
Abstract
Lactococcus lactis subspecies lactis ATCC 11454 is a natural nisin A-producing strain. It was used as the parent strain for adaptive mutations in this study to screen for high-yield nisin-producing mutants. Through a series of classical mutations with physical and chemical mutagenic agents, including ultraviolet, LiCl, and diethylsulfate (DES), followed by adaptive mutation in a nutrient medium containing high concentrations of nisin, two high-yield nisin-producing mutant isolates 3807 and 3823 were obtained. In shake flask fermentation experiments, 3807 and 3823 strains produced nisin at 2,160 IU/mL and 2,140 IU/mL, respectively, in the fermented broth—more than 3 times higher than the parent strain ATCC 11454. These mutants were relatively stable over sub-culturing for 3 passages. In a 10-L fermentor, the mutant 3807 produced an average of 11,870 IU/mL of nisin. At 700-L fermentation scale, the average yield was 10,350 IU/mL. When it was scaled up to 50-ton fermentation, it yielded an average of 9,960 IU/mL of nisin A in the fermented medium.
Introduction
Nisin is a natural antimicrobial peptide. It is the only bacteriocin approved by United States FDA as a Generally Recognized As Safe compound for use in foods to control foodborne pathogens and spoilage organisms. Nisin is bactericidal primarily effective against the growth of Gram-positive bacteria such as Micrococcus, Streptococcus, Lactobacillus, Staphylococcus, Listeria, and Bacillus species. It also inhibits the germination of bacterial endospores such as Clostridium, Alicyclobacillus, and Bacillus spores. 1 Nisin is produced by Lactococcus lactis through fermentation and is considered as a natural antimicrobial ingredient. It has thus increasingly gained popularity in recent years in the food industry. 1 –3 Today it has been widely used in more than 50 countries as a safe and natural antibacterial food preservative. 4 Nisin manufacturers and researchers have been working to improve the yield of nisin by isolating nisin-producing strains; 5,6 optimization of fermentation media; 7 –16 and fermentation conditions. 16 –19
The nisin biosynthesis and nisin-induced expression system in Lactococcus lactis have been well studied. 20 –22 Some researchers have made modifications on the nisin-producing strains by increasing copies of plasmids or genes involved in nisin biosynthesis in L. lactis, and the nisin yield was significantly increased. 23,24 Hu et al. also successfully cloned a nisin immunity gene nisl with a strong promoter P59 into a vector and then introduced it into Lactococcus lactis NZ9800 to create a recombinant strain that had 25% higher nisin resistance and produced 32% more nisin during fermentation. 25 A similar approach was utilized by Kim et al. 26 Field et al. even obtained a bioengineered nisin A derivatives with an enhanced antimicrobial activity against both Gram-positive and Gram-negative pathogens. 4 However, such genetically modified organisms (GMOs) have not been widely accepted in commercial production, partly because of the GMO safety concerns to both consumers and regulators.
In this study, an adaptive mutation process was employed as a novel breeding method to generate high levels of nisin-producing and -resistant mutant strains of L. lactis. To do this, a nisin-containing medium was used to screen nisin-resistant mutants after physical and chemical mutations of a natural nisin-producing strain. The resulting high nisin-producing mutant would be more acceptable for commercial nisin production.
Materials and Methods
Culture and Media
The starting parent strain for mutation studies was L. lactis ATCC11454, a natural nisin A-producing strain.
The liquid medium for growing the cultures and fermentation contained 1% sucrose, 1% yeast extract, 1% peptone, 2% KH2PO4, 0.2% NaCl, and 0.02% MgSO4.7H2O. The agar plates and slants were made with the same medium plus additional 2% agar. Cultures were grown at 30°C for 24 h and maintained on the agar slants at 5°C.
