
Abstract
Bioethanol is currently one of the most important sources of renewable fuel and has the potential to partially replace petroleum-based liquid fuels. The United States is the largest ethanol producer in the world, with an average production of 54 billion liters per year. Most of its production derives from cornstarch. The Carbon Intensity Index (CI) for cornstarch-derived ethanol—75.97 g of carbon dioxide equivalent (CO2e) emitted per megajoule (MJ) of energy produced—is close to the gasoline index (95.86 g of CO2e/MJ), and the use of cornstarch-derived ethanol has little impact on the emission of greenhouse gases. Compared to ethanol from cornstarch, production using sugarcane has a lower CI (56.66 g of CO2e/MJ), and greater emission reductions can be achieved by replacing gasoline or other fuels with ethanol obtained from sugarcane. Brazil, the second largest world producer of ethanol, produces about 25 billion L/y of ethanol and has the largest flex-fuel vehicle fleet in the world. Nevertheless, the Brazilian ethanol sector is in a state of economic stress caused mainly by the high operational costs and the fact that sugarcane productivity seems to have reached a plateau. In this scenario, conversion of the sugars present in lignocellulosic biomass into ethanol emerges as a promising technology with the potential to revolutionize the renewable fuels sector. In this review, we present the advances and challenges in producing and fermenting lignocellulosic sugars, demonstrating the huge potential that this technology represents for the renewable fuels sector.
Introduction
The notion that oil, one of the most ubiquitously used natural resources in current society, could be depleted began with the oil crisis in the 1970s. 1 The recent discovery of new oil reserves with high potential productivity has replaced the uncertainty regarding oil availability with economic, geopolitical, and environmental problems related to oil use. 1 The cracking processes used to produce many petrochemical commodities and the use of oil as a fuel both lead to the release of a huge amount of carbon dioxide (CO2) into the atmosphere–CO2 that was previously entrapped in the soil as hydrocarbons. The CO2 produced by the combustion of fossil fuels is a major source of greenhouse gas emissions. 2 Moreover, the concentration of CO2 in the atmosphere has increased by approximately 90 ppm in the last 50 years, representing an almost 30% increase in atmospheric CO2 levels since the 1960s. 3 Many studies analyzing the increase in the CO2 level have suggested that such changes can result in disastrous consequences for the environment. 4,5 The importance of this issue has been demonstrated by the numerous studies of the potential environmental impact of global warming resulting from CO2 emissions and by the emphasis on the replacement of energy matrices. 4,6 –9 Recently, the Group of Seven (G7) officially announced the policy of decarbonization of the global economy, which aims to phase out fossil fuels by the end of the century, replacing them with renewable energy sources. This reduction in CO2 emissions is expected to help reach the goal of restricting global warming to 2°C. 10
Given the urge to replace fossil fuels, studies aimed at the development of technologies that use renewable resources to obtain energy and chemical compounds have become more frequent in recent years. Bioethanol is a promising biofuel that has the greatest potential to replace fossil-derived fuels with low economic impact, because the production of bioethanol has the potential to be less costly than fuel from non-renewable sources. 9 Moreover, bioethanol has properties that are beneficial to the environment, contributing to the mitigation of greenhouse gas emissions. 11 These properties are supported by the Carbon Intensity index (CI), which measures the amount of CO2 equivalent (CO2e) released for each unit of energy generated by fuel. Gasoline has a CI of about 95.86 g CO2e/MJ, whereas ethanol produced from cornstarch or sugarcane juice has an average CI of 75.97 g CO2e/MJ and 56.66 g CO2e/MJ, respectively. 12,13 Bioethanol can be blended with gasoline or can be used directly as a transportation fuel because it has a higher enthalpy of vaporization, higher octane number, and higher laminar flame speed than gasoline. In addition, bioethanol is an oxygenated fuel, which reduces nitrogen oxide emissions and, when blended with gasoline, allows complete combustion, reducing the emission of carbon monoxide and unburned hydrocarbons. 11
For these reasons, many countries, including Brazil, the United States, China, and several European Union member states have established programs to stimulate bioethanol production. 11 The US and Brazil are the first and the second largest producers, respectively, of ethanol worldwide. 14 In the US, approximately 54 billion liters of ethanol are produced annually, primarily from cornstarch using alcoholic fermentation. In contrast, sugarcane juice is the primary feedstock used in Brazil, with annual ethanol production of 25 billion liters. 14 –16 In both countries, alcoholic fermentation is an efficient system for ethanol production. However, high operational costs and limited productivity of sugarcane crops has affected the Brazilian ethanol industry in recent years. The use of lignocellulosic material presents an opportunity to expand ethanol production, as it may increase land productivity–the amount of fermentable sugars produced per hectare. In Brazil, the production of sugar and ethanol from sugarcane generates several hundred million metric tons of lignocellulosic biomass, which consists of bagasse resulting from sugarcane juice extraction and the sugarcane tops and leaves left in the field after harvest. 17 The production of ethanol using lignocellulosic feedstock can also enable year-round production because lignocellulosic material can be stored. 18
Many technologies for biomass processing are being developed to enable the use of cellulosic and hemicellulosic sugars by microorganisms. 19 –21 Simultaneously, the development of genetically modified microorganisms (GMOs) capable of efficiently converting C5 and C6 sugars into ethanol makes the industrial production of second-generation ethanol feasible. 22 Efforts to establish efficient technologies for cellulosic ethanol production are ongoing, and substantial investments in the development of a new type of cane—called energy cane—are being made. Instead of the high productivity of sugar in sugarcane, energy cane has a high yield of fibers and is more vigorous and robust than the conventional sugarcane; thus, it has several economic and environmental advantages as a feedstock for second-generation biofuel production. 17
This review will discuss first- and second-generation ethanol production in Brazil, a suitable country for the production of sugarcane and consequently ethanol due to high levels of tropical sunlight irradiation, an abundance of water, and great expanses of arable land. The goal of this review is to discuss the latest advances and current scenarios of both technologies, focusing primarily on second-generation ethanol production, considering the feedstock, sugar extraction, and fermentation processes for each and noting the economic and environmental advantages and disadvantages of each process. This discussion will support future studies and guide investment policies for the production of ethanol and other biofuels.
Saccharum Hybrids: The Ideal Feedstock for Biofuel Production
Industries based on agroforestry products have historically relied on dedicated crops delivered by genetic breeding programs and variety selection all over the world. The ideal feedstock for biofuel production should be highly productive and meet industrial demands, minimizing expenses and maximizing biofuel yields. Thus, the ideal crop will need to have fast growth, a flexible harvesting period and, in particular, a desirable carbon balance, which is assessed by considering all energy flow along the production chain and the environmental impact. 23 Life cycle assessment (LCA) not only considers productivity, water, and nutrient requirements, agricultural management, and residue production, but also depends on location and climate conditions. 23
Saccharum hybrids satisfy these needs. They can be planted across a range of tropical and subtropical environments and produce the largest biomass of any annually harvested field crop. 24 Saccharum hybrids have a high photosynthetic efficiency, as they are C4 plants, and also due to successful breeding programs, which have selected the most productive hybrids over the years. 17
Modern sugarcane cultivars are hybrids that have been selected by several backcrosses from genotypes selected from crosses between Saccharum officinarum and Saccharum spontaneum. 25 S. officinarum, the noble cane, is rich in reducing sugars, which are stored in thick and soft stalks due to the low fiber content of this cane. However, this species has a shallow root system, is highly demanding of soil fertility and water supply, and is highly susceptible to diseases. 26 Therefore, the wild species S. spontaneum was used for introgressive hybridization of complementary characteristics such as vigor and hardiness as well as tillering and ratooning abilities. 27 The rustic nature of this wild species, which is highly adapted to different environments, provided modern hybrids with resistance against several abiotic and biotic stresses. 25,26
In addition to field productivity, the sugar and fiber contents of industrial, dedicated Saccharum hybrids are the primary characteristics for breeding selection. These characteristics are directly related to the genetic contribution of each parent: the greater the presence of S. spontaneum chromosomes, the higher fiber content and lower sugar content the selected hybrid will have. 25 Analysis of the wide segregation and the resulting phenotypes indicated two distinct Saccharum hybrids: the traditional sugar-producer hybrid known worldwide as sugarcane, and a biomass-dedicated hybrid rich in fiber called energy cane (Table 1 and Fig. 1). 17,25,26,28

Comparison between cultivars of sugarcane (left) and energy cane (right). Even under stress conditions, the energy cane has a vigorous root system that allows a better use of water available in the soil, resulting in higher biomass productivity in the field. This property makes energy cane one of the most competitive raw materials for second-generation ethanol production.
