
Abstract
To produce energy for in-house use, an aluminium smelter in Québec launched a study of mixotrophic cultivation of microalgae in its wastewaters with the objective of having an algae production company set up operations on site. To maximize lipid productivity and maintain the biological integrity of the consortium, specific nutrients need to be added to aluminium smelter wastewaters to cultivate the selected algae-bacteria consortium. A 23 factorial design was used to determine the organic carbon, nitrogen and phosphate inputs needed. Data on biomass and lipid productivity, as well as a “consortium integrity index,” were analyzed using a multiple linear regression model. The highest biomass productivity (0.93 g/L/d) and lipid productivity (0.023 g/L/d) were obtained using the highest tested concentration in nitrogen (0.200 g/L) and the lowest tested concentration in phosphate (0.003 g/L). No significant effect of the organic carbon—tested in concentrations of 1.64 g/L, 2.64 g/L, and 3.64 g/L (glucose) and added in two increments on days 5 and 7—on productivity for a starting cell density of 5 million cells/L was detected. To achieve maximal lipid production, the results suggested that biomass productivity should be prioritized rather than lipid accumulation in the cells through nitrogen starvation. The stability and integrity of the cultured consortium have to be maintained through an appropriate balance of nutrients. A low phosphate concentration increases the stability of the consortium, although part of the variation cannot be explained by the model. Finally, an analysis of fatty acid profiles showed that different concentrations in nitrogen and phosphorus impact the proportion of C18:1(n-9) and other minor fatty acids.
Introduction
In the current environmental and global energy context, aluminum producers are interested in using renewable sources of energy to reduce their environmental footprint and secure their energy supply. Production of algal biomass rich in lipids is a potential option. Microalgae offer high productivity per surface unit compared with other industrial plant crops and can be converted to valuable fuel or other forms of energy. 1 Some examples of such renewable energy sources are the production of biogas via methanation, direct combustion, ethanol production through the fermentation of microalgae sugars, biodiesel production by oil extraction and transesterification, and biocrude oil production through hydrothermal liquefaction. 2 –6
On the other hand, microalgae production in co-location with an industrial plant can improve the profitability and environmental benefits of the overall process. 7,8 Recycling of wastewater and other wastes that contain nutrients as well as the use of residual heat and effluent gas from industrial processes can reduce microalgae production costs. 9 –12
Many factors come into play in selecting the growth pattern and culture system. In the specific case of this study, the energy production objective and co-location with an aluminium smelter must be considered. Production under mixotrophic conditions appears to be the most convenient metabolic approach in this context. First, it promotes the fixation of industrial effluent carbon dioxide (CO2) by photosynthesis (autotrophy) and reduces greenhouse gas emissions. 5 Second, because photosynthesis may not always be optimal in areas such as Canada during the winter season, heterotrophic growth may assist in overcoming light limitation. The metabolization of organic carbon (heterotrophy) from industrial wastewaters and residues (nutrient wastes) and use of waste energy from the plant can maintain productivity year round while creating economic value from waste. 13 Mixotrophy could also be more effective in producing lipids than either heterotrophy or autotrophy. 14 The use of photobioreactors as a culture system can result in excessively high operational expenditure (OPEX) for the production of low-cost commodities such as biofuel. The use of open ponds could therefore be the best option. 15
Carbon (C), nitrogen (N) and phosphorus (P) are major constitutive elements of almost all living organisms. 16 For microalgae cultivation, these elements must be present in the culture medium, in keeping with the needs of the specific strain. The concentrations of those nutrients will impact biomass productivity and lipid accumulation in the cells. It is known that addition of a sufficient amount of carbon and nitrogen maximizes the production of biomass, while nitrogen starvation triggers neutral lipid accumulation. 17,18 The nutrient ratio also affects the fatty acid profile. For example, nitrogen deprivation increases the percent of oleic acid (C18:1 ω-9) in several species. 19 A balance between biomass production and cellular lipid concentration is essential to achieve optimal production of valuable lipids.
