
Abstract
Butanol has been used as an important solvent in various industries, and is considered a good liquid fuel substitute for gasoline for use in internal combustion engines. Biobased production of butanol has a long history and was one of the first industrial-scale fermentation processes in which Clostridium strains were employed. Recent work on metabolic engineering of Clostridium strains has resulted in considerably improved production of butanol. In addition, several non-clostridial microorganisms have also been developed for butanol production. Here we review the advances in butanol production that have employed Clostridium strains and other microorganisms.
Introduction
Butanol has been used as an important industrial solvent and also as a building block for the production of various chemicals such as acrylate and methacrylate esters, glycol ethers, butyl acetate, and plasticizers. Recently, efforts to produce chemicals, fuels and materials from biobased resources have attracted attention as a means to address sustainability and environmental issues. Biobased production of butanol has attracted renewed interest because of the advantages of butanol over ethanol as a liquid transportation fuel; butanol has similar fuel characteristics to gasoline, which, unlike ethanol's high hygroscopicity and low energy density, allows it to be used as a fuel in an internal combustion engine without any engine modification.
Typically, butanol is biologically produced by solventogenic clostridia. Butanol production through acetone-butanol-ethanol (ABE) fermentation using Clostridium strains is one of the oldest large-scale industrial processes, going back more than 100 years. 1 Recently, several fermentation processes for the enhanced production of butanol have been reported by employing metabolically engineered Clostridium and Escherichia coli strains (Table 1). 