Comparative Data Between Sugarcane and Energy Cane
Sugarcane: Feedstock for First-Generation Ethanol
Brazil started using sugarcane as a potential feedstock for bioethanol production in the 1930s, after the Institute of Sugar and Alcohol was created. However, the development of a competitive agroindustry in Brazil was not realized until after the Brazilian government established the National Alcohol Program (PROALCOOL) in the 1970s. 23 This national program stimulated not only greater process efficiency in industrial facilities, but also sugarcane-breeding programs. 17,23,25
Sugarcane is usually planted at the end of the wet season and harvested after the first cut (plant cane) every year for another 4 to 5 years (ratoons). Productivity (tons of cane per hectare) is higher with the first cuts and decreases during successive ratoons. In Brazil, approximately 9 million ha were planted with sugarcane in the 2014–2015 season, yielding approximately 635 million t of cane. The primary producer state was São Paulo (53.8%; 4.7 million ha), followed by Goiás (9.5%; 0.850 million ha), Minas Gerais (8.9%; 0.805 million ha), Mato Grosso do Sul (7.4%; 0.668 million ha), Paraná (7.1%; 0.635 million ha), Alagoas (4.3%; 0.385 million ha), and Pernambuco (2.9%; 0.260 million ha). 29
Sugarcane varieties have been developed by several research centers, including the Agronomic Institute of Campinas (IAC; Campinas, Brazil), federal universities linked to the sugarcane research network RIDESA (Inter-University Network for the Development of Sugarcane Industry), Centro de Tecnologia Canavieira (CTC; Piracicaba), and CanaVialis/Monsanto —the last two of which are private breeding companies. 30
Despite the high productivity of sugarcane varieties, first-generation technologies rely on sucrose content to produce bioethanol, leaving behind two parts of the total biomass, including structural sugars. The bagasse and sugarcane straw are usually burned to produce electrical power, which is used to run the sugarcane mill, and the excess energy is exported to the grid. One metric ton of harvested sugarcane stalks produces approximately 250 kg of bagasse (50% moisture) and 140 kg of cane straw/tops (dry basis). 31 Commonly called trash, the sugarcane tops and dead leaves (straw) that are left in the field represent 15% of the stalk weight of mature sugarcane. 17 This entire biomass residue has enormous potential to be used to produce electricity and to generate ethanol by second-generation technologies.
Energy Cane: Feedstock for Second-Generation Ethanol
The energy cane concept was initially described in Louisiana and Puerto Rico, where the first breeding attempts to obtain hybrids with high amounts of biomass regardless of sucrose content took place. This concept is now the focus of special attention as biomass has become an important source of renewable and sustainable energy. Energy cane is usually selected directly from crosses between S. spontaneum and either S. officinarum or pre-commercial sugarcane varieties, enhancing the genetic participation of the wild species. This crossing ensures not only a high yield of fibers in the selected hybrids, but also other important economic and environmental advantages. The energy cane hybrids are less demanding on agricultural inputs because they are more resistant to abiotic and biotic stresses. These advantages allow better use of water and nutrients, minimize the use of herbicides and pesticides, and permit growth in poor environments, which eliminates competition with food production. 17 In addition to the high fiber content and biomass productivity of energy cane, other important characteristics are the high tillering and high productivity in the ratoons, which enhance yield during the years of growth and increase the rate of multiplication.
In the US, several attempts to use energy cane as a potential feedstock included the release of some cultivars in Louisiana and Florida. In Brazil, several private breeding programs were started for the selection of energy cane hybrids. The Monsanto Company at its Canavialis facility selects hybrids producing 138% more total biomass per area and 235% more fiber. Other companies such as GranBio (Campinas) and Vignis (Santo Antônio de Posse) are also selecting and planning to release new energy cane cultivars soon. 17,25 Some traditional sugarcane research centers such as IAC, RIDESA, and CTC are also focusing part of their breeding programs on selecting energy cane varieties. 25
First-Generation Ethanol Production
The production and use of ethanol as a vehicle fuel have been regularly and legally practiced in Brazil since 1931 when the federal government mandated a 5% ethanol blend with gasoline. Since then, political and economic issues in Brazil have defined different phases of ethanol production. In 1973, the first oil crisis increased oil prices by approximately 300% (from $3/barrel to $12/bbl). 32 Consequently, the Brazilian government implemented PROALCOOL in 1975 to replace gasoline with ethanol obtained from biomass (sugarcane, cassava, and sorghum). For many reasons, including a favorable climate, an all-year growing season, and highly productive land, sugarcane became the sole source for ethanol production, allowing Brazil to produce over 35 million t of sugar and 28.6 billion liters of ethanol in the 2014–2015 season. 33
Before PROACOOL was established, Brazil fueled its automobile industry primarily with gasoline derived from petroleum, of which more than 80% was imported. According to the Brazilian National Agency of Petroleum, Natural Gas and Biofuels, at the end of 1979, the percentage of ethanol in gasoline blends increased and the automobile industry started using fuel blends up to E15 (15% anhydrous ethanol; 85% gasoline). 34 Consequently, ethanol production in Brazil increased by more than 12-fold during the first phase of PROALCOOL—from 220 million liters in 1975 to 2.8 billion liters in 1979. 35 At that time, anhydrous ethanol was produced in distilleries annexed to existing sugar mills.
In 1979, the world faced the second oil crisis. Crude oil prices increased from $25/bbl in 1979 to $40/bbl in 1981. This crisis motivated the large-scale production of ethanol and the expansion of sugarcane fields, enabling the construction of autonomous distilleries that produced only ethanol from sugars. In 1979, the Brazilian government signed an agreement with the vehicle industry whereby manufacturers were required to develop or adapt motor engines to use hydrated ethanol as fuel (E100). This fuel had a minimum ethanol content of 92.6%. 34 Since then, ethanol began to be used as a fuel substitute and, in 1983, neat ethanol-powered cars accounted for more than 90% of total automobile sales. 32 Nevertheless, anhydrous ethanol remained in use as an additive, and in 1985, the ratio of ethanol to gasoline was set between E20 and E25. 32
In 1986, lower oil prices and a severe economic crisis in Brazil caused neat ethanol-powered vehicle sales to decline. By 1989, the inflation rate was 1,764.86%, the highest recorded in the history of Brazil. 32 This characterized the beginning of a decline phase of PROALCOOL. The reduction of the neat ethanol fleet greatly affected the consumption of hydrated ethanol during the 1990s and early 2000s. Despite worldwide concern over environmental sustainability, which led to the signing of a document on climate change at the Rio 92 Conference (1992), it was not until 2001 that the substantial price difference between ethanol and gasoline contributed to an increase in sales of neat ethanol-powered vehicles. In 2003, the decline of PROALCOOL was completely reversed with the introduction of flex-fuel vehicles (FFVs) in Brazil, which allowed customers to fuel with both ethanol/gasoline blends (E25) and hydrated ethanol (E100). Due to the success of FFVs, the production of ethanol greatly increased and, in 2007, ethanol production reached 21.6 billion liters, 3.5 billion liters of which were exported. 36
The sugar-ethanol industry has made remarkable progress in recent decades. Since the turn of the century, ethanol production in Brazil has increased up to 170%, and future projections for Brazilian ethanol are optimistic. Domestic consumption of ethanol as a biofuel is increasing, and many countries have already approved the use of ethanol/gasoline blends, which should increase exportation. 33
Ethanol Mills in Brazil
Currently, Brazil has approximately 345 ethanol mills that produce more than 28.66 billion liters of ethanol each year. 15,16 This large production is due to three particular characteristics that differentiate the sugarcane industry in Brazil from its peers in other countries. First, most units in Brazil produce a rather high proportion of processed sugarcane, with only one third of processed feedstock acquired from third parties. In contrast, the international standard is to separate agricultural activity from industrial production. This organizational model is associated with the large territorial size of Brazil, the wide availability of fertile land that is suitable for the cultivation of sugarcane, and the agrarian tradition of Brazil.
