more valuable compounds such as L-ribose. Previously, we reported metabolic engineering of E. coli for co-utilization of glucose and xylose. 13 The E. coli mutant LMSE2, which contains three metabolic gene deletions—pgi, rpe, and eda—is capable of strictly co-utilizing glucose and xylose. 13 Due to the gene deletions, the glucose flux in the mutant is channeled directly into the PPP, converting glucose directly to R5P, whereas xylose is exclusively converted to xylulose-5-phosphate (X5P). Using R5P and X5P, the mutant LMSE2 synthesizes all the biomass precursors using the PPP. A metabolic map of the mutant LMSE2 is shown in Fig. 1a.
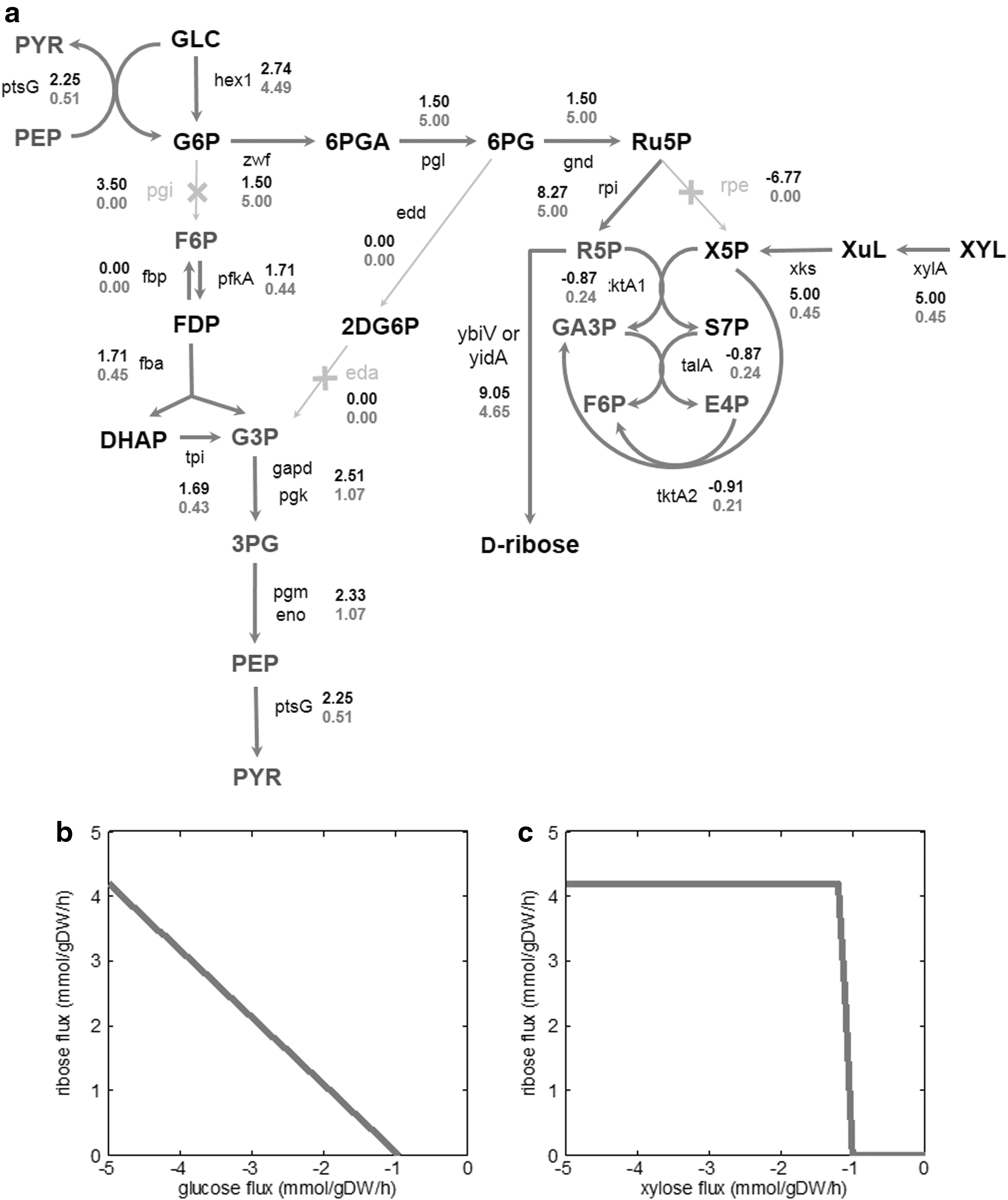
Metabolic network modifications used for production of D-ribose.
