
Abstract
Biopolymers like polyhydroxyalkanoates (PHA) are a class of metabolites with promising applications in the fields of environmental, agricultural, and biomedical sciences. Commercial production of polyhydroxyalkanoates is a high-cost process generally carried out using heterotrophic species of bacteria. Using algae for intracellular PHA accumulation is an alternative method for bioplastic production. In the present study, cyanobacterium Synechococcus elongates was identified as a potential PHA-producing organism. A range of experimental conditions that include different growth parameters of nitrogen and phosphate starvation with the addition of external carbon sources under photoautotrophic and chemoheterotrophic growth conditions were examined. Our observations revealed that the algal cells cultured in nitrogen-starved medium under phototrophic conditions yield the highest quantity of PHA (17.15%) when 1% sucrose is used as a carbon source. Similarly, under chemotrophic conditions, the highest PHA synthesis (12.98%) was observed in medium containing 1% sucrose. In case of phosphate-starved medium, the highest PHA production was recorded as 7.02% under phototrophic conditions when 1% fructose was used as a carbon source. Similarly, 5.40% PHA synthesis occurred when media was supplemented with 1% glucose under chemotrophic conditions. The present study shows that nitrogen-deficiency stress induced higher PHA yield compared to phosphate-deficiency stress in S. elongates under photoautotrophic conditions when sucrose was present as an external carbon source. Our findings indicated that 17.15% cell dry-weight of PHA was the highest yield obtained from algal forms under nutrient-deficient stress conditions. This yield is lower than the bacterial PHA-producing system. Therefore, more studies are needed to enhance the yield of PHA in algae.
Introduction
Polyhydroxyalkanoates (PHA) are polyesters synthesized and accumulated by different microorganisms, usually as they enter the stationary growth phase. PHA forms intracellular inclusions and can be catabolized to produce carbon and energy. Under conditions of limited nitrogen availability and excess carbon, some bacteria can accumulate 60–80% of cell weight as PHA. PHA is a high molecular weight reserve material stored in the form of highly reduced carbon, which does not significantly affect the osmotic potential of the cell. 1 –3
PHA is similar to polypropylene in material properties, but is biodegradable in nature. 4,5 PHA can be biodegraded completely into water and carbon dioxide (CO2) in aerobic conditions (and methane under anaerobic conditions) by bacteria in situ. PHA properties, which include natural origin, biodegradability, biocompatibility, piezoelectricity, optical purity and thermo-plasticity, make it a suitable alternative to synthetic plastics. 6 In addition to routine uses like manufacturing of bags, cloth etc., it is also known to have high-end applications in environmental, agricultural and biomedical sciences. PHA can be used to produce disposable items, artificial bones, blood vessel replacements and scaffolding materials used in tissue engineering of heart valves. 7 Poly-3-hydroxybutyrate (PHB) is a well-known type of PHA being exploited commercially. 8
The major hurdle in commercial production of PHA is the high cost of bacterial fermentation processes, which make bacterial PHA more expensive than petroleum-derived polymers such as polypropylene and polyethylene. The vital factors leading to cost escalation of PHA production are the expensive carbon sources and pure oxygen supply. For the economical production of PHA, scientists are experimenting with novel substrates and recombinant bacterial strains developed by metabolic engineering to achieve optimum yield. 9
Some wild-type cyanobacteria like Nostoc muscorum, Spirulina platensis, Synechococcus and Spirulina subsalsa, are reported to accumulate PHA from CO2. In this context, cyanobacteria are promising host systems due to their minimal nutrient requirements and photoautotrophic nature. These organisms have a short generation time, and growth requires simple inorganic nutrients such as phosphate, nitrate, magnesium, sodium, potassium and calcium as macronutrients and minor elements like Mn, Zn, Mo, Co, Fe and Cu as micronutrients. 10 Though studies indicate that bacteria are better host systems for PHA production, algae are easy to cultivate and maintain, and therefore can be used as alternative production systems.
A major breakthrough in cyanobacterial PHA research was achieved by Nishioka et al. in 2001 with a thermophilic cyanobacterium, Synechococcus MA19 strain, which was reported to accumulate PHA up to 55% dry cell weight (dcw). 11 Subsequently, Panda et al. and Panda and Mallick demonstrated PHA accumulation up to 38% (dcw) in Synechocystis sp. PCC 6803 under various specific conditions. 12,13
In the present study, an unexplored unicellular cyanobacterium Synechococcus elongates was examined as a possible potent producer of PHA. S. elongates is a rod-shaped freshwater photoautotrophic cyanobacterium that habituates freshwater springs, particularly those with a mesophilic or moderate temperature range. S. elongates belongs to the family Synechococcaceae, the genera of which are reported to synthesize PHA. For high PHA accumulation, phosphate- and nitrate-starved media supplemented with carbon sources like glucose, fructose, sucrose, glycerol and citrate were tested under mixotrophic conditions. 14 This resulted in minor growth inhibition, but generated enough stress to induce PHA synthesis.
The objective of the present study was to evaluate the effect of different carbon sources and nutrient starvation on PHA production in S. elongates under mixotrophic conditions.