2 –7 This paper reviews recent advances in the production of butanol using newly developed microbial strains. Future perspectives on the development of commercially competitive processes for butanol fermentation are also provided.
Butanol Production Using Selected Clostridium and E. coli Strains
Metabolic Engineering of Clostridia for Enhanced Production of Butanol
Genetic engineering of solventogenic clostridia is not easy, although some techniques such as a gene knockout strategy based on mobile group II intron have been reported (Fig. 1). Furthermore, the presence of complex metabolic regulatory mechanisms in solventogenic clostridia is one of the hurdles in designing and developing a high-performance butanol producer (Fig. 2). For example, regulatory mechanisms involved in the fermentation-phase shift during the typical two-phase (acidogenic followed by solventogenic) fermentation of Clostridium acetobutylicum, the flagship strain employed for ABE fermentation, are still not completely understood. Cells first produce acetic and butyric acids during the acidogenic phase of growth. Subsequently, production of butanol—as well as acetone and ethanol—occurs with reassimilation of acids from the medium during the solventogenic phase. Thus, engineering the pathways involved in acid formation and reassimilation is very important for butanol production in solventogenic clostridia.

Advances in clostridial gene knockout strategy by employing the mobile group II intron.
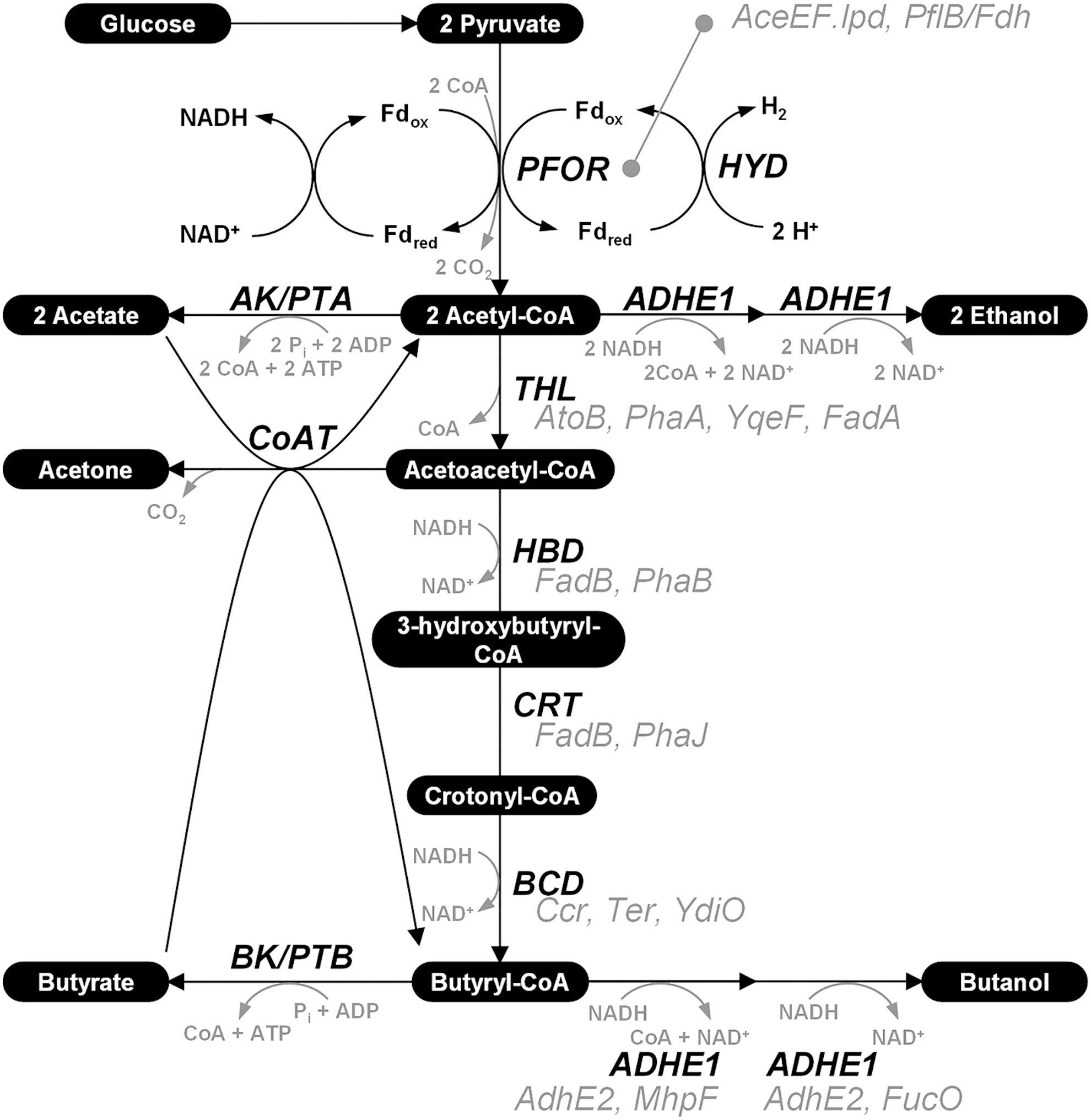
Key metabolic pathways of solventogenic clostridia. The inherent enzymes in the solventogenic clostridia are shown in black, while heterologous enzymes alternative to those of clostridia are shown in gray.
