
Abstract
The plant Baliospermum montanum was found to produce hydroxynitrile lyases (HNL) during a screen for the enzyme among fruits in markets and plant specimens obtained from a botanical garden. The enzymatic properties of the S-stereoselective HNL have been characterized using purified enzyme from the leaves of B. montanum and an Escherichia coli transformant expressing the enzyme. We determined the structure of BmHNL by X-ray crystallography and successfully engineered BmHNL to have H103C and N156G double mutants on the basis of its structure, molecular dynamics simulation with the substrate, and speculation on the reaction mechanism. The specific activity of the BmHNL H103C/N156G mutant was improved to 154 U/mg from 52 U/mg as compared with BmHNL wild type for (S)-mandelonitrile production, and the enantiomeric excess of (S)-mandelonitrile produced by the double mutant was increased to 93% from 55% compared with the BmHNL wild type. The double mutant is suitable for (S)-mandelonitrile synthesis, improving its enantioselectivity and specific activity.
Introduction
We demonstrated that there is a pathway in microorganisms in which aldoxime is metabolized to nitrile, which is then hydrolyzed to carboxylic acid. The dehydration of nitrile is catalyzed by aldoxime dehydratase, a newly discovered heme-dependent enzyme. The hydrolysis of nitrile is catalyzed by a combination of nitrile hydratase and amidase or by a single enzyme nitrilase. 1,2 A similar pathway in plants was indicated by Halkier and Møller et al. in Sorghum bicolor synthesizing cyanohydrin dhurrin from L-Tyr via aldoxime with two cytochrome P450s. 3 Different from the microbial aldoxime dehydratase, whose product is solely a nitrile from aldoxime, plant P450 catalyzes the synthesis of hydroxynitrile or nitrile from aldoxime. 4 Cyanohydrins are synthesized from hydrophobic amino acids (L-Tyr, L-Phe, L-Val, L-Ile, and L-Leu). 5,6 We demonstrated that (R)-mandelonitrile is synthesized from L-Phe via aldoxime with two cytochrome P450s in Prunus mume. 7 Cyanohydrins are stored as glycosides, which are hydrolyzed back to cyanohydrins by β-glycosidase in response to physical damage, followed by the production of hydrogen cyanide and aldehydes by the action of hydroxynitrile lyases (HNL) or by non-enzymatic reactions. It is reported that cyanogenic plants are widely distributed among 2650 to 3000 species, 5 and cyanogenesis from cyanohydrin by HNL in these plants has evolved to protect the plants from attack by insects, microorganisms, etc.
HNL catalyzes the stereoselective synthesis of chiral cyanohydrin from aldehyde and hydrogen cyanide (HCN) with a reverse reaction of its physiological use, and it is currently utilized by industry to produce chiral cyanohydrins (Fig. 1). 5,8 The history of applying plant HNL for organic synthesis has been very different from that of microbial enzymes; it has taken more than 100 years for the application of HNL in the industrial production of chiral cyanohydrins to be established since its discovery in Prunus dulcis (amygdalus). 