As glucose is exclusively converted to R5P in the mutant LMSE2, over-expression of a phosphatase capable of cleaving the phosphate bond in R5P is expected to result in production of D-ribose. Additionally, as the mutant LMSE2 can consume xylose simultaneously with glucose, xylose is expected to be utilized for synthesis of biomass precursors, thereby increasing the yield of D-ribose per mole of glucose consumed.
With the aim of producing D-ribose using the mutant LMSE2, we selected two endogenous phosphatases that could cleave the phosphate bond in R5P. A family of 23 haloacid dehalogenase (HAD)-like phosphatases in E. coli was previously screened against 80 different phosphorylated substrates. 14 Among these 23 enzymes, four HADs were active towards dephosphorylation of R5P. Two phosphatases, HAD12 (encoded by ybiV) and HAD13 (encoded by yidA), were chosen to be expressed in the mutant LMSE2, based on their kinetic properties. 14 Multiple mutants were constructed by transforming E. coli wild type and the mutant LMSE2 with the genes ybiV and yidA on an appropriate vector. The mutants thus constructed were characterized for production of D-ribose from a mixture of glucose and xylose.
Materials and Methods
FBA and Robustness Analysis
Flux distributions for D-ribose production for the wild-type and the mutant LMSE2 were obtained using flux balance analysis (FBA). All the simulations were carried out using the genome-scale model iJO1366. 15 Flux values were obtained by simulating maximization of D-ribose production. The glucose and xylose exchange fluxes were fixed at −5 mmol/gDW/h each, and the minimum biomass production was fixed at 0.1/h. To obtain a valid wild-type flux distribution, additional constraints were added to divert 30% of total glucose uptake flux into the PPP. To simulate the LMSE2 mutant phenotype, six reactions (PGI, RPE, EDA, XYLI2, PPS, and DRPA) were deleted from the model. 13 All the simulations were carried out in MATLAB environment using COBRA Toolbox v.2. 16
Robustness analysis was carried out for D-ribose production against glucose uptake and D-ribose production using xylose uptake in the LMSE2 background. The minimum biomass production was constrained to 0.1/h. The analysis was carried out using the in-built function in COBRA Toolbox v.2.
Strains and Plasmid
All the strains used in this study were obtained from the Coli Genetic Stock Center at Yale University (New Haven, CT). E. coli MG1655 was used as the wild type strain. The mutant LMSE2 (E. coli Δpgi Δeda Δrpe::kanR) was constructed using sequential P1 transduction and engineered further by adding genes, either ybiV or yidA, expressing phosphatases HAD12 and HAD13, respectively. Both the genes were amplified via polymerase chain reaction using forward and reverse primers with restriction enzyme sites for KpnI and HindIII, respectively. The bold regions of the primers represent the restriction enzyme sites:
yidA KpnI forward: AGTCATT
yidA HindIII reverse: GGATCGT
ybiV KpnI forward: AGTCATT
ybiV HindIII reverse: GGATCGT
The amplified genes were ligated into the plasmid pTrc99A, and the plasmids were transformed into the mutant LMSE2. Appropriate controls were constructed by transforming E. coli wild type with the same plasmid constructs, and also by transforming the mutant LMSE2 and the wild-type with the empty plasmid pTrc99A. All the constructed mutants with their genotypes and designations are listed in Table 2.