Nisin Assay
The antimicrobial activity of nisin was determined by using a modified agar well diffusion bioassay, 27 –29 using Micrococcus luteus NCIB8166 as the indicator strain. The indicator strain was grown at 30°C for 18–20 h and was inoculated into a pre-sterilized and tempered (in a 47°C water bath) brain heart infusion (BHI) agar medium at 0.1% of inoculation rate. After mixing well, 20 mL of the medium was poured into each Petri dish to make the agar plates for bioassay. The plates were placed on a flat bench for solidification at room temperature. After the plates were solidified, 6–7 wells (6.0 mm in diameter each) on each plate were made with a sterile stainless steel cork bore. For each well, 40 μL of sample solution was filled in. The plates were then incubated at 30°C for 18–20 h, and the diameter of inhibition zones was measured with a digital caliper. The activity was then calculated according to the standard curve obtained with nisin standard solutions in the same assay. The nisin standard solutions were prepared by dissolving a certain amount of nisin standard powder in 50 mM KCl–HCl, pH 2.0 buffer. The nisin standard powder containing 1000 IU/mg of nisin A was purchased from Sigma Aldrich (St. Louis, MO). Typically, four nisin standard solutions were used to obtain a standard curve. The standard solutions of 200 IU/mL, 500 IU/mL, 1,000 IU/mL, and 3,000 IU/mL were prepared by dissolving 20 mg, 50 mg, 100 mg, and 300 mg of nisin standard powder in 100 mL pH 2.0 buffer, respectively. The standard solutions were kept at 5°C for repeated use for up to 3 months. The standard curve is a linear straight line between the diameter of the inhibition zone and the log (nisin concentration) when the nisin concentration is in the range of 100–3,000 IU/mL. Nisin standard solutions were assayed along with the samples on the same plates under the same conditions, and standard curve was generated for calculation in each test. Minimally 4 replications were conducted in each assay, and the average diameter of inhibition zones was used in calculation. Figure 1 is a typical nisin standard curve obtained in the assay.
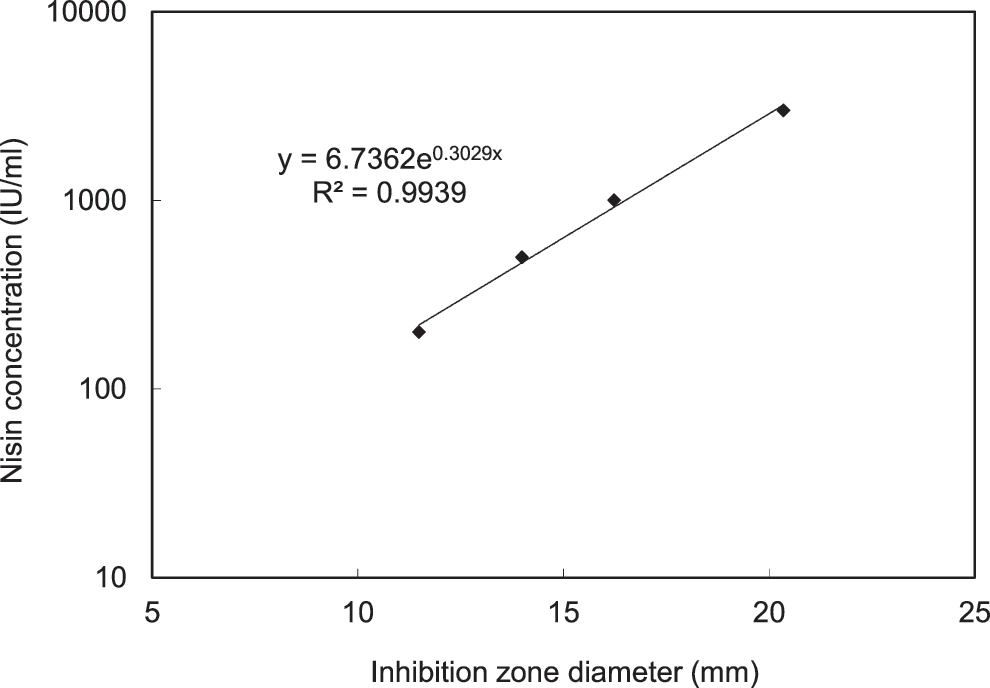
A typical nisin assay standard curve.
