
Abstract
This work aimed to optimize the process parameters for exo-polygalacturonase (exo-PG) recovery and production from Aspergillus niger ATCC 9642. Different types of solvents and extraction solutions (distilled water, NaCl, Tween 80, citrate buffer, and acetate buffer) have been evaluated for enzymes recovery using agitation (12 to 348 rpm), time (4.8 to 55.2 min), temperature (13.2 to 46.8°C), and extraction cycles (10 cycles). Our work's maximum exo-PG activity was 23 U/g wm (68 U/g dm ), obtained after 192 h of bioproduction with 9 g orange peel, 4 g corn steep liquor, and 7 g wheat bran concentrations, at 30°C, and 5 × 106 spores/g wm . The maximum recovery was obtained using NaCl (0.1 mol/L) as solvent at a 5:1 (v:w) ratio, 30-55 min, 180 rpm, in one extraction cycle. Optimum pH and temperature for crude enzyme extract were 4.5 and 55°C, respectively.
Introduction
Among industrial-use pectinolytic enzymes, polygalacturonases (PGs; EC 3.2.1.67), the most abundant pectinolytic enzymes are involved in polygalacturonic acid chain hydrolysis, with the addition of water. Pectinases are used for fruit or vegetable processing as well as in beverage, paper and cellulose, textile, and pharmaceutical industries. 1,2
Biotechnological processes, notably solid-state cultivation, benefit from low water activity inputs and microorganisms adaptable to the environment, such as filamentous fungi. 3 Such technology can produce extracellular enzymes, such as pectinase, performing a key role on the harnessing of solid residues.
The agro-industrial sector generates a considerable amount of byproduct, mainly carbon and mineral components that can be used as clean alternatives for producing high value-added biocompounds, thus reducing overall environmental impacts. 4 –7
For this reason, agro-industrial residues with high pectin levels have been researched to produce pectinolytic enzymes in solid-state cultivation, using feedstocks such as lemon and orange peel and/or pomace, 8 –11 grapes, 8,12 and passion fruit. 13 Other residues, such as wheat bran 4,14 and cashew apple husk 15 have been used as supplements.
Pectinase production and recovery by microorganisms has been studied under various conditions for solid-state cultivation processes maximization, seeking to minimize both time and cost and on maximizing its yield, productivity, and quality. Therefore, the recovery of produced metabolites is considered a major aspect since the obtained enzyme's initial recovery stage is the fermented material leaching, consisting of the preferential dissolution of one or more solutes of interest by the contact of a solid with a liquid solvent, named solid-liquid extraction. 10,16
For the development of an efficient leaching process, the influence of different variables as well as choosing the appropriate equipment should take place. Among them, solids preparation, solvent/mass ratio, process temperature, solvents pH, liquids superficial tension, system agitation level, and the occurrence or not of a chemical reaction on the extraction are to be highlighted. 10 Furthermore, the enzyme-extraction process must be performed using temperature and pH values that present great stability with equally elevated biological activity—neither damaging the biomolecules nor the solvent. 17
Therefore, the aim of this work was to optimize the process parameters to exo-polygalacturonase recovery and production from Aspergillus niger ATCC 9642 by solid-state cultivation using agro-industrial residues (orange peel, corn steep liquor, and water parboiled rice) and enzyme partial characterization.
Materials and Methods
Exo-Polygalacturonase Production in SSC
Initially, assays were carried out in order to maximize exo-PG extraction (media not optimized). Subsequently, tests were conducted varying the composition of culture medium and operating conditions in order to maximize exo-PG production. Exo-PG activity was expressed in unit (U) per gram (g) of the substrate (wet matter (wm) and dry matter (dm)).
Agro-industrial substrates
The agro-industrial substrates used were corn steep liquor (CSL), donated by Corn Products (Mogi Guaçu, Brazil), orange peel (Citrus genre—Valencia variety; juice production residue—65% moisture content) procured from a local restaurant (Erechim) and wheat bran (Olfar Ind. & Com. Óleos Vegetais Ltda., Erechim, Brazil). The agro-industrial waste remained frozen (−20°C) for a maximum 1 year period until used.
Extraction parameters evaluation
Solid-state cultivation (SSC) process has been done in a polypropylene beaker (500 mL) using 16 g ground orange peel, 2 g wheat bran, and 2 g corn steep liquor mixture. That substrate mixture was sterilized in a vertical downward autoclave (Phoenix, model AV75) at 121°C and 1 atm for 15 min. A 5 × 106 spores/gwm concentration of A. niger ATCC 9642 has been inoculated in the mixture and later incubated in a germination chamber (Tecnal, Piracicaba, Brazil; model TE401), under controlled relative humidity at 30°C for 96 h.
After cultivation period, 10 g of fermented medium diluted in solvents and/or solutions were used on the enzymes recovery process. The mixture was homogenized under controlled temperature, time period, and agitation speed. The enzymatic extract was separated from both the mycelium and the solid medium by the filtration process, followed by centrifugation (MPW Med. Instruments;Model 351R) at 4,000 rpm, for 15 min at 4°C.