Phenotypes of the Mutants Constructed in this Study
MM, minimal medium; YE, yeast extract (0.5 g/L added); LB, Luria-Bertani broth (0.5 g/L added); G, growth R, D-ribose.
Phenotypic Tests
All six constructed mutants were first tested for growth and D-ribose production using phenotypic characterization. All the mutants were grown on Luria-Bertani (LB) plates supplemented with 5 g/L glucose, 5 g/L xylose, and 100 mg/L ampicillin. Individual colonies were grown overnight in 3-mL LB medium supplemented with glucose (5 g/L), xylose (5 g/L), and ampicillin (100 mg/L). The cells grown overnight were washed in minimal medium without sugars to remove all the LB and were used to inoculate three different media: minimal medium; minimal medium supplemented with 0.5-g/L yeast extract; and minimal medium supplemented with 0.5-g/L LB. Additionally, each tube was supplemented with 1 mM of isopropyl β-D-1-thiogalactopyranoside (IPTG) to induce the plasmid. Mutants constructed from the wild type (RB-001, RB-002, and RB-003) were checked for growth and production of D-ribose after 12 hours of incubation, and the mutants constructed from LMSE2 (RB-004, RB-005, and RB-006) were checked for growth and D-ribose production after 24 hours of incubation. Growth in the tubes was measured as optical density at 550 nm (OD550), and D-ribose was detected using high performance liquid chromatography (HPLC, method described below).
Media and Batch Cultivation
For batch fermentation, the constructed mutants were cultivated in 500–mL fermenters (Applikon MiniBio; Applikon Biotechnology, Delft, The Netherlands), with 300 mL working volumes. The composition of minimal medium used for cultivation of the mutants was (per L): 3.5 g KH2PO4, 5 g K2HPO4, 3.5 g (NH4)2HPO4, 0.25 g MgSO4.7H2O, 15 mg CaCl2.2H2O, 0.5 mg thiamine, and 1 mL of trace metal stock. The trace metal stock was prepared in 0.1 M HCl and contained (per L): 1.6 g FeCl3, 0.2 g CoCl2.6H2O, 0.1 g CuCl2, 0.2 g ZnCl2.4H2O, 0.2 g NaMoO4, 0.5 g H3BO3. For pH control, 3-N-morpholinopropanesulfonic acid (0.1 M, pH 7.2, filter-sterilized) was added to the medium. 17 Glucose and xylose were used in a concentration of 5 g/L each. Additional supplementation of yeast extract (0.5 g/L) was added to the medium to enable growth of the LMSE2-based mutants. Additionally, ampicillin (100 mg/L was added to the medium to maintain the selection pressure on the plasmid). The plasmid was induced using 1 mM IPTG when the culture reached an OD550 of 1.0. Fermenters were sampled every hour or every three hours, depending on the growth rates of the mutants. The batch was kept aerobic (greater than 30% dissolved oxygen) by altering the revolutions per minute (RPM) of the impeller. The pH of the medium was maintained at 7.00 using 5 M KOH.
The inoculum was prepared by first inoculating a fresh colony in LB with 5 g/L glucose and 5 g/L xylose. The cells grown overnight were first washed and then transferred to minimal medium with 5 g/L glucose and 5 g/L xylose and grown until OD550 was between 1.5–2.0. The cells were then harvested and an appropriate amount was used to inoculate the fermenters to get an initial OD550 of 0.1. Each sample was analyzed for cell density (OD550), residual sugars, D-ribose, and coproducts (acetate, xylulose, erythrose, and fructose). The sugars were separated using a Bio-Rad (Hercules, CA) HPX-87H cation-exchange column (5 mM H2SO4 mobile phase, 0.4-mL/min flow rate, 42°C column temperature, 20 μL injection volume).