Ultraviolet (UV) Mutation
One loopful of the parent strain from slant was inoculated into 10 mL of liquid medium. The inoculated tube was incubated at 30°C for 15 h. Four mL of the growth broth was then transferred into a sterile centrifuge tube, and it was centrifuged at 4,000 rpm for 10 min. After discarding the supernatant, the precipitated cells were triple washed with a saline solution (0.85% NaCl), and were resuspended in the saline solution. The cell suspension was diluted with the saline solution to cell concentration of 108 cfu/mL. The cell suspension was treated under UV light (260 nm) at a dose level of about 10 mJ/cm2.
Nisin Adaptive Mutation
After UV mutation, 100 μL of the cell suspension was plated on agar plates in which nisin was added prior to medium solidification at 4,000 IU/g, 6,000 IU/g, 8,000 IU/g, and 10,000 IU/g concentrations. About 100 individual colonies grown on the nisin-containing plates were inoculated into individual tubes containing 5 mL of liquid medium each. The tubes were incubated at 30°C for 12 h. The cultured broth was then used as the seed inoculum for fermentation test. It was used to inoculate shake flasks at 1% of inoculation rate. The 500-mL shake flask contained 100 mL of fermentation medium. After fermentation at 30°C for 17 h, the pH of the fermented broth was adjusted to 2.0 with HCl, and the broth was heated and maintained at 85°C for 15 min, and then cooled down to room temperature. The parent strain was used as the control for the fermentation test under the same conditions. The fermented broths were analyzed for nisin activity using a bioassay method described above. The shake flask fermentation tests were conducted in triplicates. The mutants with the highest nisin yield were chosen for further mutations.
UV–LICL Mutation
The mutant strain #59 obtained from UV mutation and nisin adaptation was used as the parent strain in the combinational UV-LiCl mutation experiment. Four mL of the suspension of log phase growing cells were added to a 5-mL sterile centrifuge tube and was centrifuged at 4,000 rpm for 10 min. The precipitate was washed with the saline solution for 3 times, and finally a cell suspension of 108 cfu/mL was prepared for UV treatment. After UV irradiation at 10mJ/cm2, about 100 μL of the cell suspension was plated on each agar plate containing 0.1% and 0.2% LiCl for additional chemical mutation. The plates were incubated at 30°C for 48 h.
About 100 single colonies grown on the LiCl-containing plates were picked up and inoculated into tubes containing 5 mL of liquid medium. The tubes were incubated at 30°C for 12 h. The culture broth was then used as the inoculum for fermentation test. The fermentation test was conducted in a 500-mL shake flask containing 100 mL of fermentation medium inoculated with the culture broth at an inoculation rate of 1%. After anaerobic fermentation at 30°C for 17 h, the pH of the fermented broth was adjusted to 2.0 with HCl. The broth was then heated at 85°C for 15 min and cooled down to room temperature for further analysis. The parent strain was used as the control for the fermentation test under the same conditions. The shake flask fermentation test was done in triplicates. The mutant with the highest nisin yield was chosen for further mutations.
Diethylsulfate (DES)–UV Mutation and Nisin Adaptation
The higher-nisin-producing mutant strain #571 obtained from the UV-LiCl combinational mutations was used as the parent strain for a combination of DES and UV mutations followed by a nisin adaptation. The cell suspension was prepared in a similar way as described in previous mutations, except that the phosphate buffer (pH 6.8) was used in washing cell pallets and making cell suspension. DES was mixed into the cell suspension (108 cfu/mL) at 1% of concentration. The mixture was incubated on a shaker at 30°C for 30 min, and then the reaction was terminated by adding 25% sodium thiosulfate into the mixture. The reaction mixture was then transferred to a 60-mm Petri dish, and the dish was irradiated with UV for 10 mJ/cm2. The treated cell suspension was then plated on agar plates containing 8,000 IU/mL of nisin. The plates were incubated at 30°C for 48 days. About 100 individual colonies were picked up for fermentation test, as conducted in the previous mutation experiments. The shake flask fermentation test was conducted in triplicates. The mutant isolate with the highest nisin yield was selected for further mutation.