Materials and Methods
Test Organisms and Experimental Conditions
Cyanobacterium S. elongates was obtained from the culture collection of National Facility for Marine Cyanobacteria (NFMC), Bharathidasan University (Trichi, India). It was cultured in ASN-III medium (Artificial Sea Nutrient Media) at 24°C under illumination with cool white fluorescent light and light/dark cycle of 14/10 h (Fig. 1). After incubation for 15 days, the culture was transferred to two sets of ASN-III media; the first set consisted of nitrogen-deficient ASN-III media (media without NaNO3) while the second consisted of phosphate-deficient ASN-III media (media without K2HPO4). Variations of growth media for carbon sources were as follows: • N2-deficient medium with 1% glucose • N2-deficient medium with 1% fructose • N2-deficient medium with 1% sucrose • N2-deficient medium with 1% glycerol • N2-deficient medium with 1% citrate • PO4-deficient medium with 1% glucose • PO4-deficient medium with 1% fructose • PO4-deficient medium with 1% sucrose • PO4-deficient medium with 1% glycerol • PO4-deficient medium with 1% citrate

Growth of S. elongates in ASN-III media. Color images available online at
Two experimental sets of each of the above media were prepared and incubated for 15 days. One set was incubated under illumination with cool white fluorescent light and a light/dark cycle of 14/10 h, while the other set was incubated in the dark, thereby achieving mixotrophic conditions.
Microscopy
The morphological analysis of the PHA granules was carried out using Sudan-black staining and microscopy. A smear of the algal suspension was prepared on a slide and flooded with 0.3% Sudan-black stain for 15–20 minutes. The slide was destained under running water followed by addition of 1% aqueous solution of safranin. After incubation for 15 min, the slide was again washed with water and dried. The stained cells were observed under 100X magnification. 15
Estimation of Biomass