At first, in order to recover the exo-PG enzyme, extractor solutions with distinct ionic forces (distilled water, NaCl 0.1 mol/L, citrate buffer 0.005 mol/L—pH 5.4, acetate buffer 0.1 mol/L—pH 5.0, and Tween 80/0.1%, v/v), at 5:1 (v:w) ratio, homogenized at 175 rpm, at 20°C for 30 min have been tested. 18
Afterward, a central composite rotatable design (CCRD) 23 was performed, aiming at optimizing and/or maximizing pectinase recovery. The following independent variables were studied: extraction temperature (13.2–46.8°C, X1), contact time (4.8–55.2 min, X2) and agitation speed (12–348 rpm, X3) using NaCl (0.1 mol/L) as extraction solvent at a 5:1 (v:w) ratio. The dependent variable (response) was the exo-PG enzymatic activity measurement (U/gwm).
Successive extraction cycles (10 in total) have been studied under the maximized conditions, with and without homogenization (20 min, 30°C, and 175 rpm). For this purpose, 25 mL of NaCl (0.1 mol/L) were added to the solid fraction during solid-liquid separation for each extraction cycle.
Optimizing exo-PG bioproduction
Culture medium composition effects on exo-PG bioproduction were assessed by a CCRD 23 (17 assays with 3 central points), orange peel (9.28–22.72 g), corn steep liquor (0.32–3.68 g) and wheat bran (0–4 g) were the independent variables investigated. The temperature (30°C), moisture content (65%), cell concentration (5 × 106 spores/gwm) and time (96 h) variables were kept fixed.
Based on the CCRD 23 results, a factorial design followed by a central composite rotatable design (CCRD) 22 was carried out. The independent variables investigated were corn steep liquor (3.7–10.3 g) and wheat bran (0–7.5 g). The orange peel (9 g), temperature (30°C), moisture content (65%), cell concentration (5 × 106 spores/gwm) and time (96 h) variables were kept fixed. The dependent variable (response) was exo-PG activity.
After the production optimization stage, substrate consumption kinetics (Total Reducing Sugar, TRS), biomass, pH evolution, and exo-PG activity were examined by periodic sampling.
Exo-PG Partial Characterization
In order to determine the optimum temperature and pH values in terms of enzyme activity, a CCRD 22 was performed using an enzymatic extract from A. niger ATCC 9642. The studied range for pH was 3.09 to 5.91 and for temperature 30 to 80°C. Crude enzymatic extract stability was measured determining the enzyme activity periodically.
Temperature stability of enzymatic extract (exo-PG maximum activity) was determined using enzyme incubation in a 4.5 fixed pH and at different temperatures: −80, −10, 4, 40, 50, 60, and 70°C. The stable pH was determined at a 3–8 range by incubating the obtained enzyme extract at 55°C for the synthetic and agro-industrial media, respectively. The samples were removed at regular time intervals. Samples at low temperatures (−80, −10 and 4°C) were stored for a maximum period of 170 days. Samples at high temperatures (40, 50, 60 and 70°C) and those of different pH were stored for 24 h.
Analytical Determinations
Exo-PG activity
Exo-PG activity has been determined by the reducing groups release measurement using dinitrosalicylic acid method (DNS), initially proposed by Miller. 19 Test tubes containing 0.8 mL of citric pectin solution (0.5% w/v, Sigma) diluted in a pre-heated water-bath (37°C for 15 min) sodium acetate buffer (200 mmol/L, pH 5.5) received a 0.2 mL supernatant aliquot (enzymatic extract), subsequently homogenized and kept in water-bath for 6 min at 37°C. 20 After the reaction period, 1 mL of DNS reagent was immediately added to each test tube, then heated (in boiling water-bath) for 8 min (enzyme denaturation and color formation), followed by an ice-cooling bath.
After that, an 8 mL double tartrate sodium-potassium solution (50 mmol/L, Vetec, Sigma Aldrich, St. Louis, MO) was added, proceeding to homogenization and absorbance measurement using a spectrophotometer (model 8453, bucket G, Agilent Technologies, Santa, Clara, CA) at 540 ηm (λ540) wavelength. An activity unit (U) has been defined as the amount of enzymes needed to release 1 μmol of galacturonic acid per minute (U = μmol/min), under reaction conditions and according to the established pattern curve, using monohydrate α-D-galacturonic acid (212.16 g/mol molecular weight, Sigma) as a reducing sugar on a 0 to 1 mg/mL range. Exo-PG activity was expressed in U/g of the substrate (wm and dm).
Total reducing sugars
The amount of total reducing sugars was estimated using the 3,5 dinitrosalicylic acid method with glucose as the standard. 19
pH
The pH was monitored using a potentiometer (DMPH-2, Digimed, São Paulo, Brazil) after calibration with standard solutions for pH 4.0 and 7.0.