5 This contrasts with recent developments in the application of microbial enzymes, in which screening for the enzyme is carried out in a shorter period followed by industrial development. 9 Liebig and Böhler performed the initial research on HNL, and observed the production of HCN from cyanohydrin glycoside amygdalin with an extract (emulsin) from almond, which should have contained β-glucosidase and HNL with a current terminology. 10 Effenberger et al. and Griengl et al. used R-selective HNL (R-HNL) from P. dulcis (amygdalus) and S-selective HNL (S-HNL) from Hevea brasiliensis (HbHNL) for the synthesis of chiral cyanohydrins. 11,12
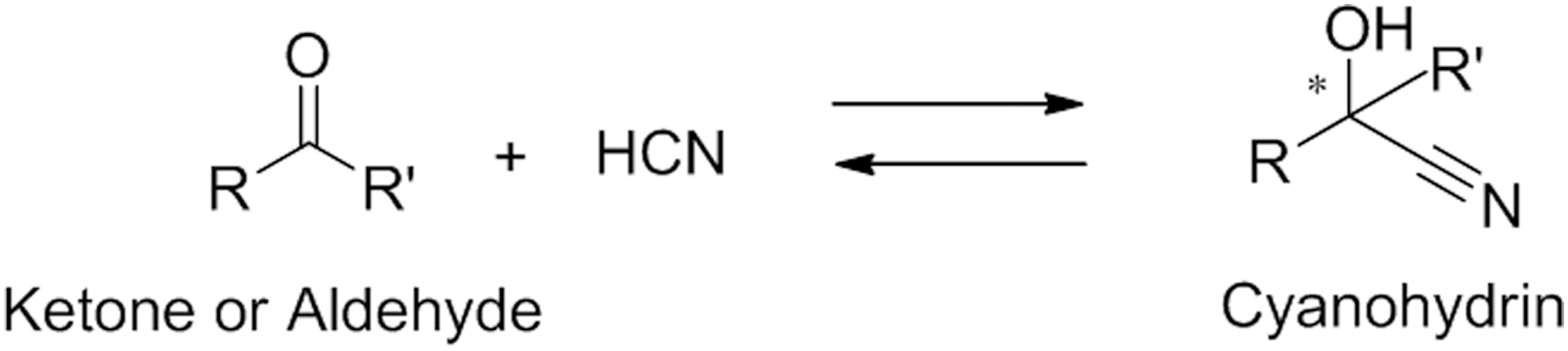
HNL catalyzes the stereoselective synthesis of chiral cyanohydrin from aldehyde and hydrogen cyanide with a reverse reaction of its physiological use.
In the current research, we summarize how we discovered a new distribution of S-HNL from Baliospermum montanum (BmHNL) and improved the properties of the enzyme by X-ray crystallography, molecular dynamics analysis, and protein engineering.
Screening for New HNLS from Plants
We endeavored to screen plants as a new enzyme source using the same methodology used to screen many microorganisms for new enzymes and reactions. 1 We were prompted to screen for new HNLs among plants, because there were limited reports on HNLs from cyanogenic plants and few enzymatic studies on HNLs.
High-performance liquid chromatography (HPLC) with a chiral column was useful to screen for new HNLs, because the crude extract of plants is often deeply colored and has viscosity caused by polysaccharides. Therefore, employing an ultraviolet spectrophotometric assay at 249 nm to determine the decomposition of cyanohydrins was ineffective. We established a method to detect the formation of chiral mandelonitrile directly from benzaldehyde and potassium cyanide using HPLC, which is more sensitive and shows the stereochemistry of cyanohydrins simultaneously. We detected R- and S-HNL activities plant homogenates of leaves, roots, shoots, spikes, and rhizomes from 163 species of plants among 74 cyanogenic and non-cyanogenic plant families from fruits obtained in the market and plant samples from the Botanic Gardens of Toyama. 13 The homogenates of 156 species were inactive, and new sources of HNLs were identified, including leaves and seeds of R-HNL producers Passiflora edulis, Eriobotrya japonica, P. mume, Chaenomeles sinensis, Sorbus aucuparia, and leaves of Passiflora edulis. Of these enzymes, HNLs from E. japonica and P. mume contained flavin adenine dinucleotide similar to almond HNL and catalyzed the R-selective synthesis of cyanohydrin from aromatic aldehydes and aliphatic ketones. 4,14 –17 R-HNL from P. edulis is a new type of HNL because the enzyme is a monomer and its molecular mass is the smallest among the know HNLs (18 kDa). 10 B. montanum leaves were identified as a new S-HNL source (Table 1). 12,13 B. montanum had been transplanted to the Botanic Gardens of Toyama, Japan from the Kunming Botanical Garden, Yunnan Province, China. 17
HNL Activity Shown by the Homogenates of Various Plants 12
The homogenate of kernels from Prunus dulcis showed 73.5 units/mg of (R)-HNL activity under the same condition.