FED-Batch Cultivation
For the fed-batch culture, an appropriate volume of 300 g/L xylose was added to the fermenters as an impulse-feed after all the xylose was consumed. The amount of feed added was chosen based on the residual concentration of glucose in the fermenter, and was such that glucose and xylose concentrations in the fermenter were equal after the feed addition. Due to the very high concentration of the xylose feed, the dilution effect in the fermenter was negligible. The impulse feed volume was approximately 3 mL. The samples were withdrawn every 3 h until the end of the batch, indicated by rising pH and dissolved oxygen levels.
Results
FBA and Robustness Analysis
Predicted flux distributions for production of D-ribose in the wild-type and the mutant LMSE2 are shown in Fig. 1a. The flux values of the wild-type show the maximum possible theoretical yield of D-ribose production. However, such high yields are achievable only if most of the carbon flux in the wild-type is diverted towards D-ribose production and growth rate is as low as 0.1/h. This flux distribution is very unlikely in the wild type as the majority of the carbon is diverted towards biomass formation. To achieve the FBA-predicted yields in the wild type, major rerouting of the central metabolism fluxes (as shown in Fig. 1a) is required.
The flux distribution of the mutant LMSE2 shows that the majority of D-ribose in the mutant is produced from glucose being diverted into the PPP. The mutant LMSE2 has three gene deletions that cause rerouting of the fluxes, and most of the glucose is diverted into the PPP. The FBA-predicted fluxes are hence very plausible, and the mutant is expected to show high D-ribose yields on glucose.
Robustness analyses of D-ribose production with respect to glucose-exchange flux (Fig. 1b) and xylose-exchange flux (Fig. 1c) in the mutant LMSE2 show that D-ribose is almost exclusively produced from glucose. Decreasing the glucose-uptake flux proportionally decreases the D-ribose-production flux, whereas xylose-exchange flux has no effect on D-ribose production. These results suggest that xylose is used by the mutant LMSE2 only for making biomass precursors.
Phenotypic Characterization
Phenotypic characterization of the constructed mutants was carried out as a first test to detect D-ribose production in E. coli. The results for the phenotypic characterization are summarized in Table 2. The control mutants (RB-002 and RB-003) showed production of D-ribose under all three media conditions, whereas no D-ribose was detected in the control expressing the empty plasmid (RB-001). The mutants based on LMSE2 (RB-005 and RB-006) did not show growth on minimal medium, but showed growth and D-ribose production on minimal medium supplemented with yeast extract and LB. The LMSE2 mutant transformed with empty plasmid did not show any D-ribose production.
The mutants RB-005 and RB-006 did not show growth on minimal medium most likely due to the high activity of phosphatase on R5P resulting in low intracellular concentrations of R5P, which precluded the production of biomass precursors such as erythrose-4-phosphate (E4P) through the PPP. Though over-expression of phoshpatase could have been depleting the pools of R5P in the wild-type-based control strains RB-002 and RB-003 as well, the phosphatase expression was expected to be more deleterious for the mutants RB-005 and RB-006. This is mainly because the wild-type-based strains could use redundant pathways to make phosphorylated metabolites such as fructose-6-phosphate and glyceraldehyde-3-phosphate (G3P), which could be used to synthesize other biomass precursors. For example, RB-002 and RB-003 could synthesize G3P using either the Embden-Meyerhof-Parnas or Entner-Doudoroff pathways, or the PPP. In contrast, the mutants RB-005 and RB-006 had to synthesize all biomass precursors using the PPP, which was disrupted because of low concentrations of R5P. Such limitations on metabolism could have caused the expression of phosphatase to be more lethal for the mutants RB-005 and RB-006 than the control strains RB-002 and RB-003.
The addition of yeast extract and LB at a low concentration of 0.5 g/L most likely facilitated the growth of the mutants RB-005 and RB-006 by supplying amino acids that these mutants otherwise could not synthesize in the presence of phosphatases. Based on the results from the phenotypic studies, yeast extract was added to the subsequent fermenter cultivations. In addition, to enable the initial growth, IPTG was added to the fermenters only after OD550 had reached a value greater than 1.0.