The cell growth was analyzed gravimetrically. Cell biomass was obtained by centrifugation (8,000 rpm, 10 min) of the culture media containing cells followed by drying the cell pellet in an oven until constant weight was reported. The biomass was expressed as cell dry weight.
Extraction of Polyhydroxyalkanoate (PHA)
The harvested cells were washed with distilled water to eliminate impurities. The cells were suspended in methanol and incubated overnight at 4°C to remove pigments. The cells were then centrifuged. Methanol was decanted and the cell pellet was dried at 60°C. These cells were then subjected to repeated extraction steps with hot chloroform. PHA was then precipitated from the chloroform solution into chilled methanol. Methanol was separated from the methanol-chloroform mixture using a separating funnel, followed by evaporation of methanol. 7 After evaporation of the solvent, PHA was obtained as a tough, translucent film (Fig 2).

PHA film. Color images available online at
Quantitative Analysis of PHA
The quantitative estimation of PHA was done spectrophotometrically, per Law and Slepecky's method. 16 PHA sample was dissolved in chloroform and transferred to a capped test tube. The chloroform was evaporated and 10 mL of concentrated H2SO4 was added. The tube was capped and heated for 10 min at 100°C in a water bath. The solution was cooled and thoroughly mixed. A sample was transferred to a silica cuvette, and absorbance was recorded at 235 nm against a sulphuric acid blank. The concentration of PHA in the sample was estimated from a standard curve of crotonic acid (Figs. 3–4). 16
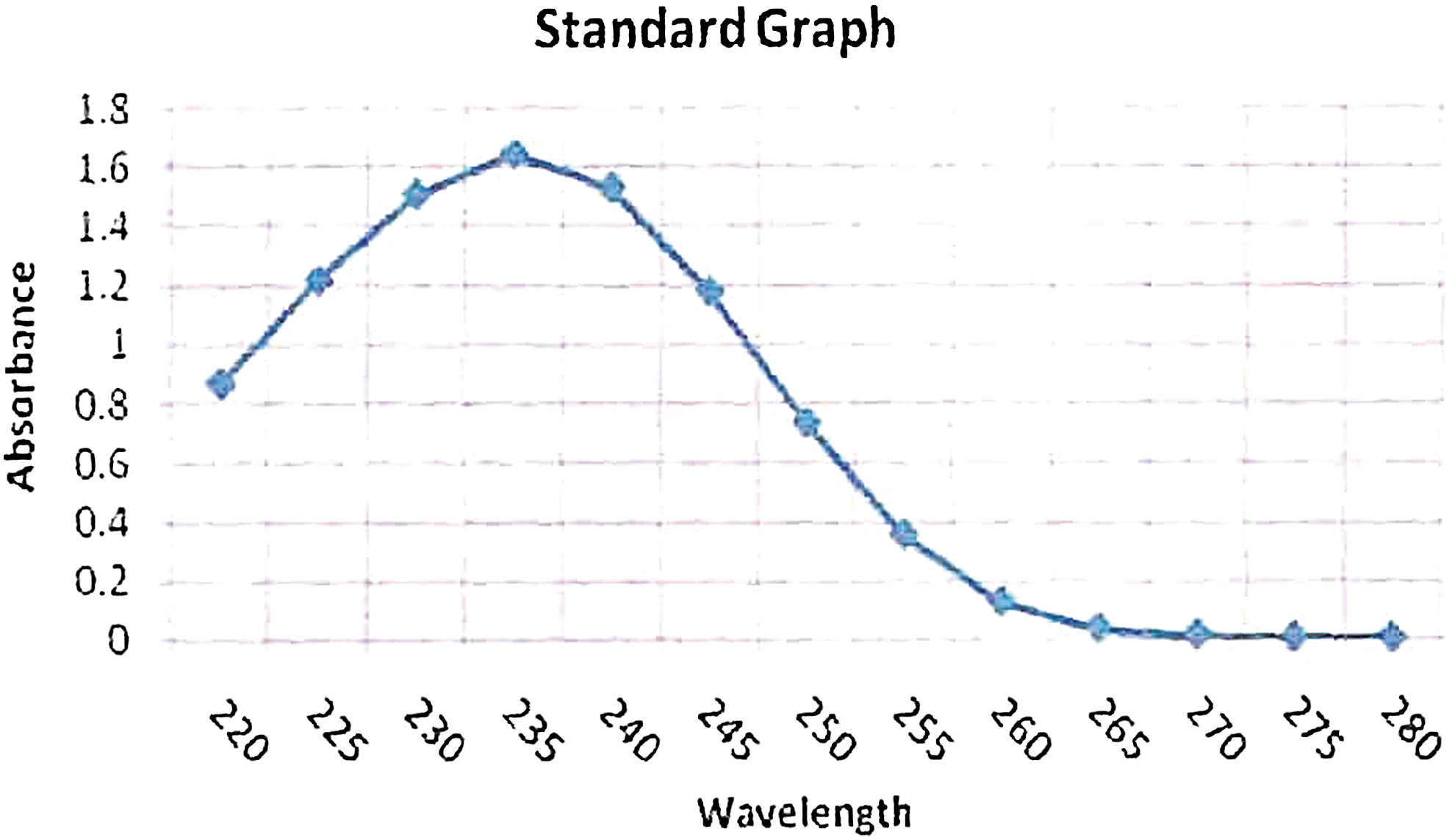
Standard graph for crotonic acid with maximum absorbance at 235 nm. Color images available online at
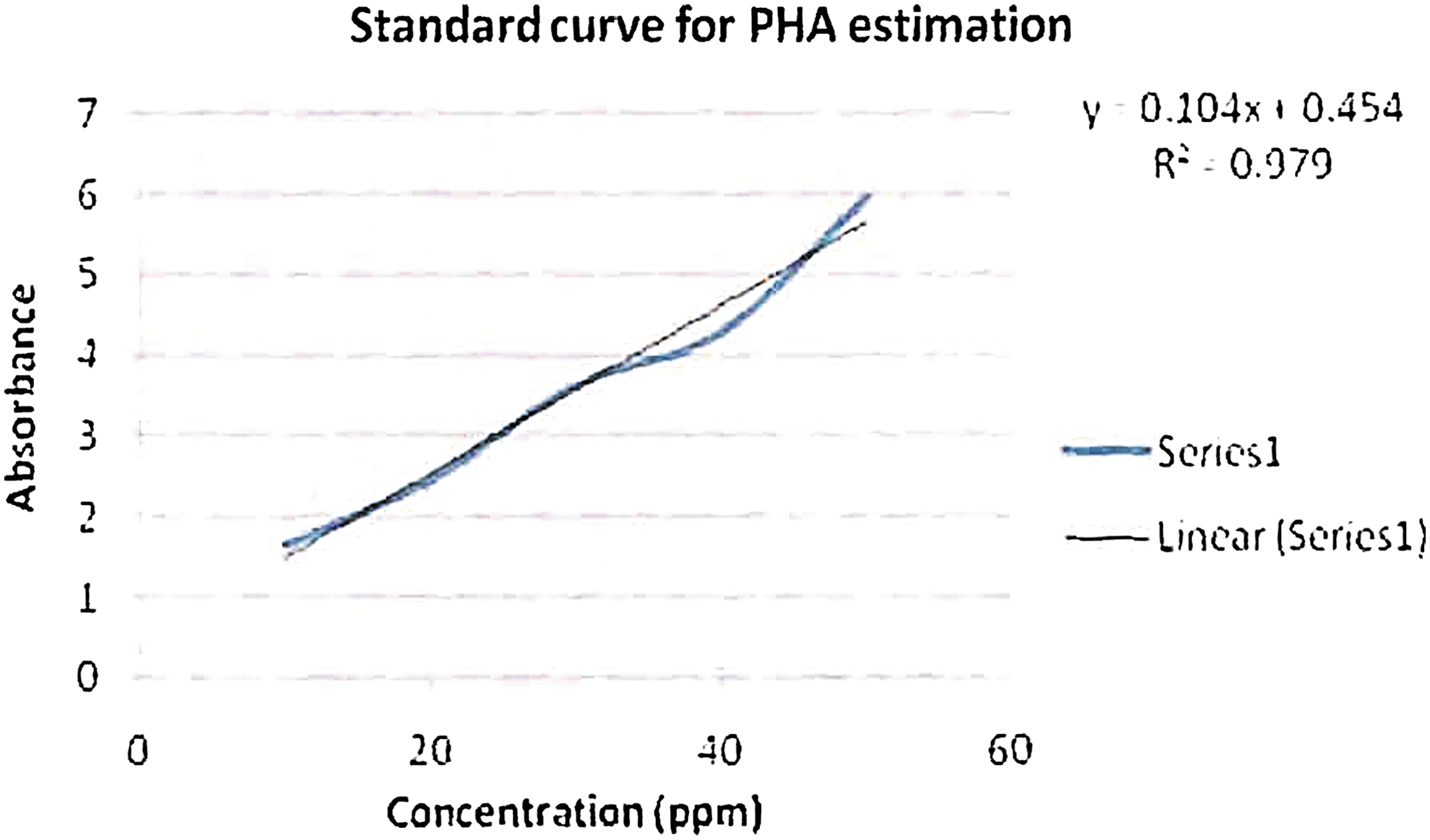
Standard curve for quantitative estimation of PHA. Color images available online at
Results and Discussion
The growth pattern of Synechococcus elongates was studied, and it was found that the culture reached stationery phase after 12 days of incubation (Fig. 5).