c DNA Cloning and the Enzymatic Properties of S-B
m
HNL
18
BmHNL was purified and the N-terminal amino acid sequence was analyzed for cDNA cloning. The hnl cDNA was cloned by rapid amplification of cDNA ends with a designed primer based on the N-terminal amino acid sequence. BmHNL's amino acid sequence was similar to those of HbHNL (56%) and HNL from Manihot esculenta (MeHNL) (55%) (Fig. 2). 20 –23 The use of regular vectors such as pUC and pET in the expression of BmHNL in Escherichia coli resulted in the production of an insoluble enzyme. To improve expression, we optimized the expression condition in E. coli with pColdI, and succeeded in the functional expression of BmHNL at low temperature. BmHNL was the third enzyme among S-HNLs expressed as soluble enzymes in E. coli. The estimated molecular mass of a subunit in BmHNL, which was obtained via sodium dodecyl sulfate polyacrylamide gel electrophoresis, is about 29.5 kDa. The dimer structure was indicated by gel filtration (molecular mass of 60–65 kDa). The substrate specificity of BmHNL was analyzed with approximately 100 carbonyl compounds (65 aromatic and 35 aliphatic). The reaction rates for the aliphatic aldehydes and ketones were lower than those for aromatics, and some aromatic aldehydes and ketones were preferred substrates compared with aliphatic substrates for BmHNL. Among the aromatic substrates, the enzyme exhibited the highest kcat /Km (catalytic efficiency) for piperonal.

Alignment of amino acid sequence of α/β-hydrolase HNL. 20 These are sequence alignment of α/β-hydrolase HNLs from Manihot esculenta (MeHNL), Hevea brasiliensis (HbHNL), Baliospermum montanum (BmHNL) and Arabidopsis thaliana (AtHNL). 21 –23 The sequence of BmHNL showed 56, 55, and 46% identity to HbHNL, MeHNL and AtHNL, respectively.
Structure Determination of B
m
HNL by X-Ray Crystallography and Analysis by Molecular Dynamics
24
We determined two different structures of BmHNL, apo1 and apo2, at 2.55 and 1.9 Å, respectively. The overall structure of BmHNL was similar to other S-HNLs (MeHNL and HbHNL). However, the substrate specificity of BmHNL was different from other S-HNLs; S-HNLs preferred substrate was acetone cyanohydrin while BmHNL showed higher affinity for (S)-mandelonitrile, which has a lower K m value. To explain the substrate preference, we compared the structure of BmHNL with other S-HNLs. In the active site of BmHNL, Ser80-Asp207-His235 and Lys236 were conserved as catalytic residues and Lys236 for the recognition of cyanide. We speculated that the reaction mechanisms of BmHNL would be the same as S-HNLs from MeHNL and HbHNL. Some specific amino acid residues were conserved around the active site and entrance region of BmHNL. Table 2 shows that F121, F178, T152, and I157 are specific residues located at the entrance region in BmHNL. 20 All of these amino acids are altered to only Leu in MeHNL and HbHNL. One found that the substrate specificity of MeHNL was improved for the synthesis of long chain cyanohydrin by the mutant W128A, and postulated that the entrance region would be important for the substrate specificity in S-HNLs. 25,26 We also speculated that F121, F178, and W128 conserved in the entrance region of BmHNL would play roles in the recognition of the benzene ring of aromatic substrates by π–π stacking interaction.
The Hydrophobic Residues Located in the Active Site of the Three S-HNLs
Amino acid number according to BmHNL. Under bar shows the residues that locate at the entrance region. Gly81, Phe121, Phe133, Thr152, Ile157, Phe178 were specific residues in BmHNL.