Batch Studies
Four D-ribose-producing mutants—RB-002, RB-003, RB-005, and RB-006—were characterized in detail using batch-cultivation studies. The cells were cultivated in minimal medium with yeast extract as a supplement. Figure 2 shows the growth and D-ribose production characteristics of these mutants. All four mutants characterized produced D-ribose from the mixture of glucose and xylose. The mutant RB-006 accumulated the highest concentration (1.16 g/L) of D-ribose. The most likely source of D-ribose in the mutant RB-006 (and RB-005) was glucose, as predicted by the simulations (Fig. 1b

Growth plots of RB-series mutants; acetate concentration.
An important difference between the wild-type and the mutant LMSE2 is the sugar-consumption pattern. Whereas the wild type shows sequential utilization of sugars, the mutant LMSE2 has been engineered to co-utilize glucose and xylose. 13 Accordingly, the mutants based on the wild type, RB-002 and RB-003, consumed glucose and xylose sequentially (Figs. 2a and b); however, the mutants RB-005 and RB-006, which were constructed in the mutant LMSE2 background, consumed xylose preferentially over glucose, while consuming some glucose simultaneously with xylose. Even though the mutant LMSE2 is designed to show strict co-utilization of glucose and xylose, the most likely reason for lower consumption of glucose and higher dependence on xylose was the additional supplementation of yeast extract. 13 As the mutants most likely synthesized some of the biomass precursors from yeast extract, the mutants did not need to utilize high amounts of glucose. Based on these results, the concentration of yeast extract was reduced to 0.2 g/L in the subsequent fed-batch experiments to force higher uptake of glucose.
The mutants RB-003 and RB-005, both expressing the gene ybiV encoding HAD12, produced more D-ribose compared to the corresponding mutants RB-002 and RB-004, which expressed the gene yidA encoding HAD13. This difference in D-ribose production was most likely due to the differences in substrate specificity of the enzymes HAD12 and HAD13. HAD13 has a higher promiscuity towards phosphorylated substrates, and acts on diverse substrates such as E4P, glucose-1-phoshpate, mannose-1-phosphate, fructose-1-phosphate, and glucose-6-phosphate (in addition to R5P). In contrast, the alternative substrate range for HAD12 is limited to fructose-1-phosphate, glucose-6-phosphate and acetyl phosphate. 14 Over-expression of an enzyme as promiscuous as HAD13 could have caused general dephosphorylation of multiple intermediates in the cell. Higher promiscuity of HAD13 could also have been the reason for reduced growth of the mutant RB-005 compared to the mutant RB-006.
Since the enzymes HAD12 and HAD13 were promiscuous, multiple coproducts could have been produced by the RB-series mutants. The fermentation samples were thus tested for production of fructose and erythrose, although neither was detected in any of the fermentation samples. The only coproducts detected in the batch cultivations were acetic acid and low amounts of xylulose. The results of the batch cultivation of the RB-series mutants are summarized in Table 3. The best-performing mutants were RB-003 and RB-006. As anticipated, the yield of the LMSE2-based mutant RB-006 was higher than the wild-type-based mutant RB-003, and this was most likely due to direct channeling of glucose into D-ribose production. The concentration of D-ribose produced by the mutant RB-006 was also higher than the mutant RB-003. One aspect in which the mutant RB-003 was better than the mutant RB-006 was productivity, and this was due to higher growth and sugar consumption rates of RB-003 compared to RB-006.
D-Ribose Production Characteristics by the RB-series Mutants a
Error values are standard deviations of three batches.
Table 4 shows the carbon balance for the mutant RB-003 and the mutant RB-006. 18,19 The mutant RB-003 consumed both glucose and xylose and produced mainly biomass and carbon dioxide (CO2). Some carbon was also lost due to overflow metabolism and was converted to acetate. On the other hand, the mutant RB-006 did not consume all the glucose and produced less biomass and less acetate. The overflow metabolism in the mutant was slower due to a lower glycolysis rate (Fig. 1a). The results indicate that although the wild-type-based mutant RB-003 is capable of producing D-ribose using expression of phosphatase, the mutant tends to waste a large fraction of carbon consumed as biomass, CO2, and the byproduct acetate. Although the LMSE2-based mutant RB-006 grows more slowly and consumes sugars at a lower rate, it is capable of diverting more carbon consumed toward D-ribose production.