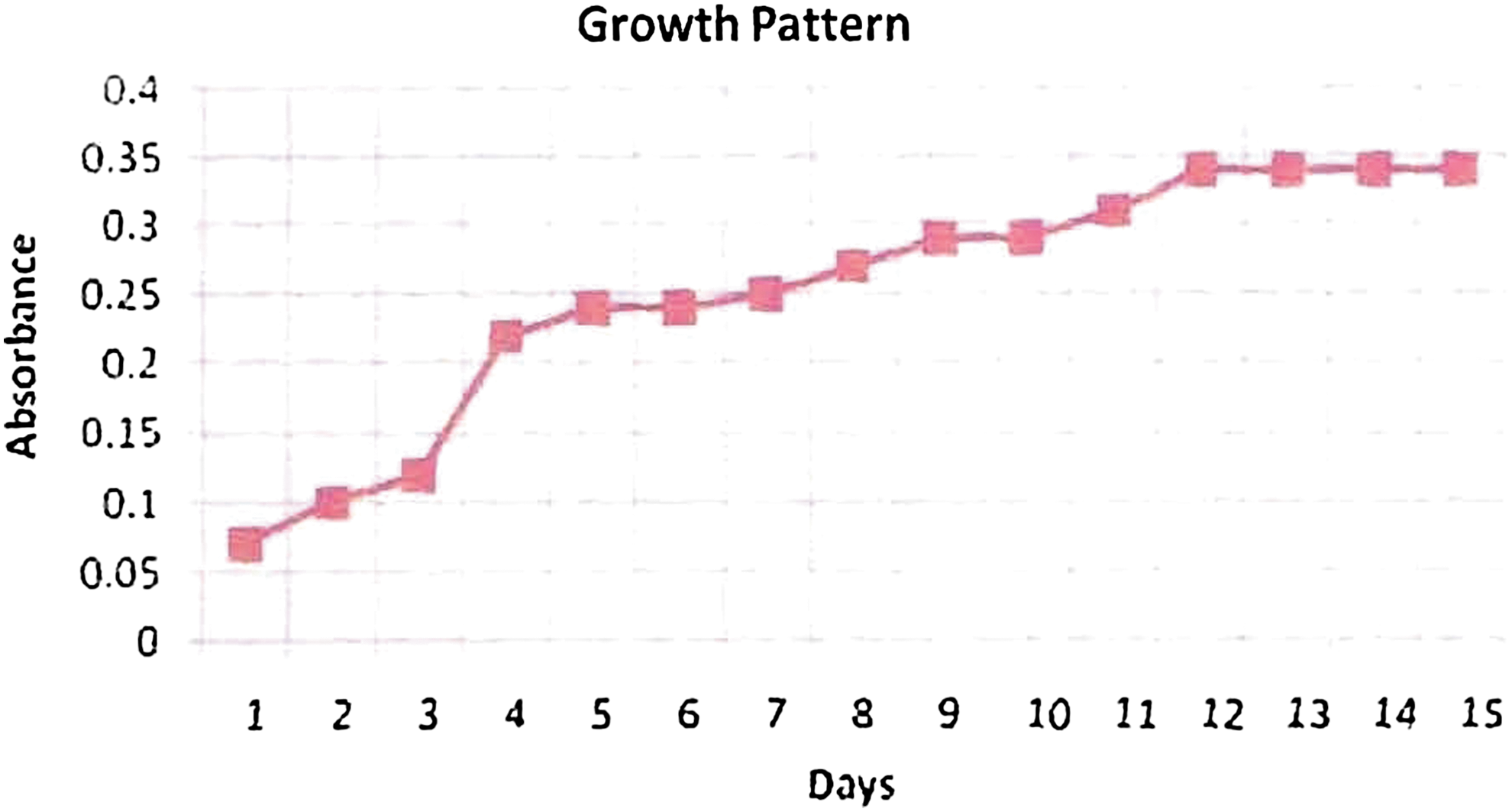
Growth pattern of S.elongates. Color images available online at
Microscopic Analysis
The morphology of Synechococcus elongates culture was observed under a light microscope (Fig. 6). On staining with sudan-black, the cells of Synechococcus elongates containing PHA granules were observed as dark-blue or black spots against red cytoplasm (Fig. 7a–b).