The second distinct characteristic of Brazil's sugarcane industry is the diversity of traded products made from the sugarcane juice and residues from milling. In addition to the sugar and ethanol produced, the excess electricity generated by burning bagasse/straw is sold to the grid. Finally, the spatial distribution of production units in a large area is possible due to Brazil's geographical position, which provides a wide variety of microclimates that make industrial-scale production of most commercial crops used worldwide economical.
In 2013, the National Food Supply Agency published the latest survey of the Brazilian sugarcane industry, concerning the 2010–2011 harvest. 15 This survey included data from 415 units in operation during that period, with 328 located in the Center-South region and the remaining 87 in the North-Northeast region. 15,16 Table 2 presents the primary productivity numbers and sizes of those units and highlights the Center-South region, which is responsible for approximately 90% of the sugar and ethanol produced in Brazil. These regions also differ regarding the growing season. During the months of April to November, the low amount of rainfall in the Center-South allows for the sugarcane harvest. In contrast, the growing season in the North-Northeast spans the months of August to February. Therefore, plant operations remain idle for a long period, operating an average of 212 days per season. The states of Mato Grosso do Sul, Paraná, and São Paulo have the best performance, with operational averages of 246, 238, and 237 calendar d/y, respectively. 15
Production Data from the Last Five Seasons and Unit Ratings According to the Volume of Processed Sugarcane
During the most recent season (2014–2015), the Center-South region presented a slight decrease in sugarcane crushing (4.3%), compared with the 2013–2014 season, processing 571,344 million t. The crushing expectation for the 2015–2016 season is for the same level as the 2013–2014 season, due to a reduction of investments in cane replanting that has increased the average age of the cane fields. Consequently, ethanol production must also remain stable in the 2015–2016 season, with an average idle capacity of 57.5%. 16
The vast majority of ethanol distilleries have integrated sugar mills that allow sugarcane to be processed into either sugar or ethanol. The production mix of such units has remained relatively stable over the past 10 years, with an average of 46% of sugarcane dedicated for the production of sugar and 54% to ethanol. 16 This integration is another feature of Brazilian plants, which can vary the destination of sugarcane juice based on the attractiveness of ethanol and sugar markets. The limitation of this variation is the productive capacity installed in each unit.
As a result of the global financial crisis that erupted in 2008, by 2014, 82 mills had ceased operations in Brazil. Of those, 59 were operating in the Center-South region (60 million t of sugarcane) and 23 in the North-Northeast (15 million t of sugarcane). According to market sources, other plants were expected to declare bankruptcy by the end of 2015 and early in 2016. 16
In contrast to the US, only two corn ethanol plants are in operation in Brazil, with a combined capacity of 145 million L/y. In the next 2 years, four more units will begin operations in the Midwest region, where corn production is primarily concentrated. These units will have a total capacity of 500 million L/y and will use the surplus volume of corn available in Brazil in a dedicated plant. Recent studies have shown the feasibility of integrating these corn mills with sugarcane mills, sharing the already installed facilities (steam and electricity) and producing ethanol throughout the entire year. 37
Yeasts for First-Generation Ethanol Production
Saccharomyces cerevisiae is the most employed yeast for ethanol production in Brazil. S. cerevisiae can assimilate and ferment a large number of carbon sources to produce ethanol. However, this yeast is not naturally capable of utilizing sugars obtained from lignocellulosic biomass such as xylose and arabinose.
The Brazilian model for ethanol production using S. cerevisiae as inoculum has been extensively described in a number of reviews. 38,39 This model is primarily characterized by the combined production of sugar from sugarcane juice and ethanol from sugarcane juice and molasses. Fed-batch ethanol fermentation begins using known specific mixes of commercial dry yeasts as inoculum. At the end of fermentation, cells are separated from fermented wine, and after 2–3 hours of acid treatment, they are reused to start a new fermentation process. The high cell density (12–18% w/v, wet basis) used in each batch ensures a short fermentation time, reaching ethanol concentrations of 7–11% (v/v)—90–92% of the theoretical sugar conversion into ethanol—in 6–11 h. Cell recycling reduces the need for inoculum propagation; thus, less sugar is destined for biomass formation. Moreover, cells can be reused 400–600 times during the entire sugarcane harvest season, which takes 200–300 days depending on weather conditions, sugarcane variety, and market demands. 38
As industrial ethanol production is performed under non-sterile conditions, different yeast and bacterial species originating from the feedstock and/or environment may compete with the inoculated yeast. Both the persistence and dominance of these species will depend on their interaction during the entire process, on the medium composition, and on the fermentation conditions, as well as on the capability of the microorganism to tolerate several stresses generated by the process itself, such as high ethanol concentration, high temperature, and acidity. Therefore, inoculated yeast could rapidly be replaced by indigenous strains that are more robust during the fermentation process. 38,40
First-generation ethanol production in Brazil began using S. cerevisiae (baker's yeast; Fleischmann's® and Itiaquara®) as inoculum. However, in the early 1990s, Brazilian distilleries started to isolate and select wild-type yeast strains that showed high persistence profiles during the entire season to start the following season using a more robust and adapted strain. 39 Since then, the use of different molecular methods, which have been described in several studies, have revealed the heterogeneity of the yeast population and demonstrated the dynamics between microorganisms during industrial fermentation processes in some distilleries from different regions of the country. 39 –41
Currently, commercial powdered yeast strains selected from industrial processes are widely used as inoculum in distilleries. The most successful examples are the CAT-1, BG-1, and PE-2 strains, which were isolated from the Catanduva, Barra Grande, and Pedra distilleries, respectively. These strains have been adopted by most Brazilian distilleries. Additionally, other strains such as SA-1, VR-1, and CR-1, also isolated from industrial fermenters, have been observed during industrial processes but have shown low persistence and dominance profiles. 39
By 2010, up to 30% of Brazilian distilleries used the PE-2 strain as inoculum. 41 The genome structure of JAY270, a strain derived from the highly adapted PE-2, has already been described. 42 This work led to a better understanding of the adaptability and higher stress tolerance of PE-2 to industrial fermentation processes. Another study described the response and transcriptional reprogramming of S. cerevisae due to bacterial contamination at industrial scale. 43 The authors showed that Lactobacillus fermentum was the primary bacterial species responsible for yeast co-aggregation, one of the most serious problems in the industrial process that leads to lower yeast performance. Such work is essential for understanding the molecular basis of the ethanol fermentation process and for providing new knowledge regarding yeast, thus improving robustness and fermentation performance.
Second-Generation Ethanol Production
Lignocellulosic biomass is formed by cellulose (a polymer of β-1,4-linked glucose units with a crystalline structure), covered by hemicellulose (a polymer composed of different fractions of glucose, galactose, mannose, xylose, and arabinose units presented in amorphous form) and lignin to provide protection against chemical and biological degradation. 44 The composition of the primary components ranges from 40–50% cellulose, 20–40% hemicelluloses, 20–30% lignin, and 5–8% extractives. Minor components of lignocellulosic biomass include proteins, lipids, pectin, soluble sugars, and minerals. One of the major challenges of second-generation technology is the difficulty of obtaining the sugars from these main components due to the complex and rigid structures of plant cell walls. 44,45 Therefore, pretreatment and enzymatic hydrolysis of biomass are necessary steps, as is the development of genetically modified microorganisms capable of consuming these sugars.
Pretreatment
Different pretreatment strategies have been developed to modify the morphological and chemical arrangements of the cell wall from lignocellulosic biomass to facilitate the hydrolysis of polymeric sugars into monomers. 46,47 During pretreatment of the lignocellulosic material, hemicellulose and part of the lignin are solubilized, while cellulose is partially depolymerized. 48,49 This process can maximize sugar digestibility by increasing the material's surface area, consequently increasing the rate of enzymatic hydrolysis.