F178 was found to be a flexible residue because it showed different forms with directional change in the structures of apo1 and apo2 (Fig. 3B). 24 To analyze the flexibility of these hydrophobic amino acids in the entrance region, molecular dynamics analysis was carried out on the structure of BmHNL (apo2). The results showed that F121 and W128 are clearly flexible residues, not only in the side chain but also along the backbone. F178 was stable only if a side chain was changed by flipping the state of the backbone (Fig. 3C). These results suggested that BmHNL has a flexible structure in solvents and can change the size and conformation of the active site or the entrance region via contributions by these flexible residues.
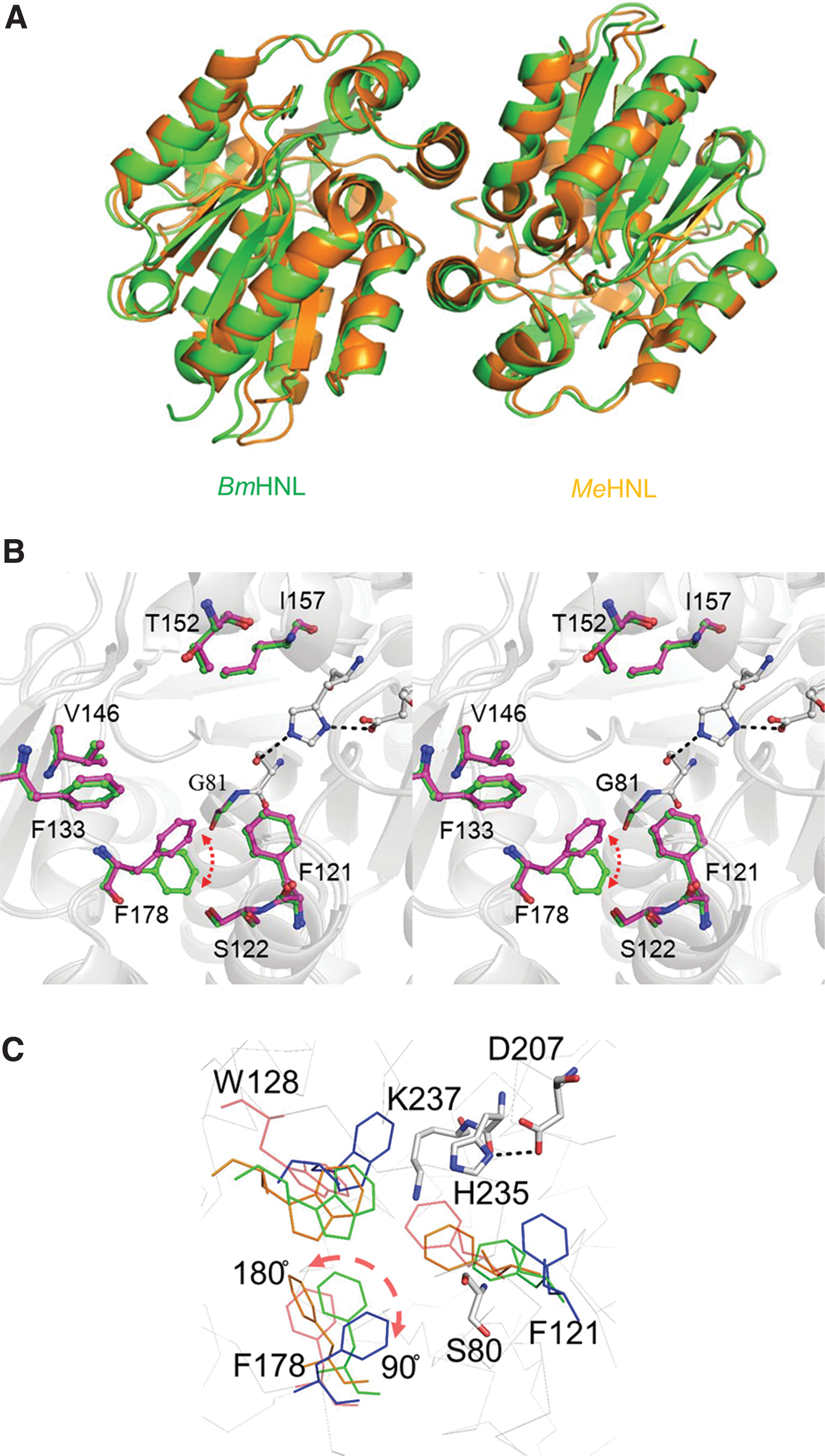
Structure and molecular dynamics of BmHNL.