High Dose UV-Nisin Mutation
The mutant strain #1520 obtained from the DES-UV mutation was used as the parent strain in this round of mutation. In this experiment, the mutation protocol was similar to the first round of UV mutation and nisin adaptation experiment, but higher doses of UV and nisin were used. Cell suspension in the saline solution was irradiated with UV at a dose of 20 mJ/cm2, and was then plated on agar plates containing 10,000 IU/ml of nisin for a combinational UV and nisin adaptive mutation. After incubation at 30°C for 3 days, about 50 surviving colonies were picked up for fermentation test. The test was conducted in triplicates. The resulting highest nisin-producing mutant was selected for strain stability test and further scaling-up fermentation experiments.
Stability Test of Mutant Strains
The high nisin-producing mutants obtained from a series of mutations were grown and maintained on agar slants as the first generation/passage of the strains (F1). They were sub-cultured on the agar slants and the slants were incubated at 30°C for 24 hours to obtain the second passage of the mutants (F2). F2 were then sub-cultured subsequently for several times to obtain F3, F4, and F5. Each passage of the mutants was tested for fermentation, and their nisin yields were analyzed. The experiments were repeated for 3 times, and the results were reported as averages with standard deviations.
10-L Fermentation
The finally obtained high-nisin-producing mutant was activated on an agar slant. A loopful of the inoculant from the slant was transferred into a 500-mL flask containing 100 mL of liquid medium. The medium contained 1% sucrose, 1% yeast extract, 1% peptone, 2% KH2PO4, 0.2% NaCl, and 0.02% MgSO4•7H2O. It was then incubated on a shaker at 30°C and 250 rpm to a log phase of growth when cell density of OD600 reached 2.0–3.0. The seed culture was inoculated into a 10-L fermentor at an inoculation rate of 1%. The fermentation medium contained 0.5% yeast extract, 2% corn steep liquor, 1.5% sucrose, 0.4% Na2HPO4•12H2O, and 0.15% MgSO4.7H2O and was autoclave at 121°C for 20 min before inoculation. After autoclave the pH was naturally at 6.8.
The fermentation was conducted at 30°C, and the pH was maintained at 6.7–6.8 with NaOH and H2SO4, and agitation was set at 250 rpm. After 18–20 h of fermentation, the remnant sugar was below 0.2%, and the nisin content towards the end of the fermentation was analyzed by agar well diffusion assay as described above.
700-L Fermentation
The finally obtained high-nisin-producing mutant was used to prepare the seed inoculum. A loopful of the activated culture slant was transferred into a 500-mL flask containing 100 mL of liquid medium. The medium contained 1% sucrose, 1% yeast extract, 1% peptone, 2% KH2PO4, 0.2% NaCl, and 0.02% MgSO4•7H2O. It was then incubated on a shaker at 30°C and 250 rpm to a log phase of growth when cell density of OD600 reached 2.0–3.0. The seed culture was inoculated at 1% rate in a 10-L fermentor to prepare the second seed, and then the second seed was inoculated into a 700-L fermentor at an inoculation rate of 1%. In the 700-L fermentation experiment, the sucrose concentration in the fermentation medium was increased to 3%, and the inoculation rate was increased to 1.5%. The rest of the conditions remained unchanged. The fermentation temperature was at 30 ± 1°C, and the pH of the medium was maintained at 6.7-6.8 with NaOH, and the agitation was set at 30HZ. The fermentation was ended when the remnant sugar was below 0.2%. The fermentation was conducted 6 times.
50-TON Fermentation
Finally, the fermentation was scaled up to production scale. In this 50-ton fermentation experiment, the finally obtained high-nisin-producing mutant was used to prepare the seed inoculum in the same way as in the 700-L fermentation, with an additional third seed preparation step using the 700-L fermentor. The fermentation medium and the inoculation rate were also the same as in the 700-L fermentation. The fermentation temperature was at 30 ± 1°C, and the pH was maintained at 6.6–6.8 with NaOH. The agitation was set at 30 Hz. The fermentation was terminated when the remnant sugar was below 0.2%. The 50-ton fermentation experiment was conducted 15 times.