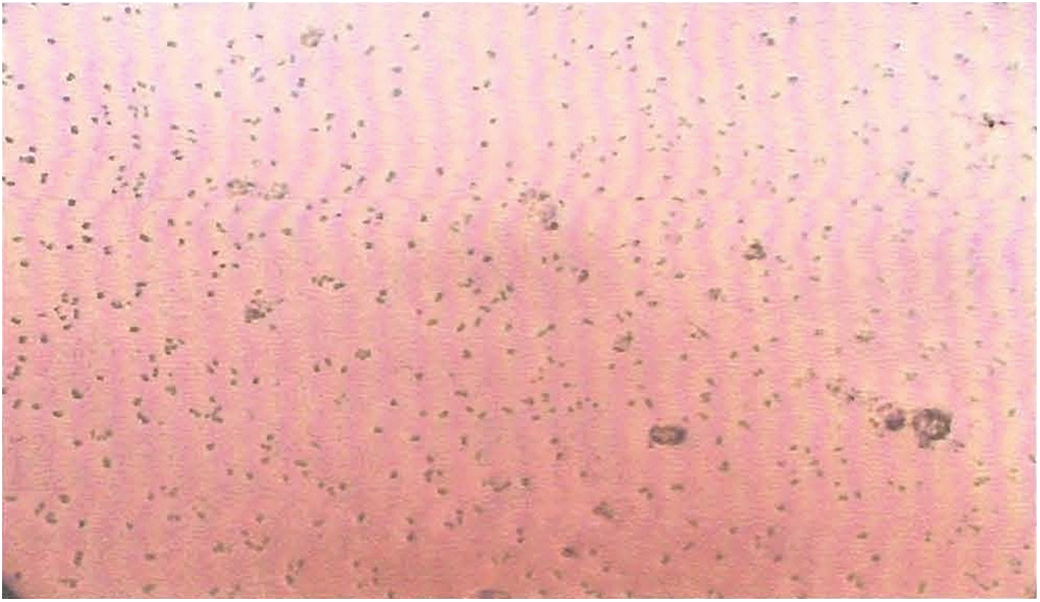
S. elongates cells observed under Light microscope (100 X). Color images available online at
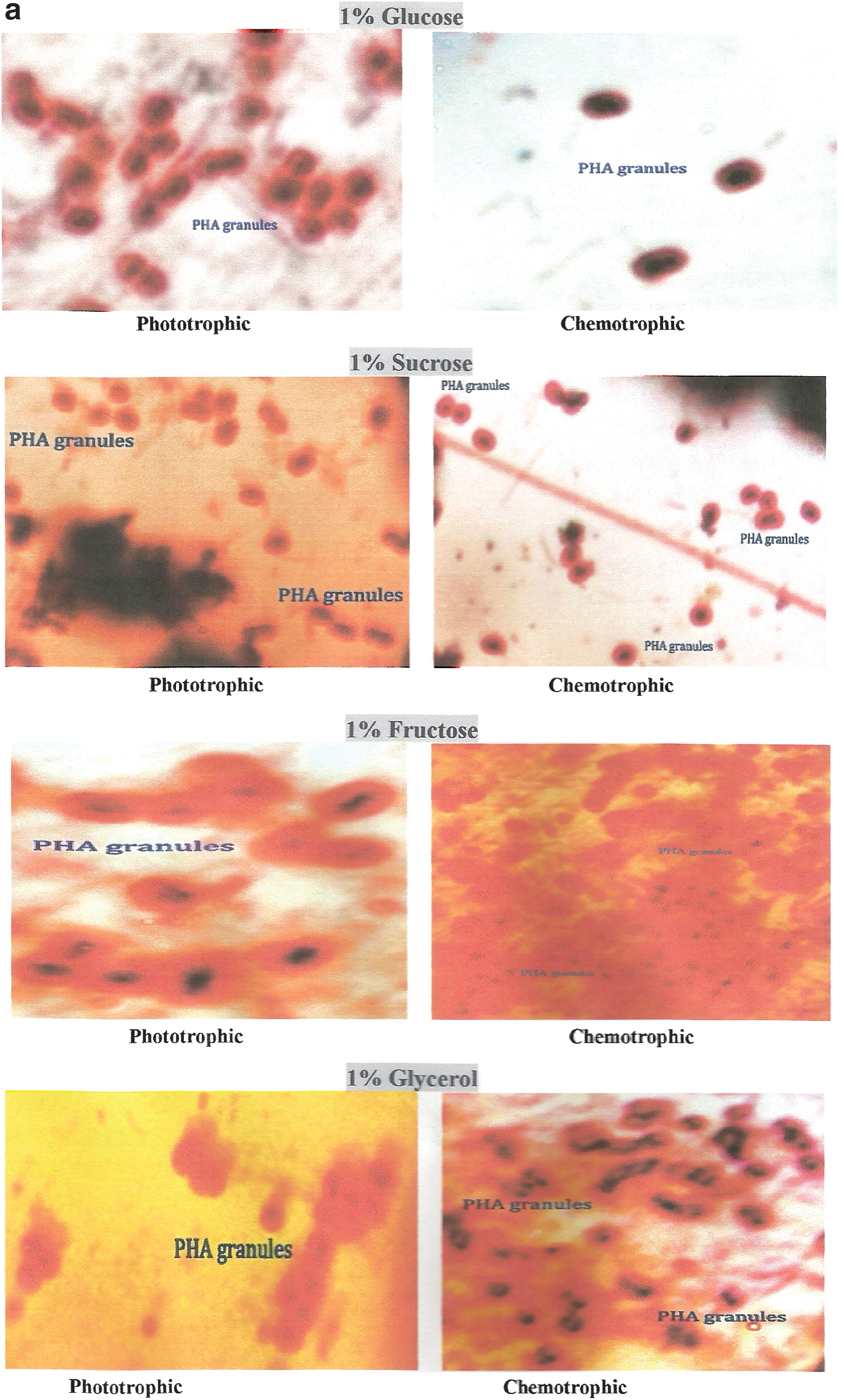
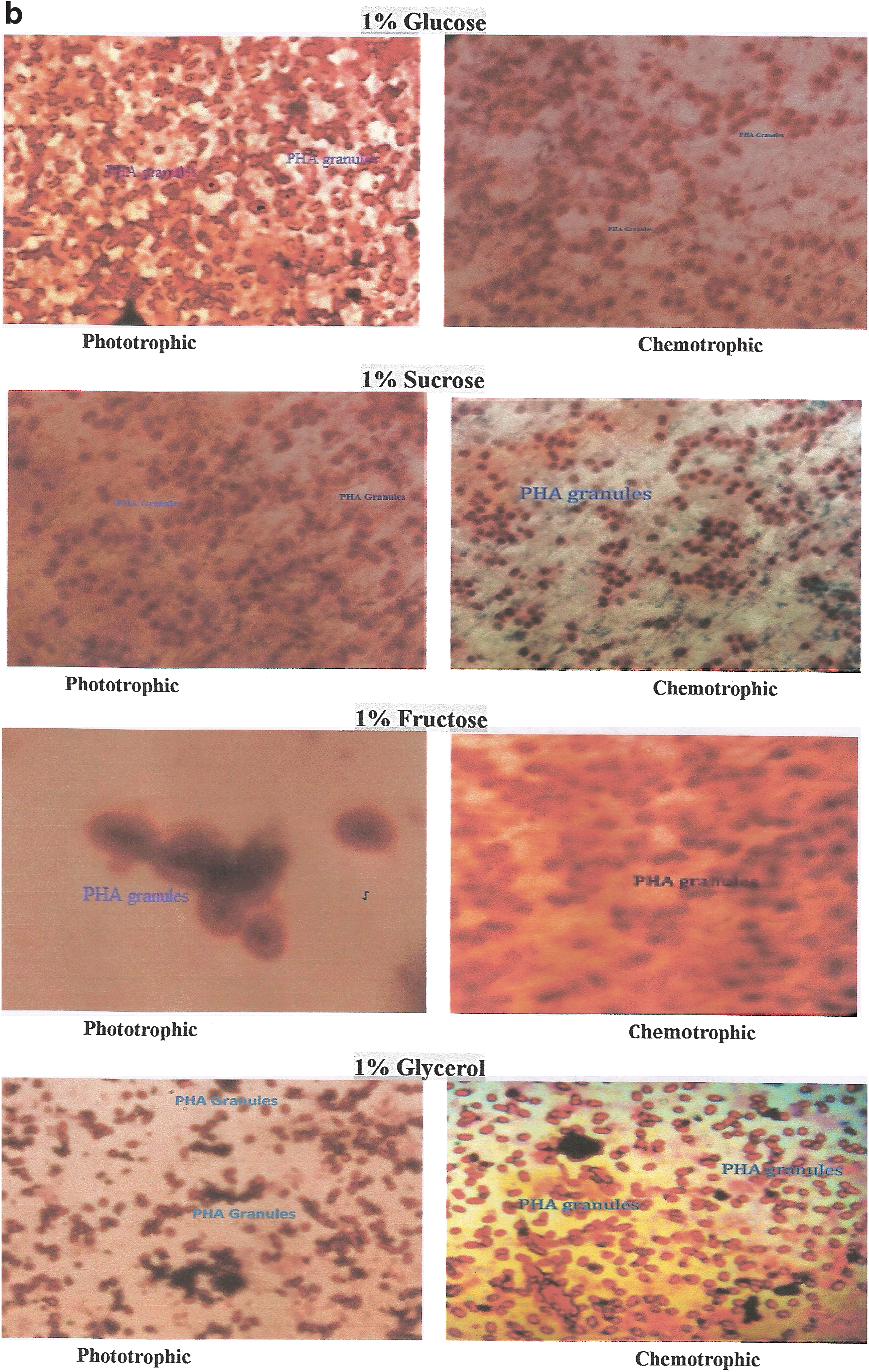
Qualitative and Quantitative Estimation of PHA Under Different Conditions
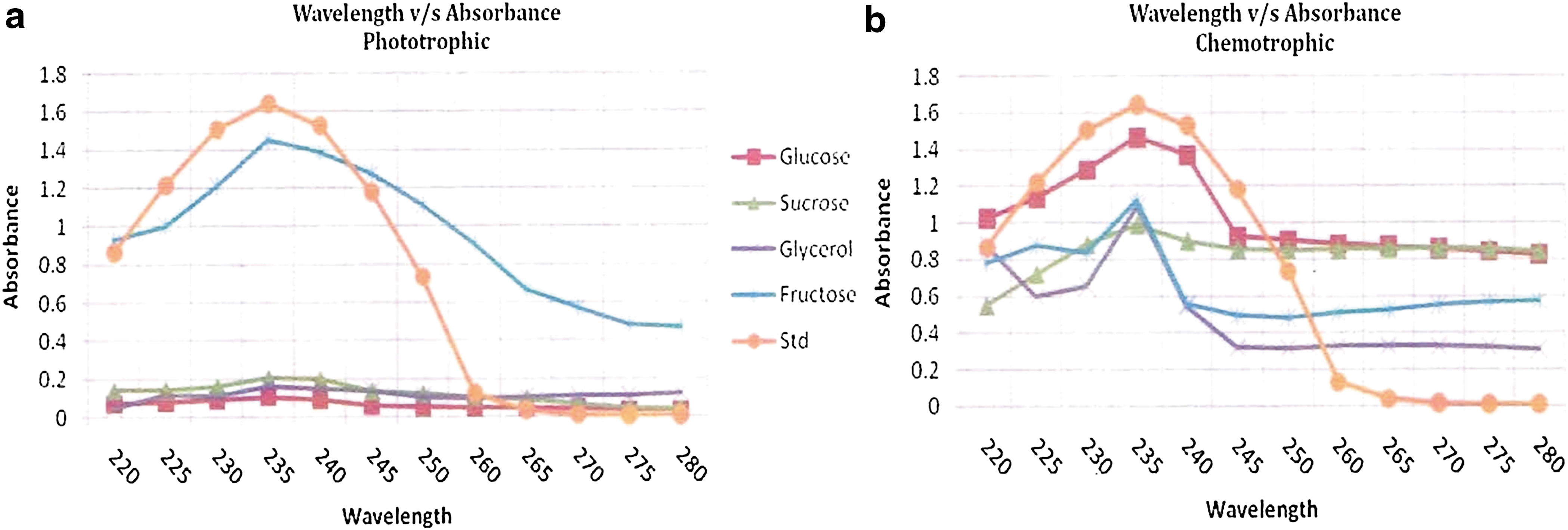
Findings in the present investigation indicated that the highest yield of PHA was obtained from the cells cultured in nitrogen-deficient ASN-III media conditioned with sucrose as a carbon source under photoautotrophic conditions as well as chemoheterotrophic conditions (Table 1, Fig. 8a–b). However, in phosphate-deficient media, the results were variable. Under photoautotrophic conditions, the highest yield of PHA was obtained from the cells cultured in ASN-III media supplemented with fructose as a carbon source, while under chemoheterotrophic conditions the optimum PHA production was observed in phosphate-deficient ASN-III media supplemented with glucose as a carbon source. (Table 2, Fig. 9).
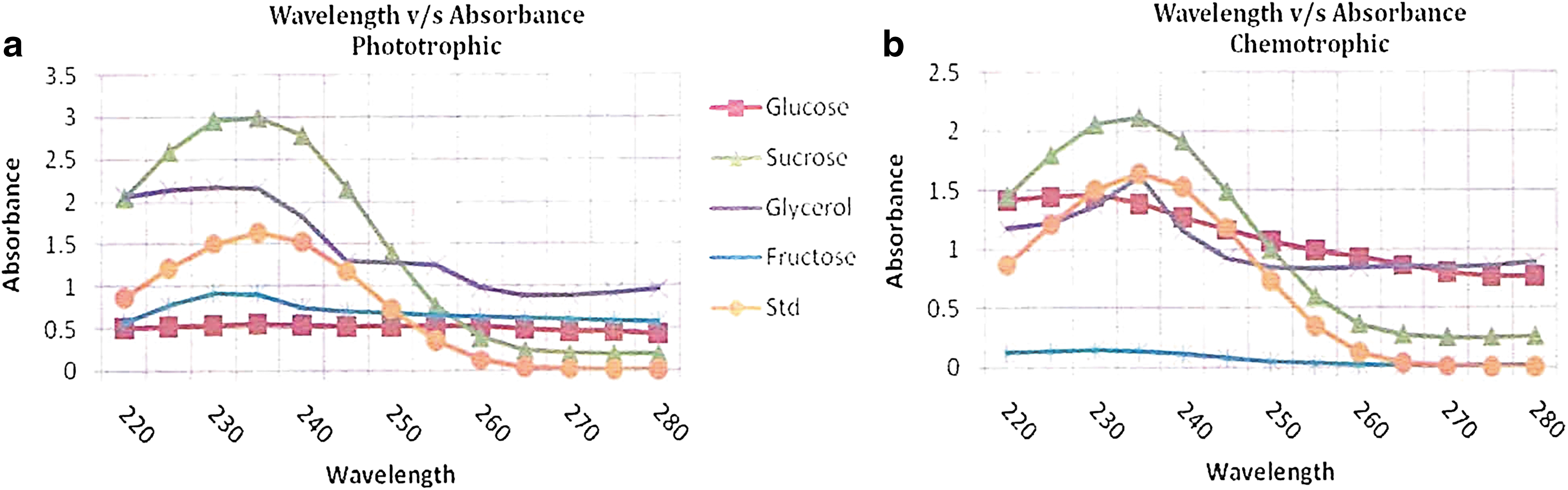
PHA Production in S. elongates Using Nitrogen Deficient ASN-III Media
Values are the mean ± SE; n = 3
PHA Production in S. elongates Using Phosphate-Deficient ASN-III Media
Values are the mean ± SE; n = 3
Discussion
The results demonstrate that growth nutrients like nitrogen and phosphorus have pronounced effects on the PHA-synthesizing ability of S. elongates. Phosphate-deficient conditions resulted in minor growth inhibition compared to nitrate-deficient conditions. 