Cell concentration
For cell concentration determination, the indirect method described by Scotti et al. measuring the amount of glucosamine was used. 21 For fungal chitin hydrolysis into N-acetyl glucosamine, a filter containing dried biomass was incubated at 25°C with 5 mL of 72% H2SO4 in a rotary shaker at 130 rpm for 30 min. After dilution with 54 mL of distilled water, hydrolysis was performed by autoclaving the mixture for 2 h at 121°C. The hydrolyzate was neutralized to pH 7.0 with 10 M and then with 0.5 M NaOH using a pH meter (Corning, Corning, NY). Finally, glucosamine was assayed by the colorimetric, in a spectrophotometer (Agilent Technologies, model 8453) at 530 ηm wavelength. For indirect glucosamine concentration determination, reducing sugar was used in a 0–0.05 mg/mL range. For fungal biomass (mg/g fermented medium) determination. Glycosamine value from A. niger culture was determined in the medium at 0 to 0.5 g of dry biomass concentrations.
Statistical Treatment
The other results were treated by variance analysis (ANOVA), followed by Tukey's test. All the analysis were performed using Statistica version 5.0 software (Statsoft Inc, Tulsa, OK).
Results and Discussion
Exo-Polygalacturonase Extraction Parameters Evaluation
Table 1 presents exo-PG activity values recovered from the fermented medium using different solvents (distilled water and NaCl) and extracting solutions (citrate buffer, acetate buffer, and Tween).
Activity of Exo-PG Enzyme Using Different Types of Solvents and Solutions in the Process of Extraction of Pectic Enzymes
Solvents (distilled water, NaCl 0.1 mol/L, citrate buffer 0.005 mol/L, pH 5.4, acetate buffer 0.1 mol/L, (pH 5.0) and Tween 80 (0.1°%, v/v)), at 5:1 (v:w) ratio, homogenized at 175 rpm, at 20°C for 30 min. Same letter between lines indicate the statistically significant difference at the level of 5% between lines (Tukey test).
NaCl solvent (0.1 mol/L) was efficient on exo-PG extraction obtaining 10 U/gwm (29 U/gdm). In this way, sodium chloride (0.1 mol/L) was established as the extracting solvent for the subsequent stages. Similar results were observed by Díaz et al., 18 where exo-PG maximum recovery was 42 U/gdm using NaCl (0.1 mol/L).
Table 2 presents a DCCR 23 (real and coded values) on the enzymes recovery and on exo-PG activity response. The noted maximum exo-PG recovery was 10 U/gwm (28 U/gdm) using NaCl 0.1 mol/L, 30°C, for 30 min at 180 rpm (Run 16).
Matrix of DCCR 23 (Real and Coded Values) with the Responses of Exo-PG Recovery (U/gwm)
X1 = Temperature (°C), X2 = time (min), X3 = Agitation (rpm); Independent variable fixed: extraction solvent (0.1 mol NaCl/L). Fermentation conditions: 16 g orange peel, 2 g wheat bran and 2 g corn steep liquor, 65% humidity, pHinitial 4.3, 30°C, 5.106 spores/gwm and 96 h.
Planning results were treated statistically and Equation 1 presented the second order coded models, describing the exo-PG activity. Non-substantial factors were added to the lack of adjustment for the variance analysis-ANOVA, the following coefficients were obtained: 0.88 correlation coefficients and the calculated F value was 1.8 times greater than the F tabulated one for exo-PG. Thus, validating the models and allowing contour curves plotting as seen in Fig. 1a.