24
Improvement of Bm
HNL by Protein Engineering: Soluble Expression, Docking Simulation, and Mutation
20
High specific activity, enantioselectivity, and temperature and pH stability would be required of HNLs used in the industrial production of chiral cyanohydrins. S-Selective BmHNL has the potential to be used in industrial cyanohydrin production because it showed broad substrate specificity toward aromatic aldehydes and ketones as well as higher temperature stability than other S-HNLs. 19 However, the enzyme has some drawbacks, as its enantioselectivity and the specific activity are lower than those of other S-HNLs. To expand the application of BmHNL for the production of various (S)-cyanohydrins, we improved the enantioselectivity and specific activity of BmHNL by screening highly active mutants from the random mutagenesis library and saturation mutagenesis at N156 of BmHNL H103C mutant.
Screening of the Highly Active Mutants from the Random Mutagenesis Library
In the first screening, a random mutant library was constructed with error-prone polymerase chain reaction (PCR) using pUC19-BmHNL wild type as a template. Highly active mutants, however, were not obtained from the library. To improve the soluble expression of BmHNL, we successfully introduced the H103C mutation to BmHNL at the same site with MeHNL, based on our prior discovery that the mutation in MeHNL caused the drastically soluble expression of the enzyme at 37 oC in E. coli. 20 We further investigated the enzyme with random mutagenesis by PCR using pUC19-BmHNLH103C as a template, and obtained