17,18 The cells incubated in nitrogen-deficient media entered the stationery phase earlier as their growth was inhibited due to unavailability of nitrogen. The excess carbon available in the media was utilized by the cells to synthesize carbon storage molecules like PHAs, which would be consumed during starvation for their survival. On the contrary, cells incubated in phosphate-deficient media remained metabolically active for a longer time and did not accumulate sufficient amount of PHA. Hence, the PHA yield was higher in nitrogen-deficient media than phosphate-deficient media. Similarly, cells incubated in photoautotrophic conditions showed higher PHA-yielding capacity than chemoheterotrophic conditions, as the cells could obtain energy from two sources: light and exogenously provided carbon. Even after exhaustion of the exogenous carbon source, the cells could derive energy from light via photosynthesis.
The current investigations also revealed that S. elongates was capable of utilizing different carbon sources. Glucose utilization in the cyanobacteria species like S. elongates occurs via the Pentose Phosphate pathway (PPP). The positive effect of glucose on PHA production could be related to the increased supply of nicotinamide adenine dinucleotide phosphate (NADPH) by elevated activity of 6-phosphoglucose dehydrogenase, the first enzyme of PPP. NADPH is a prerequisite for the activity of the enzyme acetoacetyl-CoA reductase for conversion of acetoacetyl-CoA to β-hydroxybutyryl-CoA, which is the precursor of PHA. A similar explanation is also valid for the increased PHA yield in fructose-supplemented cultures, since fructose is an isomer of glucose. 