Phosphotransbutyrylase (PTB) and butyrate kinase (BK) encoded by the ptb and buk genes, respectively, are key enzymes for butyric acid production from butyryl-CoA in clostridia. These enzymes also form a pathway that competes with the butanol-production pathway. To improve butanol production, Green et al. constructed the buk-knockout mutant Clostridium acetobutylicum PJC4BK strain by using the non-replicative plasmid pJC4BK through homologous recombination. 8 By knocking out the buk gene, the BK activity in the PJC4BK strain was decreased to 19% of that in the wild-type strain. Butyric acid production decreased to 3.3 g/L, which is much lower than the 5.7 g/L produced by the wild-type strain in batch fermentation. 8 In another batch fermentation, the PJC4BK strain produced 16.7 g/L butanol. 9 Recently, the buk gene knockout has been revived and recharacterized to demonstrate the gene-knockout method based on the mobile group II intron Ll.ltrB in C. acetobutylicum (Fig. 1). The buk mutant C. acetobutylicum generated by employing the mobile group II intron also showed similar results in the production of butyric acid and butanol in batch fermentation. 2,10 More recently, the buk mutant Clostridium beijerinckii strain has also been developed. 11 However, the final titer of butanol obtained by the fermentation of the buk mutant C. beijerinckii has not reached the level achieved with the buk mutant C. acetobutylicum. In the batch fermentation, the buk mutant C. beijerinckii produced 12.7 g/L butanol from 60 g/L glucose, which corresponds to a 1.28-fold increase compared with the parent strain. 11
The ptb mutant C. acetobutylicum was constructed using the mobile group II intron only. 2,12,13 However, two different phenotypes were reported for the production of butyric acid in the batch fermentations of the ptb mutant C. acetobutylicum. Lehmann et al. reported the butyric acid negative phenotype, which is different from the results (about 3-5 g/L butyric acid production) obtained by two other research groups, although the reason is not clear. 2,12,13 In the production of butanol, the ptb mutant C. acetobutylicum showed varying butanol production of 0.5 g/L, 8.0 g/L, and 13.9 g/L from 40 g/L, 61 g/L, and 72 g/L glucose, respectively. 2,12,13 The first can be explained by the additional second site mutation, which caused a frameshift mutation in the thl gene encoding thiolase in the ptb mutant C. acetobutylicum. 13 Taken together, in the PTB/BK pathway, the buk gene is the primary target to be disrupted for improving butanol production in C. acetobutylicum, although the production of butyric acid could not be completely abolished.
In another work, C. acetobutylicum was engineered for the enhanced production of butanol by reinforcing the direct butanol-forming pathway through the knockout of the buk and pta genes together with overexpression of the adhE1D485G gene. 2 The engineered C. acetobutylicum strain produced 18.9 g/L butanol, with a yield of 0.29 g/g of glucose in batch fermentation. 2 In a fed-batch fermentation equipped with in situ recovery, the engineered strain produced 130 g/L butanol with a yield of 0.31 g/g of glucose. 2
More recently, the whole butanol-production pathway (from thiolase to aldehyde/alcohol dehydrogenase) has been overexpressed without engineering the acid-production pathway for enhanced production of butanol. 3 In this study, the E. coli gshAB genes were integrated at the C. acetobutylicum adc gene locus. 3 The resulting C. acetobutylicum strain produced 14.9 g/L butanol with a yield of 0.34 g/g of glucose. 3 In another recent study using C. acetobutylicum, butanol production increased by increasing the adenosine triphosphate (ATP) and nicotinamide adenine dinucleotide (NADH) levels through the overexpression of the pfkA and pykA genes encoding 6-phosphofructokinase and pyruvate kinase, respectively. 14 The resulting strain produced 19.1 g/L butanol with a yield of 0.21 g/g of glucose in a fed-batch fermentation. 14
Acidogenic Clostridium tyrobutyricum strain and an acetogen Clostridium ljungdahlii have also been engineered to produce butanol. The ack-deficient C. tyrobutyricum strain was engineered to produce butanol by introducing the C. acetobutylicum adhE2 gene on the basis of the plasmid pIMP1 that has the replicon of pIM13. 4 The resulting C. tyrobutyricum strain produced 16.4 g/L butanol with a yield of 0.30 g/g of mannitol (a more reduced substrate) in a fed-batch fermentation, while 10.0 g/L of butanol (yield of 0.27 g/g) was produced in a glucose-based fermentation. 4 In a later study, the replicon of pIM13 for expression of the adhE2 gene was replaced by the replicon of pBP1 (on plasmid pMTL82151), and introduced into the C. tyrobutyricum strain without any genome engineering. The resulting strain produced 20.5 g/L butanol with a yield of 0.33 g/g of mannitol in a fed-batch fermentation. 5