Carbon Balance for the Mutants RB-003 and RB-006
The yeast extract was assumed to be 70% protein, and the carbon content of the protein was assumed to be 0.53 g/g. 18
The carbon content of the E. coli biomass was assumed to be 47% on w/w (dry cell). 19
Assuming that the unaccounted carbon was lost as CO2.
Carbon lost as CO2 is not included in this sum of total carbon.
FED-Batch Studies
The results obtained from the fed-batch cultivation studies are shown in Fig. 3. Due to the preferential consumption of xylose by the mutant RB-006, xylose was exhausted in the fermenter faster than glucose. For about every 5 g/L of xylose consumed, only 2.5 g/L of glucose was consumed. During this stage of consumption, approximately 2.5 g/L of D-ribose was synthesized. However, as RB-006 could not consume glucose alone, additional xylose was added to the fermenter to facilitate the consumption of the remaining glucose and possibly increase D-ribose titers. After addition of the feed, the concentrations of glucose and xylose were approximately 4 g/L each. With additional xylose fed into the fermenter, more glucose was consumed, and the D-ribose titer increased to 3.36 g/L.

Fed-batch fermentation with the mutant RB-006. The symbol • represents biomass; ♦ glucose concentration; ▲ xylose concentration; ■ D-ribose concentration; acetate concentration; and  xylulose concentration.
xylulose concentration.
In addition to D-ribose, most of the xylose added was converted into the byproduct xylulose (final concentration 5.4 g/L) and some acetate. These results show that the mutant RB-006 could not consume glucose even in presence of xylose and stopped growing as well as producing D-ribose. The most likely reason behind the poor glucose uptake and the poor conversion of xylose to D-ribose could be the fact that utilization of both glucose and xylose require energy. Glucose is phosphorylated during its transport inside the cell, and xylose enters PPP as xylulose-5-phosphate after being phosphorylated at the cost of an adenosine triphosphate (ATP). As the glycolytic flux in the mutant RB-006 is low, the insufficient ATP production most likely stalled the growth and uptake of the sugars.
Discussion
Unlike transketolase-deficient Bacillus mutants, transketolase-deficient E. coli mutants do not show accumulation of D-ribose. 6 Causes for the absence of D-ribose production in the transketolase-deficient E. coli could be either insufficient activity of the endogenous phosphatases or low intracellular concentration of R5P. We achieved production of D-ribose in E. coli by addressing both of these concerns. Production of D-ribose was conceived in the mutant LMSE2 based on the fact that glucose consumed by the mutant LMSE2 is channeled directly into PPP, resulting in its exclusive conversion to R5P. 13 Additionally, it was presumed that R5P produced by the mutant LMSE2 could be dephosphorylated by over-expression of an endogenous phosphatase.
Over-expression of phosphatases HAD12 or HAD13 showed D-ribose production in both wild-type and the mutant-LMSE2-based strains, RB-003 and RB-006, respectively. However, in RB-003, most of the carbon obtained from the sugars was converted into CO2 and biomass, resulting in a decreased overall yield of D-ribose. In RB-006, glucose was converted almost exclusively into D-ribose, resulting in higher yields. Another advantage of using RB-006 was the fact that the mutant could co-utilize glucose and xylose, a very important trait in industrial fermentations. 20,21 This glucose-xylose co-utilization phenotype is especially important in fed-batch fermentations, in which a mixture of glucose and xylose can be fed directly without the complication of accumulation of one sugar.