10 As sucrose is a disaccharide of glucose and fructose, a synergistic effect was observed. Citrate supplementation was found to inhibit biomass concentration significantly, which led to the negligible PHA synthesis. Similarly, addition of glycerol had no significant effect on PHA accumulation. This may be attributed to the fact that cells failed to regulate the uptake and metabolism of glycerol into the cell. 14 Thus, maximum yield was achieved in the cells supplemented with sucrose in nitrogen-deficient media under photoautotrophic conditions.
Table 3 compares S. elongates PHA yield (in % dcw) with that of other cyanobacterial strains reported in literature. PHA yield from S. elongates was comparable with the yield obtained from other cyanobacterial strains where nutrient limitations were used as a PHA-stimulating factor. Further research involving application of Research Surface Methodology tools for media optimization could give better results.
Comparative Account of PHA Yield (%dcw) from S. elongates with PHA Yields Obtained from Other Cyanobacterial Sources
The commercial methods of PHA production mostly use bacteria as the producing organisms, and industrial waste like sugarcane molasses or whey as a substrate. 19 The use of synthetic media is restrictive due to cost. Currently, there are no significant reports of PHA production using algae at commercial scale, and there is abundant scope for researchers to undertake studies related to PHA production in algal species. With the same pretext, the current research proposed a model for PHA synthesis using S. elongates as the producer organism.