During pretreatment, hemicellulose is broken into oligomers and xylose, arabinose, galactose, mannose, acetic acid, and glucose; only a small fraction of cellulose produces glucose. Under certain pretreatment conditions, xylose and arabinose are decomposed to furfural; similarly, hydroxymethylfurfural (HMF) is formed by dehydration of hexoses (glucose, mannose, and galactose). If conditions persist, furfural can be decomposed to generate formic acid, and HMF can be broken down into formic acid and levulinic acid. 50 Acetic acid is released from acetylated hemicellulose during pretreatment or hydrolysis and is one of the primary lignocellulosic inhibitors, present at significant concentrations in most hydrolysates. Moreover, during pretreatment, the solubilized lignin can be decomposed into phenolic compounds. Overall, these products are considered inhibitors of microorganism metabolism and decrease the performance of the fermentative processes ( Fig. 2 ). Several pretreatment technologies exist and can be grouped into four types: chemical (acid, alkaline, oxidative delignification, and organosolv), physical (milling, pyrolysis, and microwave), physicochemical (steam explosion, ammonia fiber explosion, and CO2 explosion), and biological (biopulping via fungi, bacteria, and specific enzymes). 21

Main routes of inhibitor formation. Lignin decomposition generates phenolic compounds, while furan derivatives and aliphatic acids are formed as a result of carbohydrate degradation from hydrolysis of cellulose and hemicellulose.
Chemical Pretreatments
The
Physical pretreatment
Physicochemical pretreatments
During
Biological pretreatment
Biological pretreatment uses microorganisms (fungi and bacteria) to degrade lignin and hemicellulose. This process is mild, with low energy use, but due to microorganism metabolism takes days to weeks for completion. Moreover, some of the released sugars are used as carbon sources by the microorganisms, which leads to low efficiency. 20,45
Enzymatic Hydrolysis
After pretreatment of the lignocellulosic biomass, a large sugar fraction remains in the form of polysaccharides and oligosaccharides. These polymers must be further broken down into sugar monomers to allow for their conversion into ethanol. Several approaches for the breakdown of cellulose have been studied, including enzymatic hydrolysis, which employs protein blends commonly termed “enzymatic cocktails” to cleave glycosidic linkages, and acidic hydrolysis, which makes use of strong acids such as H2SO4. 63,64 So far, enzymatic hydrolysis has been the preferred approach for industrial applications due to the amenable working conditions (e.g., 50–60°C and pH 4.5–5.5), higher yields, and reduced utilities consumption. In addition, enzymatic hydrolysis reduces the formation of fermentation inhibitors and equipment corrosion, issues commonly associated with acidic hydrolysis. 65
A number of enzymatic cocktails have been developed and are commercially available for industrial applications. These cocktails contain groups of enzymes that break down cellulose and hemicellulose into glucose, xylose, arabinose, and other monosaccharides. The hemicellulose fraction of biomass, due to its lower degree of polymerization and highly branched nature, is more easily depolymerized than cellulose, which has a crystalline structure. In this review, only the enzymatic hydrolysis of the cellulose will be discussed.
Mechanism of enzymatic hydrolysis
The conversion of cellulose to glucose requires the involvement of three enzyme groups: exoglucanases or cellobiohydrolases (CBHs), endoglucanases (EGs), and β-glucosidases (BGs). CBHs are divided into CBHI and CBHII. Both enzymes attack the crystalline side of cellulose; however, the former works on the reducing ends, while the CBHII acts on the nonreducing side of cellulose. The disaccharide cellobiose is a product of these enzymes. The EG group cleaves the amorphous regions of cellulose and provides new chain ends for the action of CBHs, resulting in oligomers that will be hydrolyzed to cellobiose later by CBHs. The BGs cleave the glycosidic bond of cellobiose, releasing glucose as the final product of the hydrolytic process. 44
In addition to these central groups of enzymes described above, there are also the so-called “accessory enzymes,” reported to increase the yield of released sugars during cellulose hydrolysis. The most studied of these enzymes are xylanase and the recently discovered lytic polysaccharide monooxygenases (LPMOs). 66 Xylanases catalyze the conversion of hemicelluloses into monosaccharides, while LPMOs catalyze the oxidative cleavage of cellulose, introducing chain breaks in this polysaccharide. Both favor the access of cellulases to cellulose. 44 LPMOs represent a new approach in the deconstruction of lignocellulosic biomass by enhancing the activity and decreasing the loading of classical enzymes. The addition of these enzymes to enzymatic cocktails improves hydrolysis effectiveness at high consistencies and reduces the costs of the hydrolysis step. They could be important for the future of biorefineries. 44
Enzyme cocktail production
Enzyme cocktails for cellulose hydrolysis are used in several industries, including textile, detergent, food, pulp and paper, and corn ethanol. However, it is the production of biofuels such as ethanol and butanol and other biochemicals derived from biomass that could turn the cellulase market into the largest worldwide enzyme market. 67 In addition, the development of cellulase production at competitive cost is an important issue and a requirement for the economic feasibility of second-generation ethanol production.
Currently, commercial enzyme preparations available for lignocellulosic biomass hydrolysis are produced by fermentation of genetically modified strains of Trichoderma reesei. 68 This filamentous fungus is known to be an efficient producer of cellulases and hemicellulases, which act in synergy to degrade lignocellulosic materials. 69 However, other filamentous fungi are also capable of producing efficient cellulolytic systems, such as those belonging to the genera Humicola, Chrysosporium, Penicillium, Acremonium, and Aspergillus. 70,71 Among these genera, Aspergillus has received special attention as an interesting alternative to T. reesei in the search for new commercial preparations and processes that more efficiently hydrolyze cellulosic substrates. 70,71
The cellulolytic complex secreted from strains of T. reesei is known to have low levels of β-glucosidase enzyme production (approximately 0.5%). 68 This low level of production can result in cellobiose accumulation, which will inhibit the CBHs and consequently reduce hydrolysis efficiency. 72 Thus, external supplementation with β-glucosidase is generally required from other sources of enzyme complexes such as Aspergillus sp. 73 Another strategy to improve β-glucosidase productivity in T. reesei is through heterologous expression of this enzyme derived from Aspergillus sp. or from other filamentous fungi. 74 New approaches for cellulase production are now in development, including cellulosome complexes derived from bacterial genera such as Cellulomonas, Thermobifida, Ruminococcus, and Clostridium. 75
Although cellulolytic enzyme production as a commercial cocktail is already available for use in the conversion of lignocellulosic biomass at industrial scale (i.e., Cellic® CTec3 and Accellerase), the cost still needs to be optimized to make second-generation ethanol production more competitive. Thus, diverse strategies, including genomics, proteomics, protein engineering, directed evolution, and optimization of culture conditions are being extensively applied to improve the properties of enzymes and microorganism producers. 76
Challenges of industrial enzymatic hydrolysis
Industrial-scale hydrolysis has serious challenges still to be overcome in terms of efficiency, costs, dosages, hydrolysis time, and process configuration. Hydrolysis time, hydrolysis at high solid concentrations, and enzyme dosage must be improved to achieve high hydrolysis performance. Hydrolysis time is important for reducing the capital and operational costs; the shorter the residence time, the smaller the tank volume and peripheral equipment (pumps and heat exchangers) required for the process. Then, the hydrolysis process should be performed in as short a time as possible. Currently, industrial-scale hydrolysis should not take longer than 72 hours.
The total solids at the beginning of hydrolysis is directly related to the sugar content at the end of hydrolysis, the higher the total solids are at the beginning of hydrolysis. The higher the sugar concentration of the hydrolysate will be at the end of hydrolysis. High sugar concentrations should allow higher ethanol titers to be obtained during fermentation, allowing for the reduction of fermentation tanks volumes and a more profitable ethanol distillation process. Therefore, hydrolysis should be performed with 20–25% w/w of total solids to reach between 10–12% w/w of fermentable sugars (considering 70% of hydrolysis yield minimum) and obtain an ethanol concentration greater than 4% w/w (considering about 88–92% fermentation yield). The enzyme cost is considered to be approximately 20% of the ethanol production cost. 77 Enzyme dosage optimization without increasing hydrolysis time or yield loss is an important issue for the economic feasibility of industrial enzymatic hydrolysis.
Second-Generation Ethanol Fermentation
Lignocelullosic hydrolysates are rich in sugars that are easily fermentable, such as glucose, mannose, and galactose, and sugars that require the use of genetically modified S. cerevisiae to be converted into ethanol, such as xylose and arabinose. Genetic modification of yeast for C5 sugar consumption will be examined further in this review. Here, the factors related to processes that have relevant influence on second-generation fermentation, such as yeast propagation, will be discussed.