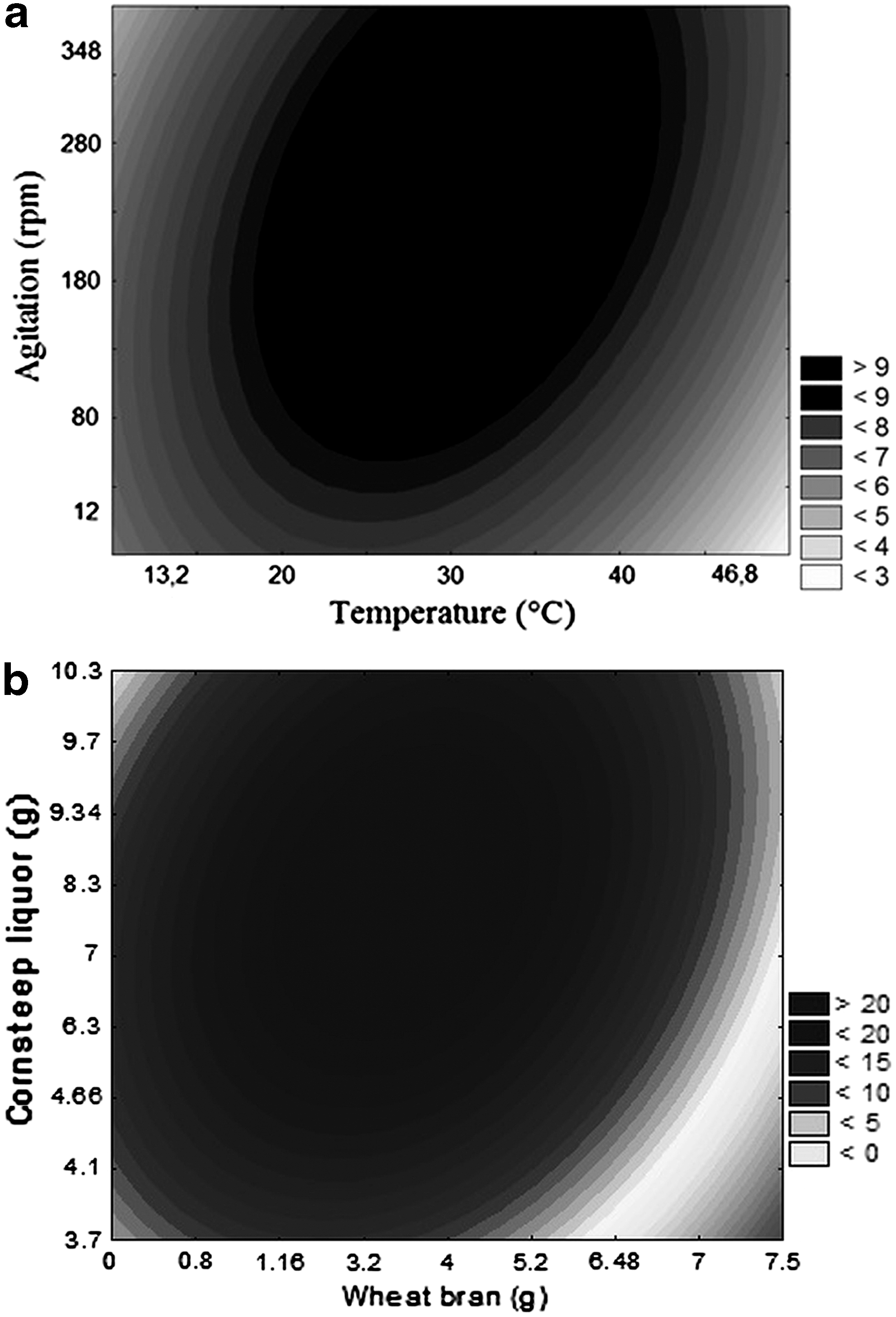
where exo-PG is exo-polygalacturonase activity (U/gwm), X1 is temperature (°C) and X3 is agitation speed (rpm).
According to Fig. 1a, an exo-PG recovery optimization region of approximately 10 U/gwm exist in the 25–35°C and 180–300 rpm ranges. Some studies on the literature evaluated different parameters on pectinolytic enzymes recovery. 3,10,13,15,22 It is important to mention the temperature range obtained during the enzyme-recovery optimization process (25–35°C) is noteworthy on industrial activities, as it corresponds virtually to room temperature, not requiring any additional energy expenditures during extraction stage.
On the first extraction cycle, recoveries of approximately 9 U/gwm (26 U/gdm) for exo-PG were obtained from 10 extraction cycles (data not shown). Pectinase yield reduced significantly (p < 0.05) on the second cycle and was subsequently kept practically constant (p > 0.05). Similar works have been mentioned in the literature. Castilho et al. assessed the evolution of pectinases concentration produced by A. niger on extracts originated from successive rinsing and observed a significant decrease in enzymatic activity on each new extraction, providing fairly diluted extracts and next to zero activity on the final solvent passages. 22
Díaz et al. evaluated exo-PG extraction efficiency from the fermented grape pulp using distilled water as solvent, indicating that more enzymes (exo-PG) could be separated from the fermented solid after the sixth extraction stage. 18 Singh et al. assessed PG (A. carbonarius) recovery parameters and noted that after the third cycle, they increased from 10 to 42 U/mL presenting a result proportionally superior to the one found in this study that was 10 to 13 U/gwm. 23 Rodríguez-Fernández et al., on their studies regarding PG enzyme (A. niger F3, 96-h fermentation) successive extraction (6 cycles), observed the possibility to recover 70% of all enzymes in three extraction cycles. 10
Exo-PG Optimization Production
Table 3 presents both coded and real values of the complete 23 factorial design and the response in terms of exo-PG activity. Maximum exo-PG production was 15.5 U/g wm (44 U/g dm ) in a medium composed of 74.1% orange peel, 9.3% wheat bran and 16.6% corn steep liquor (run 14), 65% moisture content, 5 × 106 spores/gwm pHinitial of 4.3, at 30°C for 96 h.
Matrix of the CCRD and Response as Exo-PG Production
X1 = Orange peel (g), X2 = Wheat bran (g), X3 = Corn steep liquor (g). Independent variables fixed: 5.106 spores/gwm, moisture content of 65%, at 30°C for 96 h; bX2 = Wheat bran (g), X3 = Corn steep liquor (g). Independent variables fixed of the orange peel (9g), 5.106 spores/gwm, moisture content of 65%, at 30°C for 96 h.