Butanol production was also demonstrated in a metabolically engineered C. ljungdahlii, which offers the potential of producing butanol from syngas through carbon fixation via the Wood-Ljungdahl pathway. 15 The whole C. acetobutylicum butanol-production pathway (from thiolase to aldehyde/alcohol dehydrogenase) was introduced into C. ljungdalhlii. During the mid-exponential growth phase, the resulting strain produced 0.15 g/L butanol from syngas in batch fermentation, while the butanol titer was less than 10% of the peak concentration at the end of fermentation. 15
Metabolic Engineering of E. Coli for Butanol Production
E. coli has also been metabolically engineered for butanol production. Introduction of the whole C. acetobutylicum butanol-production pathway in E. coli has not resulted in significant butanol production. Butanol production of less than 1.2 g/L was obtained with a yield of less than 0.03 g/g in glucose-based fermentation. A low butanol yield has also been observed when similarly engineered Bacillus subtilis, Pseudomonas putida, Lactobacillus brevis, and Saccharomyces cerevisiae were employed. 16 –20 The major bottleneck in butanol production by these host strains has been identified as the weak activity of butyryl-CoA dehydrogenase encoded by the bcd gene, which could be overcome by employing the Treponema denticola ter gene encoding trans-enoyl-CoA reductase instead of the bcd gene. 6,21
In metabolic engineering of E. coli for butanol production, the genes responsible for byproducts formation—frdABCD for succinic acid, ldhA for lactic acid, and adhE for ethanol—were knocked out to provide more carbon flux toward butanol production. 6 In addition, the synthetic butanol pathway was constructed by the overexpression of the T. denticola ter, E. coli atoB, and C. acetobutylicum hbd, crt, and adhE2 genes. 6 Also, additional NADH was supplied from formate by the heterologous enzyme encoded by the Candida boidinii fdh gene. 6 The final engineered E. coli strain produced 15 g/L butanol, with a yield of 0.36 g/g of glucose in an anaerobic flask culture, while 30 g/L butanol with a yield of 0.29 g/g was obtained in an anaerobic fed-batch fermentation equipped with gas-stripping. 6 Another interesting strategy has been developed for the production of butanol by E. coli through the use of a reversed beta-oxidation pathway, which is similar to the clostridia butanol-production pathway in the reaction sequences. 7 The metabolically engineered E. coli produced 14.0 g/L with a yield of 0.33 g/g of glucose through the reversed beta-oxidation pathway in a baffled shake-flask culture. 7
Development of Fermentation Processes
One of the major problems negatively affecting industrial-scale butanol production is the low butanol titer, caused by butanol toxicity to cells. To overcome this problem, various bioprocesses, including batch, fed-batch, continuous fermentation, and integrated fermentation with in situ recovery have been studied. Batch and fed-batch fermentations offer simple operation, which reduces contamination risk. Recently, pilot-scale (200L) batch fermentation was demonstrated using the engineered C. acetobutylicum BKM19 (pIPA100) strain for butanol-ethanol-isopropanol production. The engineered C. acetobutylicum BKM19 (pIPA100) strain produced 15.4 g/L butanol, 9.6 g/L ethanol, and 3.5 g/L isopropanol from 76.0 g/L glucose. 22
The integrated fermentation process with in situ recovery has been developed to improve butanol production by decreasing butanol toxicity. Among several in situ recovery techniques, adsorption and gas stripping have recently been applied to butanol fermentation using a metabolically engineered strain. For example, fed-batch fermentation of the engineered C. acetobutylicum BEKW (pPthlAAD**) strain was performed in an integrated process having a recovery column packed with sephabeads (Shimadzu, Tokyo, Japan) as a butanol adsorbent. 2 Culture broth was circulated through the sephabeads column when the butanol concentration in the fermentor reached 7 g/L. The operation of the integrated fed-batch fermentation with in situ recovery was performed with recycling of the sephabeads every 4–8 h. 2 In this fed-batch process, the C. acetobutylicum BEKW (pPthlAAD**) strain produced 585.3 g of butanol from 1,861.9 g of glucose in 111 h, which resulted in a yield of 0.31 g/g and a butanol productivity of 1.32 g/L/h (Table 1). 2
Gas stripping has also been successfully applied to enhance butanol production by the metabolically engineered C. acetobutylicum and E. coli strains. 23 The C. acetobutylicum PJC4BK (pIPA3-Cm2) strain produced 26 g/L butanol during the fed-batch fermentation, with gas stripping in 45 h. 23 The engineered E. coli strain produced 30 g/L butanol with a yield of 0.29 g/g of glucose in an anaerobic fed-batch fermentation with gas stripping. 6 More details on recovery techniques, such as adsorption, liquid-liquid extraction, perstraction, reverse osmosis, pervaporation, and gas stripping can be found in previous review papers. 24,25 These in situ recovery processes will lead to a reduction in overall cost for fermentative butanol production.