Yeast propagation
In the context of fermentation, yeast propagation is necessary to obtain sufficient cell mass to start the fermentation process. As explained above, in the Brazilian process for producing first-generation ethanol, this step was not a usual part of the process because fermentation is performed using cell recovery by centrifugation during the entire season. Thus, the cell mass produced during fermentation is used as inoculum for the next fermentation. However, for second-generation production, yeast recovery is more difficult to perform due to high concentrations of insoluble solids such as lignin, insoluble unhydrolyzed sugar, and insoluble ashes. Even if yeast recycling were possible, it would not be recommended because the genetically modified yeast could be replaced by wild-type strains and other contaminants. Therefore, yeast propagation is an important step before every batch fermentation for second-generation ethanol production.
S. cerevisiae has the ability to grow using anaerobic or respiratory metabolism depending on culture conditions. For ethanol production, anaerobic metabolism is preferred; however, for yeast propagation, a complete respiratory metabolism is essential to reach high cell yields. Under total respiratory metabolism, the yield is 0.5 g cell mass per gram of glucose. 78
Yeast propagation is a well-understood process that depends on several factors including sugar concentration, aeration, culture medium composition, yeast strain, and reactor configuration. To promote respiratory metabolism in S. cerevisiae, high rates of aeration (approximately 1–2 vvm), sugar concentration lower than 5 g/L, and a specific bioreactor configuration (i.e., airlift) are necessary. The culture medium is another important factor. For industrial yeast production, sugarcane molasses is commonly used as the culture medium basis. This matrix contains not only sugars but also phosphates, salts, and vitamins required for yeast growth. When using sugarcane molasses, only a nitrogen source such as urea or ammonium is normally supplemented. In the Brazilian scenario, the availability of sugarcane molasses is not a problem due to the stability of the sucrose industry; however, if molasses is not available, then medium development will be necessary. In general, sucrose (VHP sugar), urea, potassium monophosphate, vitamins (i.e., biotin and thiamine), and minerals (Mg+, Mn+, Fe+, and others) are added to supplement the culture medium.
A robust yeast strain is also needed for efficient yeast propagation. The efficiency of respiratory metabolism can vary according to the yeast strain used. To begin the development of a second-generation yeast strain (xylose-consuming strains), the selection of suitable hosts with high performance under respiratory metabolism is also necessary. During the development of a yeast strain for xylose consumption, the yeasts are submitted to long periods of evolutionary assays in addition to metabolic engineering. During these periods, robustness and other features such as the capability to grow under aerobic conditions may be lost. Therefore, constant checking of the intrinsic capabilities of the strains during development is strongly recommended.
A robust strain and a well-designed process can ensure large amounts of yeast in a short culture period, with low production costs. Large amounts of yeast are desired for the next stage of fermentation because a high mass of yeast as inoculum can reduce the fermentation time and consequently increase ethanol productivity.
Fermentation
A successful fermentation is one that reaches high yields and high productivities and produces high ethanol concentrations. For first-generation fermentation, these performance parameters seem to have been reached. But for second-generation fermentation, the scenario remains under extensive development. The primary issues that need to be optimized are robustness of the genetically modified yeast, xylose consumption rate, inhibitor tolerance, culture medium cost, and process configuration. Therefore, for this process to be successful, a combination of a robust yeast strain with a well-designed fermentation process configuration is essential. Below, the more accepted process configurations that have been applied at industrial scale will be briefly discussed.
Usually, due to the close relationship between hydrolysis and fermentation, both process configurations are designed together. Approaches include separate hydrolysis and fermentation (SHF), simultaneous saccharification and fermentation (SSF), hybrid hydrolysis and fermentation (HHF), and consolidated bioprocessing (CBP). Currently, SHF and HHF stand out because developed technologies for industrial enzyme hydrolysis and fermentation (enzyme cocktails and commercial yeast) are related to these concepts.
SHF involves hydrolysis and fermentation in separate tanks, allowing optimal conditions for each process (hydrolysis at 50–55°C and fermentation at 30–35°C). Drawbacks to this process include enzyme inhibition via byproducts in addition to the high capital and operational costs (additional tanks and high energy requirements). In contrast, HHF involves hydrolysis in a separate tank until glucose release decreases. The reaction is then transferred to another tank and the temperature is lowered for fermentation, during which hydrolysis continues at low rates concomitant with ethanol production. Hence, this process can optimize capital and operational costs compared with SHF because tank volumes and residence times would be lower. Nevertheless, the success of this process depends on fine tuning hydrolysis to fermentation transfer, enzyme features, and hydrolysis at high total solids concentration. 68
SHF and HHF are currently used commercially in Brazil. The Iogen technology, implemented by Raizen (São Paulo), is a SHF process, while the PROESA technology used by GranBio at BioFlex 1 is similar to the HHF concept. 79,80
In addition to the above considerations, during hydrolysis processes with 20–25% total solids and independent of efficiency, the hydrolysates should have approximately 5–10% w/w insoluble solids, which are composed primarily of lignin, insoluble sugars, and ashes. These solids (primarily lignin) are important because they will be directed to the boiler for steam and energy generation. However, the stage at which the solids will be retired is another key factor that determines process configuration.
Solids separation can be performed upstream or downstream of fermentation. Removing the solids upstream of fermentation can allow easy integration with first-generation distillation or even with first-generation fermentation (if only C6 is used) by mixing first- and second-generation streams (at a beer well or at a fermentation tank), thus gaining process flexibility. This configuration can reduce the investment cost by using equipment already installed at first-generation mills, which reduces the distillation cost impact of a low ethanol concentration (4% w/w) and even allows cell recovery. However, solids separation before fermentation can cause significant sugar losses (since sugars would be dragged with solids during separation) and restrict the process for the use of SHF.
The other alternative is solids separation after distillation, which avoids sugar losses and allows for the utilization of SSF or HHF approaches. However, it involves special equipment design (fermentation tanks, pumps, heat exchanger, distillation equipment, and cleaning system) for working with high solid contents. For example, Iogen technology uses solids separation upstream of fermentation, whereas PROESA technology removes the solids after distillation.
Based on the process information presented above, it is possible to compare first- and second-generation ethanol production, considering the different steps involved in each process and the use of different varieties of cane. Figure 3 summarizes the levels of production of first- and second-generation ethanol that can be achieved, including the productivity, sugar content, and fiber content of sugarcane and energy cane cultivated in 1 ha of arable land. 81,82 Despite the lower soluble sugar content present in energy cane, its juice can also be used for the production of significant amounts of ethanol. The lignocellulosic material of sugarcane and energy cane (bagasse, tops, and leaves) can be processed for ethanol production or as fuel for boilers to produce the heat and electricity required to supply the plant during its operation. However, given the high productivity and fiber content of energy cane, more second-generation ethanol or more energy can be generated from this biomass compared to sugarcane. Although representing overall numbers from ethanol production, the data in Fig. 3 highlight the potential of energy cane as a substrate for fermentation processes and how much ethanol production can be improved using second-generation technology.