This fact can be better observed (Pareto chart of effects—data not shown) where wheat bran and corn steep liquor variables presented a positive significant effect (p < 0.05), demonstrating the displacement of such variables to upper levels could lead to an increase in enzyme activity. The orange peel variable presented a negative significant effect (p < 0.05). Considering that orange peel is the major inducer for pectinase formation, 24 the peeling concentration could influence the compaction medium and mass transfer due to porosity between particles. The lower level of these variables was 9 g (−1), so they were fixed from the culture medium in the sequential experimental design.
A 22 full experimental design was performed in order to maximize enzyme production. Results from this step are presented in Table 3. It can be verified that maximum exo-PG production was 20 U/g wm (58 U/g dm ) in the medium composed of 9 g orange peel, 4 g wheat bran and 7 g corn steep liquor (runs 9, 10 and 11), 65% moisture content, 5 × 106 spores/gwm, pHinitial of 4.5, at 30°C for 96 h.
The statistical analysis of these results allowed the development of a second-order coded model (Equation 2), which described the exo-PG activity as a function of the analyzed independent variables (corn steep liquor and wheat bran), within the studied range. The model was validated by variance analysis with a 0.97 correlation coefficient and the F calculated value was 2.92 times greater than the one listed in statistical tables. The empirical model was used for the contour curve plotting presented in Fig. 1b.
The maximum exo-PG activity was 20 U/gwm (58 U/gdm), which maximizes exo-PG bioproduction (Fig. 1b) in the agro-industrial medium at a 9 g orange peel concentration in a 4 to 7 g range of corn steep liquor and wheat bran, respectively.
Where exo-PG is exo-polygalacturonase activity (U/g wm ), X2 is wheat bran and X3 is corn steep liquor.
Figure 2 presents the kinetic evaluation of exo-PG activity, biomass production, substrate consumption (reducing sugar), and the optimized conditions pH. Exo-PG maximum activity (Fig. 2a) was 23 U/g wm (68 U/g dm ), obtained after 192 h of bioproduction. Reducing sugar concentrations (Fig. 2b) decreased 83% up until 96 h of cultivation.
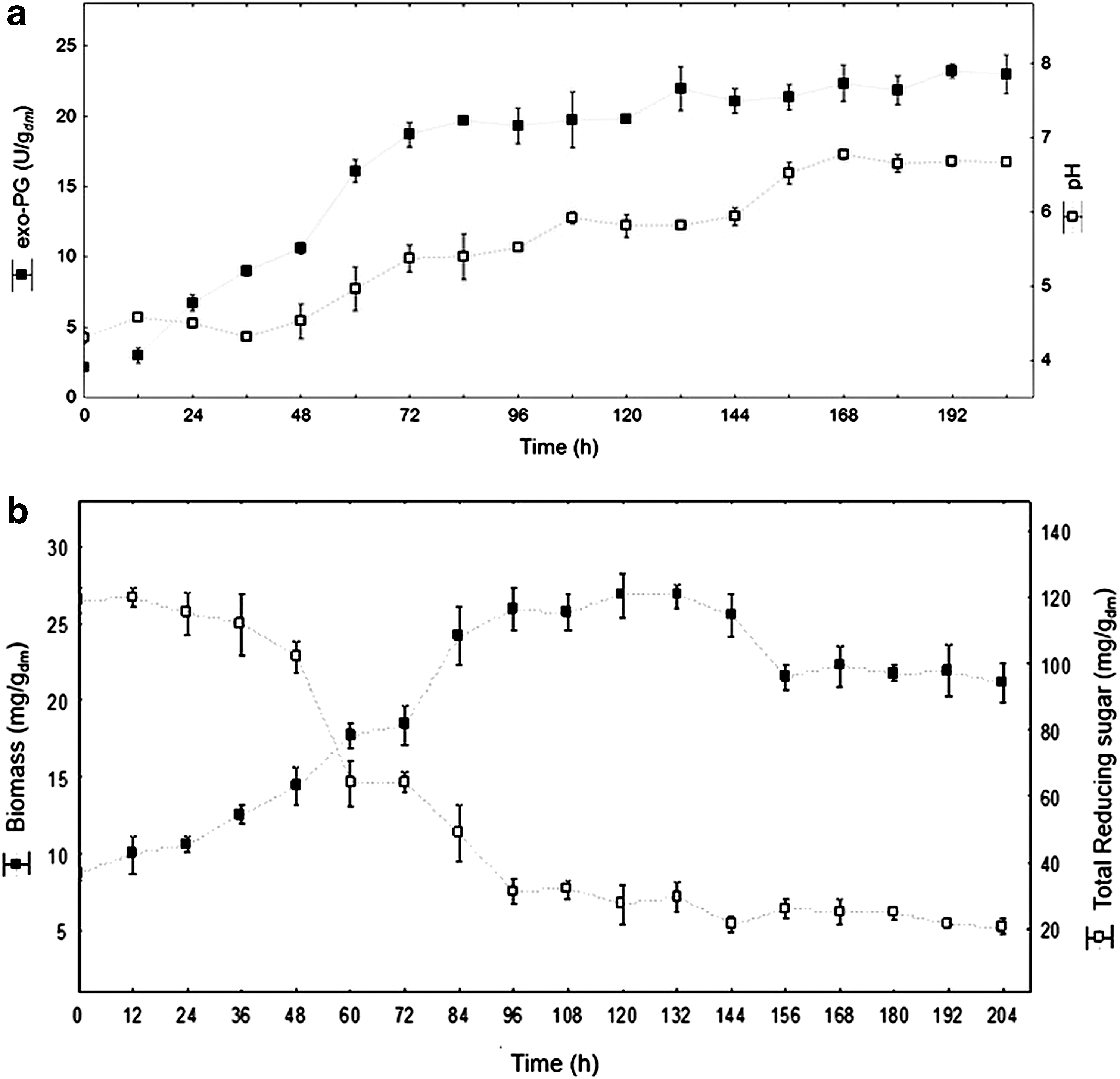
Kinetic curves for bioproduction of the optimized conditions
Mata-Gómez et al. evaluated pectinase activity from Aspergillus sojae ATCC 20235 in a medium containing beet sugar as a carbon source and obtained the maximum production (123 U/g) in 8 days. 25 Silva et al. obtained the maximum PG activity of 55 U/g after an 8-day cultivation period using orange peel residue and wheat bran (1:1, w:w) ratio, in solid-state cultivation by Penicillium viridicatum. 26 Heerd et al. obtained the maximum activity of 28 U/gdm using a mixture of orange peel:wheat bran (70:30, w:w) ratio in pectinase production by Aspergillus sp. 27