some improved mutants (H103C/N156D, H103C/T152I, H103C/I157V) with higher activity. Since the mutation included specific hydrophobic residues of BmHNL in the active site at N156, and the mutation was most effective in increasing activity, a docking simulation of BmHNL with the substrate (S)-mandelonitrile was carried out using the molecular operating environment program. The results showed that the N156 residue could not directly interact with (S)-mandelonitrile because the N156 residue is not located within 5 Å of (S)-mandelonitrile (>8.0 Å) (Fig. 4) in the predicted structure of BmHNL complexed with (S)-mandelonitrile. 20
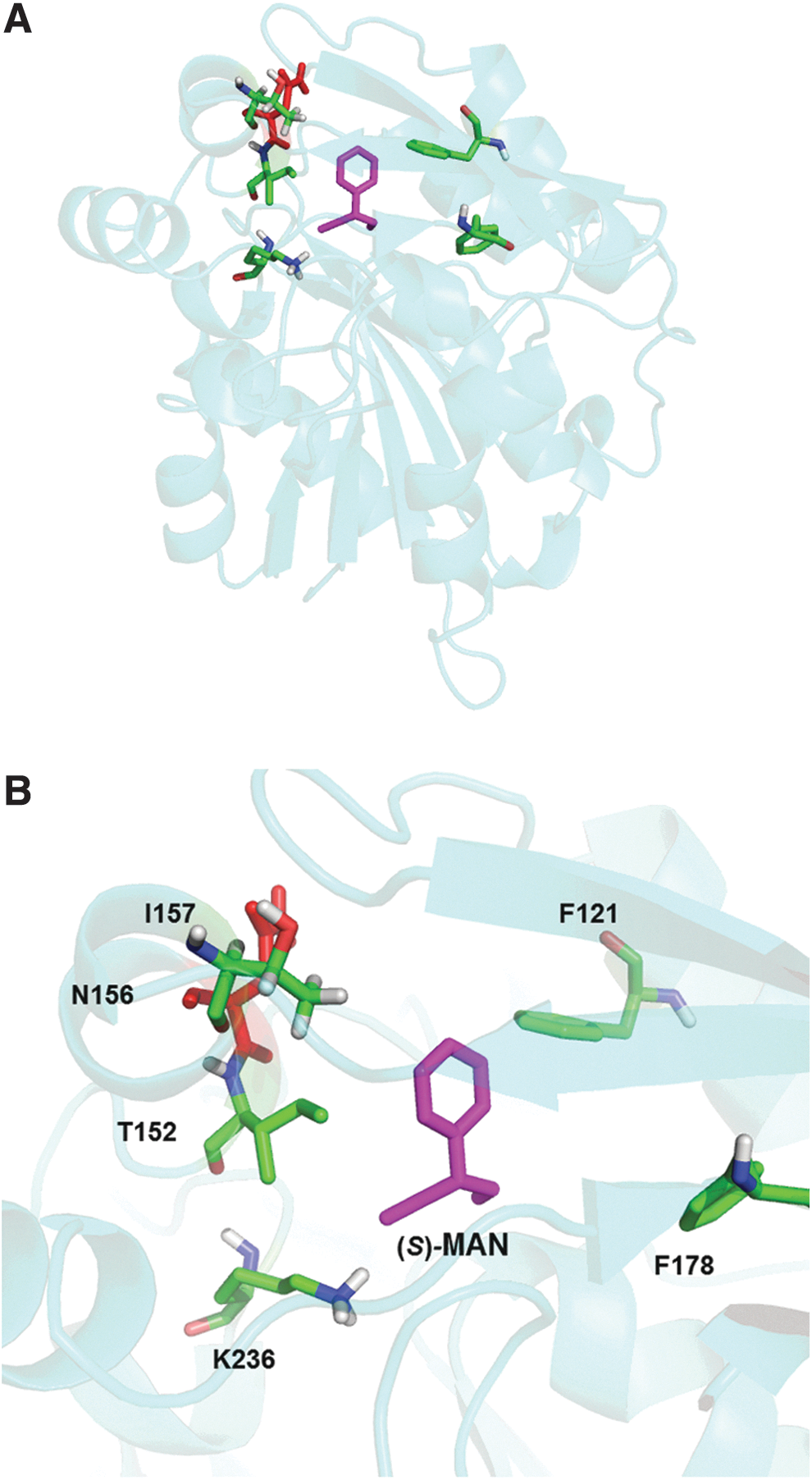
Docking simulation of BmHNL with (S)-mandelonitrile.
20
Overview of the predicted structure of BmHNL with (S)-mandelonitrile by docking simulation;
Saturation Mutagenesis at N156 of B
m
HNL H103C Mutant
The N156 mutation was further investigated in detail because the specific activity of H103C/N156D was higher than that of the other highly active mutants (H103C, H103C/I157V, and H103C/T152I), even though N156 residue does not directly interact with the substrate (S)-mandelonitrile. By replacing N156 with each of the 19 amino acids by sauration mutagenesis, we found that the H103C/N156G mutant showed the highest specific activity among all the variants. The specific activities for (S)-mandelonitrile production of the purified H103C/N156D and H103C/N156G mutants showed 117 and 154 U/mg of protein, respectively, which were 5- to 7-fold higher than that of the H103C mutant (22.8 U/mg of protein).