Results and Discussion
Classical and Adaptive Mutations for Strain Improvement
In the nisin-adaptation experiments, the number of colonies on each nisin-containing plate decreased as the nisin concentration in the medium increased. It was found that the plates containing 8,000 IU/g of nisin had 30–150 colonies per plate. Therefore, the nisin concentration in the agar medium was chosen at 8,000 IU/g for mutant adaptation screening following a physical or chemical mutation.
The parent strain L. lactis ATCC11454 had a nisin activity of 640 IU/mL in the shake flaks fermentation test, and after the first round of UV mutation flowed by nisin adaptation, the two best mutants—#59 and #114—were obtained. Their nisin activity in the same shake flask test was 1,210 IU/mL and 1,190 IU/mL, respectively.
In the second round of mutation, strain #59 was further mutated by UV-LiCl combinational treatments, and the two best mutants, #571 and #573 with nisin yields of 1,570 IU/mL and 1,470 IU/mL, respectively, were obtained.
In the third round of mutation, as a result of a combinational treatment of strain #571 with DES, UV, and nisin adaptation, the mutant strain #1520 with an improved nisin yield of 1,870 IU/mL in the shake flask fermentation was obtained.
Finally, upon the high dose UV mutation and nisin adaptation in the fourth round of mutation, two high-nisin-producing mutants, #3807 and #3823, were obtained. They produced 2,160 IU/mL and 2,140 IU/mL of nisin, respectively, in the shake flask fermentation test.
The four rounds of mutation processes and genealogy of the mutants with their nisin yields are summarized in Fig. 2.
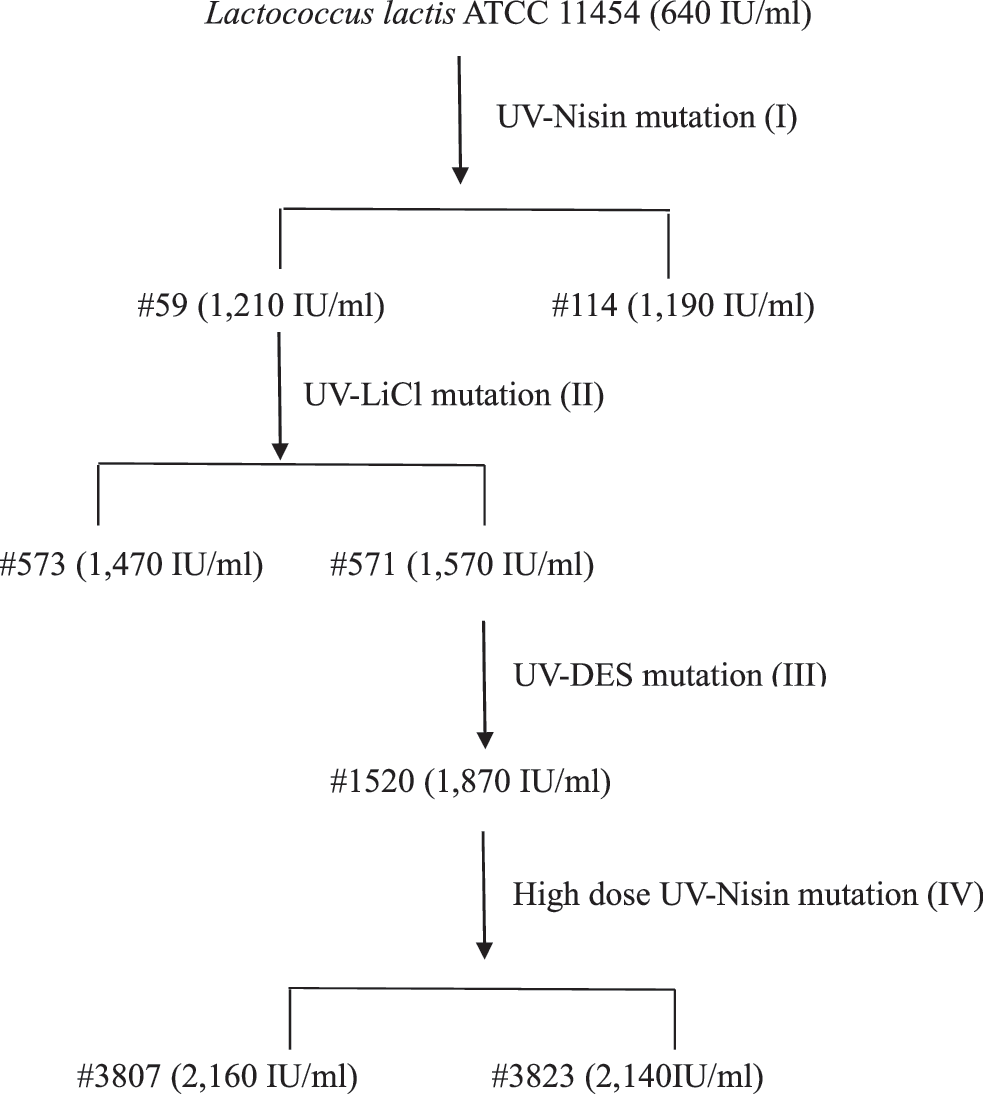
Genealogy of the mutants with their nisin yields
The traditional mutation when combined with adaptative mutation or evolution appears to be an effective way to improve nisin-producing strains. Through a series of physical and chemical mutations by UV, DES, LiCl and nisin treatments, nisin production yield of was improved by more than 3-fold compared with the original strain under the shake flask fermentation conditions.
The Stability of Mutants
The stability test results suggest that the high-nisin-producing isolates obtained from a series of mutations and adaptations were fairly stable in the first 3 passages of sub-culturing and started to lose nisin yield very slightly afterwards (Table 1).