Fermentation medium pH (Fig. 2a) showed a slight decline in the first 36 h (pH 4.2), and a progressive increase up to 204 h (pH 6.8) occurred after 72 h. Similar behavior was also noted in other studies. 28 –30 It is possibly associated with ammonia release in the medium resulting in protein breakdown and oxidation of amino acids of the carbon sources used in pectinase and A. niger production.
Biomass reached its maximum of 27 mg/gwm (77 mg/gdm) after 84 h culture (Fig. 2b). The curve behavior presented a classical profile since the adaptation phase in the first 24 h of exponential multiplication of 24–84 h and the stationary phase after 84 h took place. Hendges et al. evaluated biomass during the endo-PG production from A. niger (T0005/007-2) and the maximum value after 72 h was 100 mg/gdm. 31
Other reports in the literature can be observed on bioproduction kinetics. Diaz et al. found that sugar concentration decreases drastically after 120 h of fermentation employing the substrate as orange peel and an orange peel and apple pomace mixture. 8 The pH decreased from 4.0 to 3.5 in the first 3 h, and increased and maintained itself at a constant 4.8 after 4 h. Exo-PG maximum activity was 17 U/g after 14 days using orange peel.
Souza et al. used passion fruit residue (40% moisture and 1% nitrogen source) in the production of pectinases production by A. niger. 32 The pH remained nearly constant (pH 4.9) during the 72-h solid-state culture. The maximum PG activity was 21 U/gwm, after 66 h.
Fontana et al. evaluated the production of exo-PG from A. niger (T0005007-2) using a substrate with wheat bran base as a carbon source, and the maximum activity was 232 U/gwm at 48 h. 33 The pH decreased from 5.0 to 4.1 and increased to 5.2 and remained unchanged after 48 h during the exponential phase.
Exo-PG Partial Characterization
Optimum temperature and pH
Optimum pH and temperature evaluation was performed using a CCRD 22. Table 4 presents the experimental design matrix with the studied variables ranges and the responses in terms of exo-PG activity. The highest activity was 26 U/g wm (75 U/g dm ) (runs 9, 10, and 11) at 55°C and pH 4.5.
Matrix of 22 CCRD (Real and Coded Values) of the pH and Temperature Optimum with the Responses of Exo-PG activity (U/gwm)
X1 = pH, X2 = temperature (°C).
These results were statistically treated and empirical models were obtained (Equation 3). This model represents exo-PG activity behavior in terms of temperature and pH over the studied range and were validated using variance analysis-ANOVA, which showed a 0.97 correlation coefficient and F calculated values 3.55 times greater than the tabulated ones—as shown in the contour curve plotting in Fig. 3 that demonstrates the maximum exo-PG production occurs around pH 4.5–5.5 and temperatures of 40–60°C. These results corroborate with those obtained by other authors on polygalacturonase production by A. niger.