Despite the advantages, the mutant RB-006 posed certain limitations in D-ribose production, such as limited conversion of glucose-xylose mixture to D-ribose, lower sugar-uptake and growth rate, and dependence on yeast extract for growth. Results obtained from the fed-batch characterization show that the xylose feed was almost exclusively converted to xylulose, and glucose was not completely consumed. The reason for limited conversion of glucose and xylose to D-ribose and lower growth rate was most likely limited ATP production by the mutant. ATP is required for phosphorylation of both glucose and xylose as the first step of catabolic pathways, and is also required for biomass production. Low ATP production in RB-006 was due to the metabolic make-up of the mutant, which supported low glycolytic flux (Fig. 1a). Finally, the dependence on yeast extract for growth was due to high phosphatase activity causing depletion of R5P, resulting in no flux through the PPP for production of biomass precursors.
All of these issues associated with RB-006 are addressable and can be solved by additional engineering of the mutant and/or a follow-up treatment such as adaptive evolution. Tight control over the expression of the phosphatase gene would solve multiple problems associated with the mutant. Low ATP production as well as limited production of biomass precursors in the mutant are mainly due to limited flux through the PPP. High phosphatase activity leading to dephosphorylation of R5P depletes the R5P pools, stalling the flux through the PPP. Controlled activity of the phosphatase gene, allowing sufficient intracellular pools of R5P to drive the PPP, can ensure ATP production to allow continuous glucose and xylose uptake and production of biomass precursors to reduce the dependence on yeast extract. Controlled expression of the phosphatase gene can be easily achieved using tightly regulated systems used to express toxic proteins in E. coli. 22 The mutant LMSE2 is capable of growing without yeast extract. 13 If the phosphatase gene is completely repressed during the growth phase of the mutant RB-006, it would not require yeast extract supplement, as all the biomass precursors would be provided from co-utilization of glucose and xylose. To eliminate the dependence on yeast extract and to improve the sugar uptake rates, adaptive evolution of the mutant can also be carried out.
For the E. coli mutants described in this study to compete with the industrial Bacillus strains, multiple bioprocess considerations must also be investigated. One of the important characteristics of industrial strains is high sugar tolerance. In E. coli strains, sugar tolerance is strain-dependent. 23 –25 The mutants in this study were constructed in K-12 MG1655 background, which is a common industrial strain of E. coli and is known to grow at high glucose concentrations of 40 g/L. 23 Another consideration in industrial fermentations is the dissolved oxygen concentrations in the fermenters. Wild-type E. coli can convert up to 50% carbon to CO2, thereby losing a significant amount of carbon under aerobic conditions. 26 Anaerobic and microaerobic conditions can potentially increase product yields by conserving the carbon lost as CO2. 26 The current study characterized the mutants under high dissolved oxygen conditions in the fermenters (>30%). Cultivation under reduced oxygen (microaerobic) conditions is expected to increase the yields of D-ribose by minimizing the loss as CO2. Optimum time for induction can also critically affect the productivities of the target compounds. The biomass produced by the mutant RB-006 was lower compared to the wild-type-based mutant RB-003, which increased the batch time and decreased the productivity of D-ribose. Optimum induction time of the plasmid can further improve the productivity of D-ribose by allowing sufficient biomass formation required to achieve higher sugar consumption rates and higher production rates.
In conclusion, we demonstrate the construction of E. coli mutants capable of producing D-ribose from mixtures of glucose and xylose. Using over-expression of endogenous phosphatase HAD12, and feeding xylose, we were able to achieve higher titers and yields of D-ribose. Though high concentrations of D-ribose were achieved using the LMSE2-based mutant RB-006, a significant amount of xylose was converted to xylulose, indicating the need for additional metabolic engineering and bioprocess optimization to achieve industrially relevant titers.
Footnotes
Acknowledgments
The authors thank NSERC Bioconversion Network (Guelph, Canada) for funding the research and Professor Vincent Martin, Concordia University (Montreal, QC) for providing plasmid pTrc99A.
Author Disclosure Statement
No competing financial interests exist.