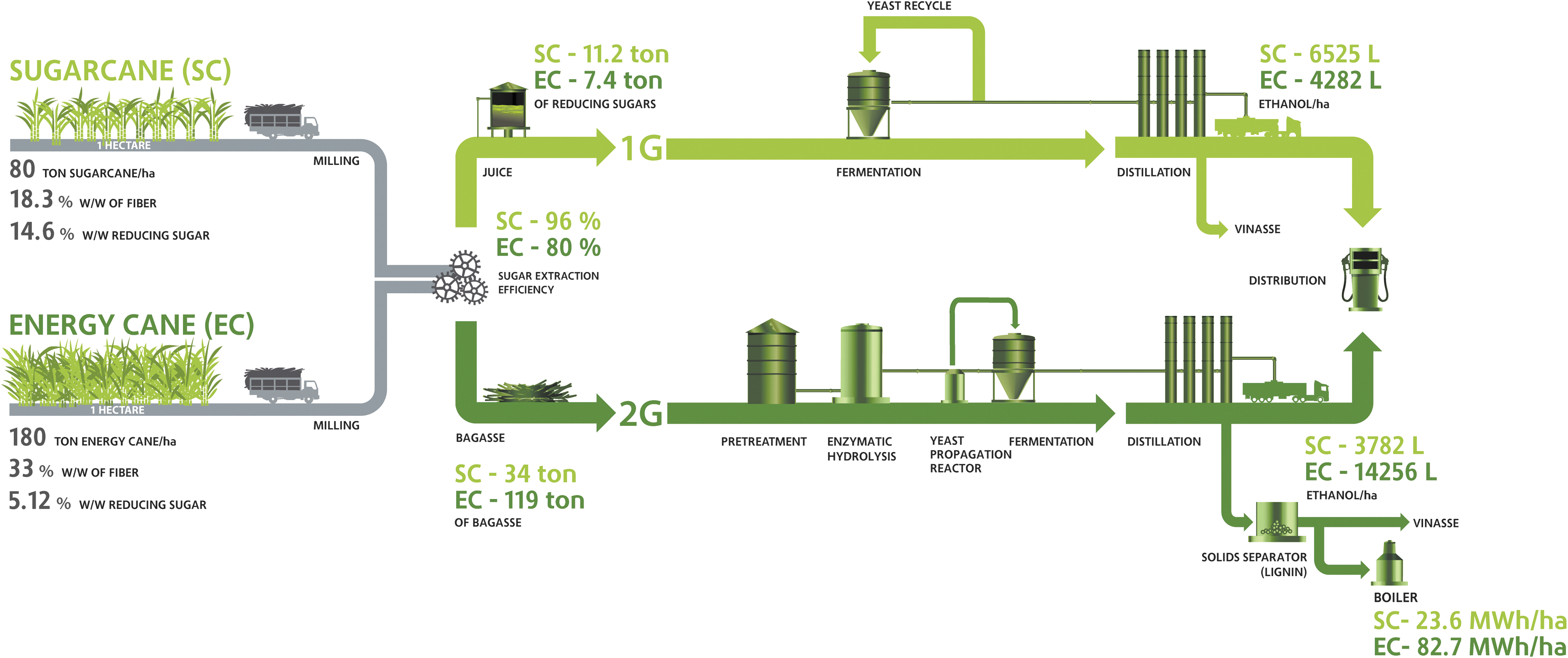
Schematic representation of potential first- and second-generation ethanol production from 1 ha of sugarcane (SC) and energy cane (EC). The calculation of ethanol production and energy generation was based on data already published and unpublished data from energy cane cultivars from GranBio Investimentos SA, considering the same efficiency for fermentation process and distillation in the first- and second-generation technologies. 81,82 The total amount of lignocellulosic material, including bagasse, leaves, and tops, was expressed in bagasse equivalent with 50% w/w of humidity. Sugar losses were not considered only in the pretreatment and hydrolysis of biomass. The calculations consider only the potential energy and ethanol that can be produced and do not take into account the energy required to operate the plant.
First- and-second generation processes can also be integrated. The lignocellulosic residues from the first-generation process (bagasse and straw) can feed the second-generation process. In the cogeneration unit, residues from the second-generation process (mainly lignin) together with bagasse and straw supply the steam and electricity for both processes, and the surplus can be sold to the grid. 18 Brazil currently has a great competitive advantage for the installation of new integrated units since the mills already installed provide raw materials and facilities for such deployment. The integrated interface industry is comprised of production areas traditionally found in current first-generation sugarcane processing, such as cane reception, extraction, processing, and concentration of the juice; C12 carbon stream fermentation and distillation; and generation and distribution of steam and electricity. The second-generation sector also has areas for biomass processing, such as pretreatment, hydrolysis, propagation of GMOs, and dedicated fermentation tanks for C5 or C5 and C6 carbon streams. 81
Yeasts for Second-Generation Ethanol Production
As previously mentioned, lignocellulosic biomass is composed of many sugars that are present in cellulose and hemicellulose chains and can be naturally fermented by S. cerevisae, such as glucose, mannose, fructose, and galactose. However, the pentoses derived from hemicellulose, L-arabinose and D-xylose, cannot be consumed by wild-type strains of this yeast. Because D-xylose is the most abundant pentose, composing up to 25% of the total sugar content in some hydrolysates, strains able to consume this sugar efficiently are essential to make the process economically viable. 55,83
S. cerevisiae cannot convert xylose to ethanol. However, it is capable of fermenting xylulose, an intermediate of xylose catabolism. This was demonstrated when the xylose isomerase enzyme was added to the extracellular medium and xylose consumption and ethanol production were observed. 84 Because S. cerevisiae can ferment xylulose, many efforts have been employed to introduce the two better-known pathways for xylose consumption. In bacteria, the conversion of xylose to xylulose can be performed in one step with the enzyme xylose isomerase (XI). This enzyme is most common in bacteria; it has also been found in some anaerobic fungi. 85,86 The second pathway, which is found in most naturally xylose-utilizing fungi, is a two-step reduction-oxidation reaction catalyzed by xylose reductase (XR) and xylitol dehydrogenase (XDH). In the next step, xylulose is phosphorylated by the enzyme xylulokinase (XKS1) to xylulose 5-phosphate, which is metabolized through the pentose phosphate pathway and then channeled into glycolysis to ethanol.
Some yeasts, such as Scheffersomyces stipitis, Candida shehatae, and Spathaspora passalidarum, are naturally capable of fermenting xylose and converting it to ethanol using the two-step oxidative-reductive pathway. 87 However, S. cerevisiae is still the primary choice, being the most-used microorganism for ethanol production in industrial processes and having been demonstrated to be able to ferment sugars present in lignocellulosic hydrolysates with a high content of inhibitors. 55
The XR/XDH pathway
S. cerevisiae has genes from the oxidative-reductive pathway XR and XDH in its genome, but they are poorly expressed and the yeast is not able to grow on xylose. 88 The most common approach to engineering S. cerevisiae for xylose consumption utilizes the genes XR and XDH from S. stipitis, coupled with overexpression of the gene XKS1 from S. cerevisiae. In the first reaction, xylose is reduced to xylitol by the enzyme XR using nicotinamide adenine dinucleotide phosphate (NADPH) or nicotinamide adenine dinucleotide (NADH) as a cofactor, although the majority of XRs have a higher affinity for NADPH. Xylitol is then oxidized to D-xylulose by the enzyme XDH exclusively using NAD+ as a cofactor. Under anaerobic conditions undesirable fermentation by-products accumulate, such as xylitol and glycerol, thus negatively affecting the ethanol yield from D-xylose. 83,89
The preference to express the genes of S. stipitis in S. cerevisiae is made because of the high yield of ethanol that can be achieved from xylose using this yeast under controlled oxygen conditions. Most other yeasts that are able to consume xylose primarily produce the undesirable byproduct xylitol. 87,88 The S. cerevisiae strains transformed with these genes, although capable of xylose consumption, still accumulate considerable quantities of xylitol and consequently have low ethanol yield caused by the imbalance of cofactors. 89 Additional metabolic engineering steps were applied to enhance the xylose flux, and several approaches to regulate the redox balance of cofactors in these strains were tested.
Manipulations in redox balance and the amount of cofactors produced by the cell can be a powerful tool for improving the fermentation performance of the yeast. 90 Improving the redox balance in xylose-modified S. cerevisiae strains could be achieved by several approaches that model the cofactors flux into the cell to adjust the levels and make them available for the heterologous enzymes of the xylose pathway. This improvement is necessary to achieve a strain with low byproduct production and high ethanol yield and productivity.