Evaluation of Improved Characteristics and Synthesis of (S)-Mandelonitrile
Enantioselectivity of Mutated B
m
HNLs (H103C, H103C/N156D, H103C/N156G) to Evaluate the Improved Enantioselectivity for (S)-mandelonitrile Synthesis by the Mutation at N156
Under the original reaction conditions, (S)-mandelonitrile formed by three mutants (H103C, H103C/N156D, H103C/N156G) was compared by determining the reaction velocity of total mandelonitrile production ((R)-mandelonitrile plus (S)-mandelonitrile). The enantiomeric excess (ee) values of (S)-mandelonitrile produced by the catalysis of H103C/N156D and H103C/N156G mutants were higher than that of the H103C mutant (Fig. 5). 20 The ee of (S)-mandelonitrile produced by the H103C/N156G mutant increased up to 93% by adding more enzyme, as compared with the wild type enzyme showing the maximum ee of 55% in the same condition.
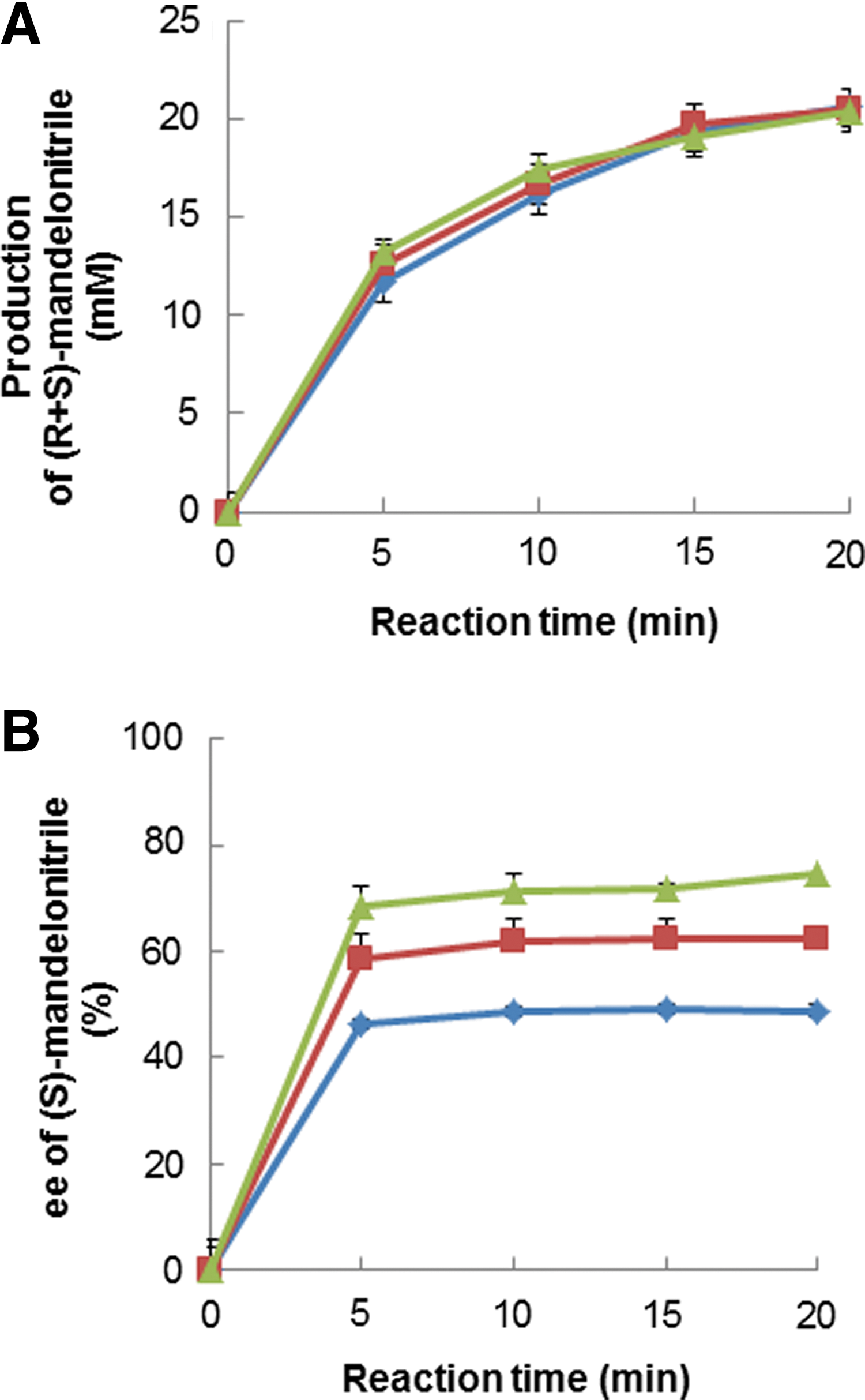
Enantioselectivity for (S)-mandelonitrile of purified enzymes from BmHNL variants.