Nisin Yield of Mutants #3807 and #3823 as Affected by Sub-Culturing
In order to maintain the mutant's capability to produce a maximum level of nisin in commercial fermentation production, the working vials should be made directly from the master stocks which should be prepared in large quantity from the first cell line of the mutant. If more than 3 passages of sub-culturing are required, a culture purification process may be conducted and the fermentation of the isolates should be tested to ensure the best performance of the purified isolates. To maintain the stability of the mutants, it is recommended that nisin be added to the maintenance medium to keep the selective pressure for the mutants to maintain their nisin resistance attribute which is important for high yield nisin production.
10-L Fermentation
The 10-L fermentation experiment with the mutant #3807 was repeated 3 times, and the nisin yield results are summarized in Table 2. The nisin yield was significantly increased compared with the shake flask results. This was probably due to pH control in the 10-L fermentor. On average, it yielded 11,870 IU/mL of nisin in the fermented broth after 20 h of fermentation.
The Nisin Yield in 10-L Fermentation with Mutant #3807
The repeatability of the fermentation results was also good. This is a very positive indication of scalability of the process with the new strain.
700L Fermentation
To test its scalability, the nisin fermentation with the mutant #3807 was scaled up to 700 L, and the fermentation was run independently 6 times. Nisin yields in 700-L fermentors are shown in Table 3. The fermentation time in 700-L fermenters was shorter than in 10-L fermentors, and the nisin yield was slightly lower. The average nisin yield after 14-h fermentation in the 6 batches of the 700-L fermentors was 10,350 IU/mL.
The Nisin Yield in 700-L Fermentation with Mutant #3807
The larger fermentor seemed to have faster fermentation, and nisin was produced more rapidly in the fermentor. It only took 14 h of fermentation, and the average nisin yield was close to that of 10-L fermenters at 20 h of fermentation. The repeatability of the fermentation was good too, as the nisin yield in the 6 batches was within a reasonable range. The 700-L fermentation results confirmed the scalability of the new strain and fermentation.