The xylose isomerase pathway
The isomerization of xylose to xylulose occurs in a single step via the xylose isomerase enzyme. This enzyme does not use cofactors; therefore, no redox imbalance and no accumulation of byproducts occur, and a higher yield of ethanol can be reached. Based on nucleotide sequences, xylose isomerases are divided into two families: family I has GC-rich DNA content and is shorter by 40–50 residues at the N-terminus, whereas family II has low GC content and an extended N-terminal region, forming a more diverse group. 86,91
For many decades, attempts to express heterologous bacterial XI in S. cerevisiae have not been successful. Attempts to express XI from Escherichia coli, Bacillus subtilis, Actinoplanes missouriensis, Clostridium thermosulfurogenes, and Streptomyces rubiginosus have failed to produce a functional enzyme in S. cerevisiae. 83,92 –95 The most likely reasons to explain the inactivity of this enzyme in S. cerevisiae are improper folding of the protein, the internal pH of the yeast cells being different from that of the original organism, and the absence of an essential cofactor or metal ion. 92,95 The first positive result was the xylose isomerase from Thermus thermophilus, which was functionally expressed in S. cerevisiae, but with low activity, because T. thermophilus is a thermophilic organism and the optimum activity of this enzyme occurs at 85°C. 96
In 2003, after the discovery of xylose isomerase in the anaerobic fungus Piromyces sp., it was possible to obtain mutants of S. cerevisiae that had high activity of this enzyme, with values ranging from 0.3–1.1 μmol/mg protein/min and capable of growth on xylose as the sole carbon source. The functional expression of this gene in S. cerevisiae is probably due to the similarity of mechanisms for protein re-folding and the cytosol conditions between these two eukaryotes. 85 Although this strain is able to grow on xylose as the sole carbon source, it presents slow growth and incomplete xylose consumption. Other modifications were necessary, including evolutionary engineering procedures for obtaining a strain capable of rapid anaerobic xylose fermentation. 97 In 2009, the expression of XI from the anaerobic fungus Orpinomyces sp. also resulted in a strain of S. cerevisiae with high fermentation yield. One reason for this enzyme's activity in S. cerevisiae is that unlike most XIs, which have optimum activity at temperatures above 60°C, the Orpinomyces XI presents optimal activity at 37°C, near the growth temperature of S. cerevisiae. 86
After many attempts to express a prokaryotic XI with high activity in S. cerevisiae failed, the XI from the anaerobic bacterium Clostridium phytofermentans presented kinetic parameters and activity in S. cerevisiae comparable to these anaerobic fungi at 30°C, despite having low sequence similarity to the XI's from Piromyces sp. and Orpinomyces sp. Moreover, this enzyme presents an advantage of demonstrating lower inhibition by xylitol, a difference from the other XIs. 98 Thus far, a small number of xylose isomerase genes have been functionally expressed in S. cerevisiae, leading to efforts to prospect new enzymes with high activity in yeast. The efficient expression of the pathway that uses xylose isomerase in S. cerevisiae represents advantages compared to the route that uses the enzymes xylose reductase and xylitol dehydrogenase, avoiding the imbalance of cofactors in the strain and generating a yeast with higher ethanol yield.
Improving xylose consumption in engineered yeast strains
The engineering of xylose metabolic pathways in S. cerevisiae is not sufficient to achieve xylose fermentation at industrially relevant rates. Generally, genetic engineering must be followed by an evolutionary engineering step in which the yeast is continuously propagated in xylose-containing medium. 99,100 This step can be performed through repeated batch cultivation or continuous growth in chemostats and, depending on the ability of the recombinant yeast to grow on xylose, low amounts of glucose may be added. As the yeast grows, random mutations can occur, increasing the genetic and phenotypic diversity of the initial monoclonal population. Isolates carrying mutations that confer faster growth on xylose will be selected for and increase in frequency through time. Although unable to ferment xylose, engineered yeast strains have been reported to grow well on xylose-containing medium under aerobic conditions. 100,101 Efficient xylose fermentation can be achieved through evolutionary engineering by gradually decreasing the oxygen levels available to the evolving culture. Several yeast strains capable of efficiently fermenting xylose have been described in the literature, and all of these strains underwent a period of evolutionary engineering. 22,101,102 However, the genetic basis of their evolved phenotypes has yet to be thoroughly elucidated. As this information becomes available, reverse engineering the identified mutations into recombinant strains and developing rapid xylose-fermenting yeasts in a short period of time will be possible.
A number of rational engineering approaches have also been applied to improve xylose utilization in engineered yeasts with varying degrees of success. 103 For instance, the over-expression of endogenous enzymes of the non-oxidative part of the pentose phosphate pathway has been shown to result in significant improvements. 101,103 Another commonly employed modification is the deletion of the GRE3 gene in strains harboring XI pathways. GRE3 encodes an aldose reductase that can convert xylose into xylitol. Xylitol is a known inhibitor of XI, and the GRE3 deletion has been reported to increase xylose fermentation rates. 101 In recombinant xylose-utilizing S. cerevisiae, xylose is transported into the cells by hexose transporters, which have poor affinity for this pentose sugar. Thus, the expression of heterologous xylose transporters has been intensively studied and positive results have been obtained. 103
Lignocellulosic inhibitors and inhibition mechanisms
One of the primary barriers for the efficient conversion of lignocellulosic hydrolysates into ethanol is the presence of fermentation inhibitors. As shown in Figure 2, inhibitory compounds such as aliphatic acids, furan aldehydes, and phenolics are formed or released during biomass pretreatment and hydrolysis. 104 –106 These molecules affect an array of cellular processes in yeast cells, which must spend energy expelling or converting these molecules into less toxic compounds and repairing any incurred cell damage.
Aliphatic acids commonly found in lignocellulosic hydrolysates include acetic acid, levulinic acid, and formic acid, and their concentrations are dependent on pretreatment conditions. In their undissociated form, these weak acids are liposoluble and may permeate the yeast cell membrane. In the near-neutral pH of the cytosol, the acids dissociate, lowering the intracellular pH (pHi) and driving the cell to pump out protons through plasma membrane ATPases. This adenosine triphosphate (ATP) expenditure is exacerbated by the intracellular accumulation of anions, which inhibit the activity of glycolytic enzymes. 107 In an attempt to maintain intracellular ATP levels, the yeast cell increases ethanol production while diverting energy from cell growth. This metabolic shift may explain the ethanol yield and productivity increases observed in the presence of low concentrations of acetic acid. 50,99 However, the aliphatic acid content in industrial lignocellulosic hydrolysates typically surpasses these beneficial levels. Moreover, their inhibitory effect is influenced by other factors such as the overall medium composition and the fermentation pH. 105 The concentration of undissociated acid depends on the acid dissociation constant values (pKa, which are 4.76, 4.64, and 3.75 for acetic, levulinic, and formic acids, respectively) and on the pH of the medium. As the medium pH decreases, the concentration of the protonated acid and its toxicity to the yeast cell increase.
Furfural and HMF have been associated with low growth rates, reduced ethanol productivity, and DNA damage in yeast cells. 104,105 S. cerevisiae is able to convert both HMF and furfural into less toxic alcohols. However, these reduction reactions are catalyzed by NAD(P)H-dependent reductases. High concentrations of these inhibitors result in the depletion of the cofactors NADH and NADPH, which are needed for numerous reactions in the cell. Detoxification may also occur at the expense of ATP, possibly through their transport out of the cell by efflux pumps. 108 Although HMF and furfural have similar inhibition and detoxification mechanisms, HMF has been shown to be converted at a lower rate than furfural and to cause longer lag phases, likely due to the lower membrane permeability by HMF. 105
Lignin degradation during pretreatment processes releases various phenolic compounds that inhibit the fermentation of lignocellulosic hydrolysates. Phenolic inhibitors include 4-hydroxybenzoic acid, vanillin, catechol, ferulic acid, coumaric acid, and others. 50,99 Similar to other classes of inhibitors, the identity and concentration of phenolics found in a specific hydrolysate will depend on the feedstock used and on the pretreatment method applied. 105 Low molecular weight phenolic compounds have been found to be most toxic, and various mechanisms of inhibition have been described. These mechanisms include cell membrane damage, decrease in intracellular pH, translation inhibition, and DNA mutagenesis. 104
Several strategies have been successfully employed to improve yeast tolerance to lignocellulosic inhibitors. Evolutionary engineering in inhibitor-laden media is a commonly used approach. A number of genetic modifications have also been reported to increase inhibitor resistance, and naturally resistant isolates have been identified in high-throughput screening studies. 99,101,104 Nevertheless, the inhibitory effect of these compounds remains a challenge. This condition is especially true for the fermentation of the xylose present in lignocellulosic hydrolysates. Xylose fermentation produces less energy per sugar molecule consumed than glucose, making it more difficult for the yeast cell to cope with the inhibitors. This uneven effect demonstrated that the inhibition of xylose fermentation by acetic acid could be reversed by the addition of glucose. 109 Moreover, the economic viability of lignocellulosic ethanol production depends on the hydrolysis of biomass at high solids loading. This process generates hydrolysates with high concentrations of inhibitors, making the development of inhibitor-resistant yeast strains a crucial step for the success of the second-generation ethanol industry.