18
Mandelonitrile production was carried out under standard assay conditions using 0.65 U of each enzyme and the amount were determined by HPLC;
Kinetic Analysis of Mutated B
m
HNLs
In the kinetic analysis of BmHNL mutants, the catalytic efficiency shown by the H103C/N156G mutant (double mutant) for (S)-mandelonitrile was improved to 251/mM/s from 157/mM/s for the H103C mutant, while that of the (R)-mandelonitrile by the double mutant decreased to 8.8/mM/s from 29.7/mM/s, accompanying the increase in the Km value for (R)-mandelonitrile caused by mutation at N156 (Table 3). The E-value of the double mutant for (S)-mandelonitrile also improved to 28.5 from 5.38 of the H103C mutant, with 5.3-fold improvement from that of the H103C mutant. As mentioned above, the N156 residue of BmHNL cannot directly interact with (S)-mandelonitrile and is located in the α-helix structure on the surface of BmHNL. However, the mutation at N156 improved the catalytic efficiency and the ee of (S)-mandelonitrile. We therefore presume that conformational change of the α-helix structure including some effective residues (T152, I157, etc) for HNL activity would have resulted from N156 mutation in BmHNL.
Kinetic Parameters of BmHNL Mutants with (R)- or (S)-Mandelonitrile 20
The kinetic parameters were determined along with the degradation activity of (R)- or (S)-mandelonitrile by detecting kinetic slope at 280 nm. k cat = (V max)/(60 × 29 kDa molecular mass of the subunit), E-value = ((k cat/K m of (S)-mandelonitrile)/(kcat /Km of (R)-mandelonitrile)).
Conclusions
Six R-HNLs, including the novel type HNL from P. edulis and an S-HNL from the leaves of B. montanum, were identified as new sources of HNLs in a screening of 163 plant species belonging to 74 cyanogenic and non-cyanogenic plant families. The amino acid sequence of BmHNL showed a high similarity to other S-HNLs, and BmHNL was the third enzyme among S-HNLs expressed as a soluble enzyme in E. coli. BmHNL showed broad substrate specificity toward aromatic compounds, albeit with lower enantioselectivity and specific activity compared with other S-HNLs. In the structure and molecular dynamics analysis, some hydrophobic residues (F121, F178, and W128) showed flexible form and flipping of the side chain in the BmHNL structure. The broad substrate specificity shown by BmHNL toward aromatic compounds is attributed to its flexible structure in solvents, the fact that it can change the size and conformation of active site, and the presence of flexible residues at the entrance region. We further improved the enantioselectivity and specific activity for (S)-mandelonitrile synthesis by random mutagenesis and saturation mutagenesis. We have been successful in engineering BmHNL to have H103C and N156G double mutants, which is suitable for (S)-mandelonitrile synthesis, improving its enantioselectivity and specific activity.
Footnotes
Acknowledgments
We thank Dr. Shogo Nakano, currently at Shizuoka Prefectural University (Shizuoka, Japan) for X-ray crystallography and molecular dynamics studies on HNL from B. montanum when he belonged to the Exploratory Research for Advanced Technology (ERATO) Asano Active Enzyme Molecule Project.
Dedicated to Romas Kazlauskas on the occasion of his 60th birthday.
Author Disclosure Statement
No competing financial interests exist.