34
–39
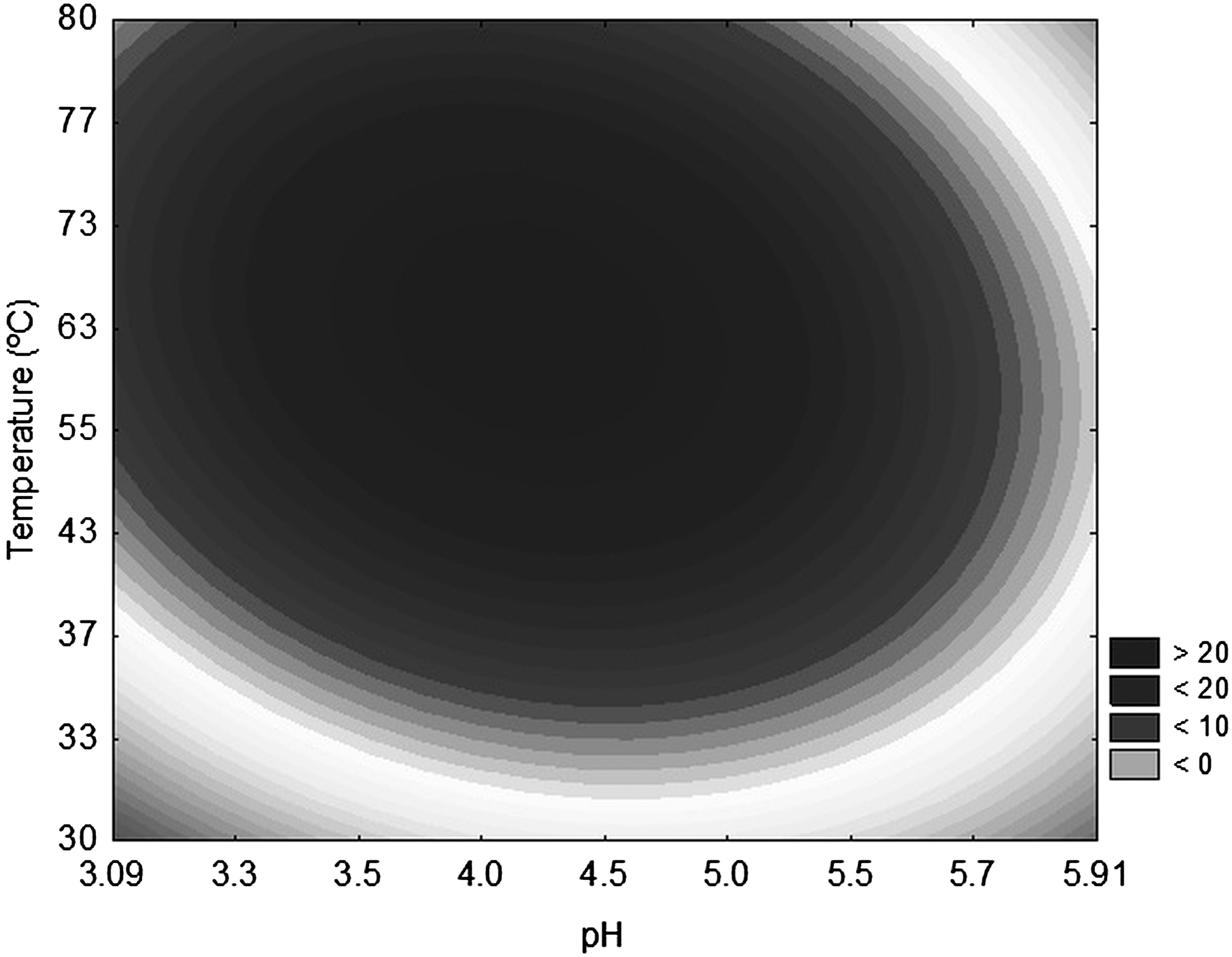
Contour curve for exo-PG as a function of temperature (°C) and pH.
where exo-PG is exo-polygalacturonase activity (U/gwm), X1 is pH and X2 is temperature (°C).