Conclusion
Cyanobacteria have the potential to produce biopolymers like PHA. Due to their minimal nutrient requirements and ability to grow—even in wastewater—in the presence of CO2 and sunlight, cyanobacteria can be explored as a source for PHA production. Although the yield of PHA is lower compared to heterotrophic bacteria, it can be increased by various means, such as nutrient-limiting conditions, stress conditions, the application of different PHA-enhancing precursors, and metabolic/genetic engineering. In the current study, the highest PHA yield of was obtained in the medium supplemented with sucrose, a disaccharide. This leads to the conclusion that high molecular weight carbohydrates are potent carbon sources for PHA synthesis. The study also provided first-time evidence of the potentiality of S. elongates for PHA synthesis. The current research work thereby developed a lab-scale process for synthesis of PHA in algal cells using synthetic carbon sources.
Footnotes
Acknowledgments
We thankfully acknowledge, Director, Institute of Science, Mumbai, for his valuable support. We extend our grateful acknowledgment to Dr. L. Uma, Director, National Facility for Marine Cyanobacteria (NFMC; Tamil Nadu, India) for providing unialgal strain of S. elongates. We also like to acknowledge faculty associated with Instrumentation Facility, Institute of Science, for help in characterization studies.
Author Disclosure Statement
No competing financial interests exist.