50-TON Fermentation
To further validate the scalability of the new strain, the fermentation was scaled up to 50 ton as the real production scale and repeated 15 times. The nisin yields are shown in Table 4. The average yield of the 15 batches of fermentation was 9,960 IU/mL at the end of fermentation, just slightly lower than the 10-L and 700-L fermentation yield. The repeatability at this full production scale was quite reasonable. The standard deviation of these 15 batch yield results was 950 IU/mL.
The Nisin Yield in 50-Ton Fermentation with Mutant #3807
As a comparison, the parent strain ATCC 11454 was tested in 50-ton fermenters under the same fermentation conditions for 14 h in 3 replicates. The nisin yield results are summarized in Table 5. Although the nisin yield of the parent strain was significantly improved in the 50-ton fermentor compared to the shake flask results, it is still way lower than that of mutant #3807.
The Nisin Yield in 50-Ton Fermentation with the Parent Strain ATCC11454
At this point, the commercial scalability of the new strain has been completely verified, and the nisin yield was acceptable from an industrial production standpoint. As long as the strain is maintained properly according to normal production protocols, the strain is suitable for commercial production with a high yield of nisin A and a relatively short fermentation time.
Conclusions
When countries reject or ban GMO crops over safety concerns, many companies often turn to developing new strains using classical mutagenesis, wherein plants are subjected to radiation treatments or dosed in toxic chemicals that randomly scramble genes to produce new traits. In the fermentation industry, classical mutagenesis or mutation is also still one of the most widely used strain-improvement technologies today. The resulting mutants are generally considered non-GMOs. Despite the fact that the mutagenesis process is much less precise than genetic modifications—in which scientists take a gene that gives rise to a desired trait—mutagenesis is unregulated and widely accepted by consumers. Some products made from mutated strains can even be sold as organic. Many major nisin manufacturers have conducted their own mutations on the natural isolates to improve their nisin yield, but such efforts and information have largely been kept as trade secrets by nisin manufacturing companies. Very limited information on the mutation of nisin-producing strains is available in the public domain.
In nature, nearly all microorganisms are capable of adaptation. Adaptation is a dynamic, ongoing, life-sustaining process by which living organisms adjust to environmental factors or changes for the ultimate purpose of survival, reproduction, and an optimal level of functioning. Adaptation is the evolutionary process whereby an organism becomes better able to live in its habitat or habitats. 30 Adaptive evolution results from the propagation of advantageous mutations through positive selection, and it is a conventional approach to obtaining natural mutants with enhanced phenotypic properties as desirable traits, such as higher tolerance to acid, heat, salt or other stress factors. 31 Due to its non-GMO nature, adaptive evolution has gained renewed interest in recent years for improving industrial production strains to increase yield or enhance certain targeted traits or properties. 32,33
As a traditional breeding method, mutagenesis remains an effective way to improve certain desirable traits or properties of microorganisms. As it is not regulated in the same way as GMOs, mutation is a very practical and widely acceptable approach for industrial strain improvement. In this study, a combinational concept of traditional mutation and adaptive mutation was utilized, and the nisin yield was significantly improved. The adaptive mutation was used as an additional pressure following traditional mutations, and, at the same time as a selective medium to screen for nisin-resistant mutants. Nisin resistance seems to be a necessary trait of high nisin-producing mutants. By selecting an appropriate mutation dosage and nisin-resistance level, through a series of traditional and adaptive mutations two high nisin-producing mutant isolates were obtained. The mutants were relatively stable over sub-culturing. They were also proved to be readily scalable and were able to produce a high level of nisin in large-scale fermentations. These mutants have great potential in the commercial nisin-A production. Since both mutation and adaptation processes are considered natural and non-GMO process, the combinational treatment is a very practical, simple, and low-cost technology for industrial strain improvement with minimum hurdles to both consumer and regulatory acceptance in countries where GMOs are a concern.
Footnotes
Author Disclosure Statement
No competing financial interests exist.