Compared to batch or fed-batch fermentation, continuous fermentation has the advantages of increasing productivity and reducing the preparation time and lag phase. Furthermore, membrane cell recycling bioreactors (MCRB) have additional advantages, such as allowing culture broth homogeneity and achieving high cell density in the bioreactor. Thus, MCRB can be applied to achieve significant improvement in the volumetric productivity of butanol. Recently, two engineered clostridial strains were evaluated using MCBR. Malaviya et al. reported butanol production from glycerol using the mutant Clostridium pasteurianum strain. 26 The MCRB fermentation was operated for 710 h, and the maximum volumetric productivity of butanol was 7.8 g/L/h. 26 More recently, butanol has been produced with volumetric productivities of 10.7 g/L/h by MCRB fermentation of the mutant C. acetobutylicum BKM19 strain. 27
Conclusions
Butanol has received renewed interest as an effective substitute for gasoline, in addition to its continued use as an important industrial solvent. To make biobased butanol a cost-competitive transportation fuel, the overall costs of production need to be reduced by developing a high performing strain capable of delivering butanol at high titer, yield, and productivity from inexpensive substrates with increased butanol tolerance. The development of efficient fermentation and downstream processes are also necessary. Non-food biomass or, even more preferably, waste biomass should be used as a substrate. Beyond traditional substrates, syngas fermentation has recently been attracting attention. Syngas can be produced from non-food biomass as well as natural gas.
Although metabolically engineered C. acetobutylicum and E. coli strains can produce butanol with high yield from glucose, the butanol titer is capped at around 20 g/L due to butanol toxicity. As mentioned previously, this is one of the bottlenecks in industrial-scale butanol fermentation. Although integrated bioprocesses that couple fermentation and in situ recovery can be a solution, it is still a much better option to have butanol-tolerant strains. Systematic approaches are needed to improve butanol tolerance of the microorganism. As a recent example, a systems-level computational framework was developed for clostridial fermentation that combines metabolic reactions, gene regulation, and environmental cues. 28 Such a systematic framework helps our understanding of complex clostridial metabolism and physiology, which can be extended to uncover the mechanisms of butanol tolerance in clostridia in the future.
Engineering the enzymes involved in the butanol-forming pathway can further improve butanol production. In a recent study, C. acetobutylicum thiolase was engineered for reduced susceptibility toward its inhibitor CoASH through random mutant library screening. 29 The C. acetobutylicum ATCC 824 expressing the mutant thiolaseR133G/H156N/G222V showed an 18% increase in butanol production compared with the control strain. 29 In a more recent study, another non-redox regulated mutant thiolaseV77Q/N153Y/A286K incapable of forming intermolecular disulfide bonds showed a higher enzyme activity than the wild-type enzyme. 30
The crystal structures of several clostridial enzymes involved in the central metabolic pathway were recently described, including C. acetobutylicum crotonase and Clostridium butyricum (S)-3-hydroxybutyryl-CoA dehydrogenase. 31,32 Redesign of these enzymes on the basis of the three-dimensional structures of the wild-type enzymes can lead to the development of better enzymes for butanol production. With all of these advances, it is expected that industrial-scale fermentative production of butanol from various carbon sources will be realized again.
Footnotes
Acknowledgments
This work was supported by the C1 Gas Refinery Project from the Ministry of Science, ICT, and Future Planning through the National Research Foundation of Korea.
Author Disclosure Statement
No competing financial interests exist.