Brazilian Biodiversity Access and Risk Assessment
Brazilian wild-type yeasts are robust and productive microorganisms. However, the use of genetically modified wild-type yeast strains, such as a xylose-fermenting yeast, at commercial scale requires the approval of the Brazilian government regarding biodiversity access and risk assessment related to the use of GMOs.
Genetic resources have always been considered the heritage of humanity. Thus, these resources should be available as a source of raw materials for products that benefit all populations. However, in indiscriminately exploring the biodiversity of developing countries, including intellectual property rights, some institutions from industrialized countries have promoted biopiracy. To maintain sustainable use of Brazilian biodiversity, Law No. 13.123 of 2015 requests that a company declare the use of biodiversity through an electronic registration and share the benefits of products ready for marketing with the community. 110
The use of GMOs raises some concerns regarding potential health and environmental risks. To minimize risks in Brazil, the National Technical Biosafety Commission (CTNBio) is responsible for a rigorous risk assessment based on Normative Resolution number 5 of March 12, 2008. This resolution gives provisions regarding the rules for the commercial release of GMOs and their derivatives. 111 Briefly, the requesting company must submit to CTNBio a careful GMO risk-assessment study containing information regarding GMO risk classification, including details of the genes that have been introduced, such as their functions, original organisms, and methods used for genetic modification. Regarding environmental risks, the proposal must present information about spore production and resistance to desiccation; use of agents with sterilizing activity against the GMO; possible effects of the GMO on the quality of the water, air, and soil; and the GMO's ability to survive and disperse in the environment. GMOs consumed as food require that an assessment of risks to human and animal health be completed. Since 2009, CTNBio has completed the evaluation of at least five GMOs, all of which are related directly or indirectly to biofuels production. 112 In all cases, the decision of CTNBio was favorable to commercial approval.
Second-Generation Ethanol Plants Around the World
Most of the second-generation ethanol plants currently in operation began production in 2014, with three in the US and two in Brazil. Before 2014, one pilot plant was opened in Alpena, MI, in 2012, and the first commercial-scale plant was built in Crescentino, Italy, in 2013. Together, these plants are designed to produce approximately 435 million L/y of ethanol. The first commercial plant began production on October 10, 2013. Beta Renewables (Tortona, Italy), a subsidiary of Chemtex (Wilmington, NC), and Grupo M&G (Tortona), opened its cellulosic ethanol plant in Crescentino. It was the first plant designed to produce ethanol from agricultural residues such as rice straw, wheat straw, and Arundo donax (a high-yielding energy crop) at commercial scale using enzymatic conversion. This facility was designed to produce 75 million L/y of ethanol. Chemtex and M&G Group have invested in the development of a pretreatment and hydrolysis technology for transforming lignocellulosic material into sugars. This technology, called PROESA, can properly function with a wide range of feedstocks and includes an integrated solution for ethanol and power production. The hydrolysis process functions with an enzymatic cocktail supplied by Novozymes (Bagsvaerd, Denmark), and the fermentation step allows the conversion of C6 and C5 sugars into ethanol. A continuous pilot facility was constructed in Rivalta to validate this technology. 113
The POET (Sioux Falls, SD) and DSM (Herleen, The Netherlands) cellulosic ethanol plant, which is associated with a traditional corn ethanol plant in Emmetsburg, IA, started operation in September 2014. Called Project Liberty, it is designed to produce 94.5 million L/y of ethanol based on corn stover, but the intention is to expand its biomass consumption to corn cobs, leaves, husks, and some stalks. 114
Abengoa's (Seville, Spain) plant, which started operation in October 2014 in Hugoton, KS, was designed to produce 95 million L/y of ethanol. It uses corn stover residues and proprietary technologies to convert biomass to ethanol. These technologies include pretreatment processes and enzymatic hydrolysis. 115
Raízen (São Paulo), a joint venture between Royal Dutch Shell (Hague, The Netherlands) and the Brazilian firm Cosan (São Paulo), in 2015 inaugurated a cellulosic ethanol plant designed to produce 40 million L/y of ethanol. The plant uses Iogen Energy's cellulosic biofuel technology and is located adjacent to Raízen's Costa Pinto sugarcane mill in Piracicaba, São Paulo. Iogen provided Raízen with process technology and designs that were tested and proven at Iogen's demonstration facility in Ottawa, Canada. The Raízen facility, which is the second cellulosic ethanol plant in the Southern Hemisphere, will use sugarcane bagasse and straw as feedstock and cellulolytic enzymes provided by Novozymes. 116
The first cellulosic ethanol facility in the Southern Hemisphere was GranBio's plant, called Bioflex 1, which started operation in September 2014, in São Miguel dos Campos, Alagoas, Brazil (Fig. 4). At the time, it was the largest cellulosic ethanol facility in the world, designed to produce 82 million L/y of ethanol. GranBio's second-generation plant enables Brazilian ethanol production to be increased by 50% per ha due to the use of agricultural residues (straw and bagasse). The company is able to harvest and store 400,000 t/y of straw; in this case, cane fields do not need to be expanded. Additionally, GranBio, in partnership with Caeté, a first-generation ethanol plant from the group Carlos Lyra, located next to Bioflex 1, made a cogeneration system of steam and electricity that is fed by the byproducts of second-generation ethanol production, namely, sugarcane bagasse and lignin. Due to the biomass that is used and the production process, in 2014, the Air Resources Board in California confirmed GranBio's second-generation ethanol as the cleanest fuel produced at commercial scale in the world based on carbon intensity (7.55 g CO2e/MJ). The calculation considers factors such as raw material harvest, inputs and energy consumption, and transportation and distribution through a port in California. 117

Commercial plant for the production of second-generation ethanol. The Bioflex 1 unit from GranBio Investimentos SA was built in São Miguel dos Campos, Alagoas, Brazil, and started operation in early 2014 with a nominal production capacity of 82 million L/y. GranBio and Caeté, from the Carlos Lyra group, created a partnership for the integrated production of steam and electricity. Installed next to Bioflex 1, the co-generation system is fed by sugarcane bagasse and lignin—a byproduct of producing second-generation ethanol.
In October 2015, DuPont opened the world's largest cellulosic ethanol plant in Iowa, Nevada. The fully integrated plant uses corn stover as raw material and can produce almost 120 million L/y of ethanol, representing a huge reduction in greenhouse gas emissions compared to gasoline. In addition to the environmental advantages, the biorefinery will generate new jobs and new sources of revenue, since DuPont is collaborating with local farms that will provide 375,000 dry t/y of corn stover. 118,119
Conclusions
Bioethanol is an example of an efficient renewable fuel that has great potential to replace fossil fuels as is already happening in Brazil. Despite the high efficiency of first-generation ethanol production, the economic viability of this system is challenged by the high operational costs and limited land productivity. Second-generation ethanol has appeared as a promising, feasible, productive, and cheap alternative to improve alcohol production. The possibility of using the lignocellulosic matrix as a source of fermentable sugars for the production of fuels is a sustainable alternative. This has allowed the development of new technologies in all stages of the process, from the selection of feedstock for biomass production to the development of microorganisms capable of fermenting the sugars from this complex material. Despite the high complexity of technology for second-generation ethanol production, it is becoming a reality through the launch of the first industrial units focused on ethanol production from lignocellulosic material. The pioneers of this new technology are now producing the cleanest biofuel at commercial scale in the world, an important achievement in an era focused on global decarbonization policies and environmental concerns related to global warming.
Footnotes
Acknowledgments
The authors acknowledge Banco Nacional do Densenvolvimento (BNDES), Financiadora de Estudos e Projetos (FINEP), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Fundação de Amparo á Pesquisa do Estado de São Paulo (Fapesp) for financial support of ethanol fermentation research. We also thank Instituto Agronômico de Campinas (IAC) and RIDESA/UFAL for scientific collaboration.
Author Disclosure Statement
The authors are all employed or research associates at GranBio/BioCelere, Campinas, Brazil.