Crude enzymatic extracts temperature and pH stability
It was revealed, from the stability evaluation results at 40, 50, 60, and 70°C (Fig. 4a) that crude enzyme extract showed greater stability at 40°C, keeping 80% of its original activity for 24 h. It was observed from stability evaluation results at low temperatures (Fig. 4b) that when the crude extract was stored at −80° and −10°C, it remained 70% stable at 4°C for 21 weeks and 14 weeks, respectively. The stability of crude exo-PG enzyme extract to a different pH level was performed by monitoring the behavior as a time function. The results obtained are presented in Fig. 4c, and it was verified that crude enzyme extract presented the greatest stability at pH 3.0 to 5.0, retaining 70% of its initial exo-PG activity, after 1 h storage. Maller et al. found stable PG (A. niger) values at 60°C for 90 min and pH at 3 to 5. 37
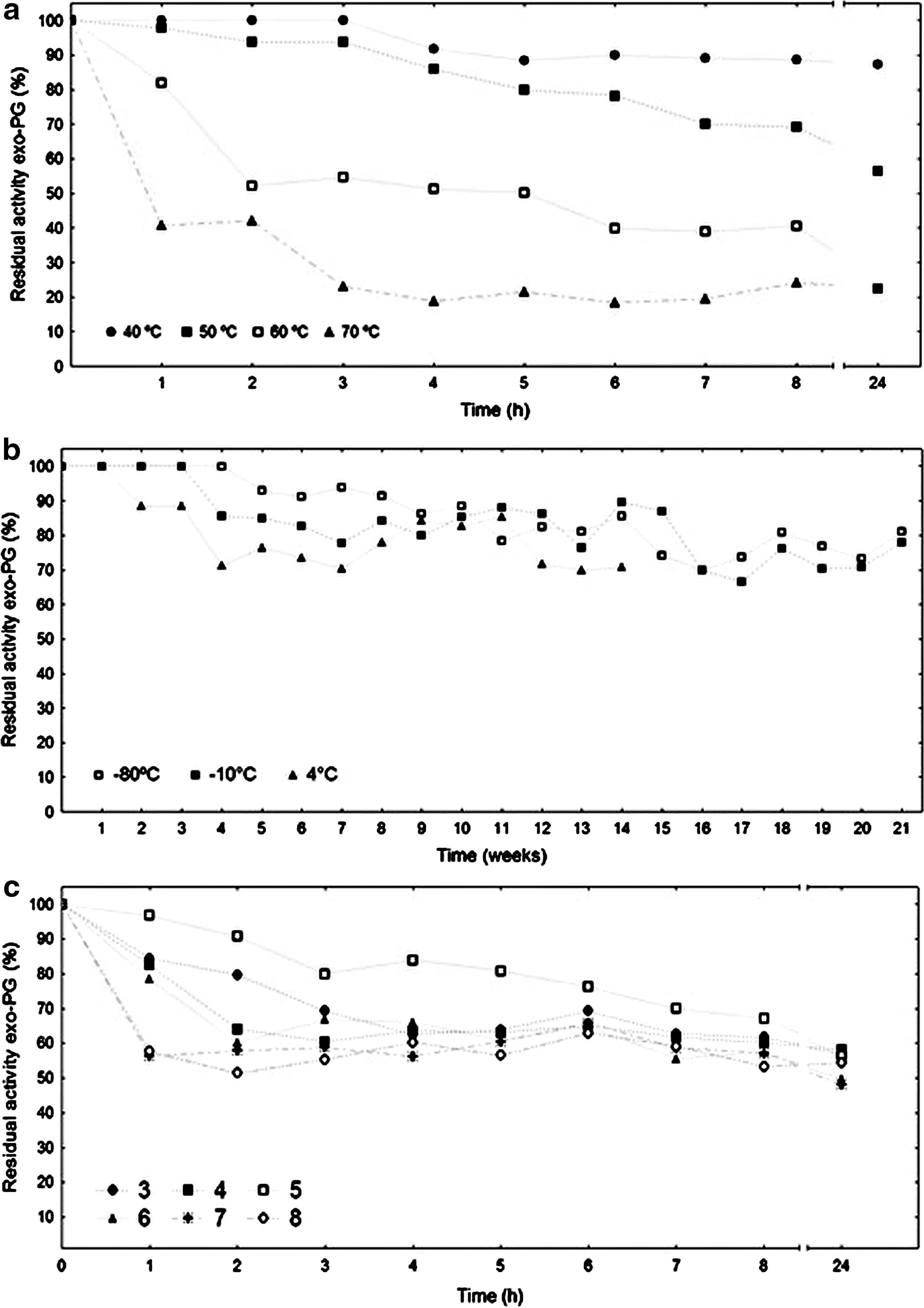
Influence of
Conclusion
The maximum exo-PG recovery was 9 U/gwm (26 U/gdm) using NaCl (0.1 mol/L) as solvent at a 5:1 (v:w) ratio, 30–55 min time period, 180 rpm agitation, and in one extraction cycle. In our work, the maximum exo-PG activity was 23 U/g wm (68 U/g dm ) obtained after 192 h of bioproduction with a 9 g orange peel, 4 g corn steep liquor, and 7 g wheat bran concentration, at 30°C, and 5x106 spores/gwm. The optimum crude enzyme extract pH and temperature were 4.5 and 55°C, respectively. The enzyme extract retained 70% stability of enzyme activity stability when stored at −80° and −10°C for 21 weeks at pH 3.0–5.0 for 1 h, respectively. The enzymatic extract shows potential applications for acidic foods (e.g., fruit juices).
Footnotes
Acknowledgments
The authors would like to thank the National Council for Scientific and Technological Development (CNPq), Coordination for Improvement of Higher Education Personnel (CAPES) and the Rio Grande do Sul State Research Support Foundation (FAPERGS).
Author Disclosure Statement
No competing financial interests exist.