Mixotrophic microalgae cultivation in open ponds requires the right nutrients concentration and balance to ensure algae growth and control contamination from unwanted microorganisms (fungi, protists, rotifers, undesired bacteria, and algae) that compete for food. Several ways to reduce competition and maintain the stability in algae cultures are reported in the literature: (1) increasing the microalgae inoculum size, 20 (2) using a consortium rather than a pure strain, 21 (3) adding a small quantity of organic carbon by fed-batch rather than a large by batch, 22 and (4) maintaining an alkaline pH of the culture medium. Koller et al. also observed that culture medium containing a high nutrients concentration limits the development of undesired microorganisms. 23 Nutrients concentration and balance can thus affect microalgae productivity indirectly by increasing or reducing contamination.
In the context of this study, an algae-bacteria consortium, mainly Chlorella sp., was isolated from aluminium smelter wastewaters. The consortium was cultivated under mixotrophic conditions, in the same wastewater, to obtain algal cells with a high lipid content. Because smelter wastewater does not contain enough nutrients to support optimal growth of the consortium, supplements were added. To keep production cost low, local industrial and agricultural food wastes can be used to replace conventional nutrient sources. However, tests with conventional nutrients have to be done at the outset to determine the consortium's actual requirements for each element, and avoid any interference with other compounds.
The purpose of this study was to determine appropriate concentrations of the main nutrients (C, N, and P) needed to achieve high lipid productivity and to observe the effect they have on consortium biomass production. To compare the stability of the strain under particular conditions (C, N, P concentrations), an integrity index for the consortium was measured. The consortium integrity index is based on the proportion of Chlorella sp. in relation to the total number of observed microorganisms (including any undesired organisms). Fatty acid profiles were also analyzed to evaluate the effects of C, N and P concentrations on oil quality for further conversion into biodiesel by transesterification.
Materials and Methods
Strain and Culture
The algae-bacteria consortium cultivated in this study is a native strain, dominated by a green Chlorella sp. that was isolated from aluminium smelter wastewater, by enrichment of samples followed by directed selection. The same aluminum smelter effluent-based culture medium was used in this study. The wastewater is non-potable freshwater that contains few solids (1.2–3.6 mg/L), with metals present in concentration meeting environmental standards. The seed culture was cultivated in an aluminum smelter effluent-based medium, to which Bold Basal medium nutrients were added 24 with the following modifications: KNO3 0.750 g/L, KH2PO4 0.700 g/L, MgSO4·7H2O 0.300 g/L, and sodium ferric EDTA 0.028 g/L. Glucose was added as an organic carbon source in fed-batch mode, with an initial 0.5 g/L provided at day 5 and a second 0.5 g/L provided at day 7.
The inoculum used in this study was collected from a seed culture in exponential growth phase. It had a concentration of 5 × 107 algae cells/mL and represented a volume of 10% (v/v) of the medium, for a starting cell culture of 5 × 106 algae cells/mL. Cultures were grown in 1-L Erlenmeyer flasks containing 500 mL of medium. They were agitated using an orbital shaker at 130 rpm, under fluorescent lighting 12 h/day with an intensity of 20 μmol photons/m/s (measured by a LI-COR Model LI-250 photometer, Lincoln, NE). The temperature was controlled at 21°C and the pH was measured on a daily basis (pH meter Symphony, SB70P model VWR, Radnor, PA). CO2 supply was from the air, by exchange through the foam cap.
The experiment was carried out with organic C, N, and P at three different concentrations (Table 1), based on previous unpublished work. For Mg and Fe, the same nutrient concentration as seed culture was added. All nutrients were added in the medium at day 0, except glucose. The culture was grown for 9 days, including a 4-day acclimation period and an exponential growth period starting after the first glucose injection on day 5. A second injection of the same glucose concentration was done 48 h after the first one, on day 7. The C concentrations at levels −1, 0, and 1 (Table 1) correspond to a total addition of glucose of 1.64 g/L, 2.64 g/L, and 3.64 g/L, respectively.
Factors and Levels of the 23 Factorial Design
Biomass Concentration and Productivity
The Borowitzka and Moheimani method was modified and used to measure biomass concentration (dry weight). 25 Ten mL of final culture were filtered on a previously weighed glass microfiber filter (Whatman 934-AH, porosity 1.5 μm, Maidstone, United Kingdom) and washed with demineralized water. Filters were dried for 24 h at 60°C and weighed again.
Productivity was calculated for the exponential growth period according to the following equation (1):
Pb = Biomass productivity (g/L/d)
Bf = Final biomass (g/L)
Bi = Initial biomass (g/L)
T = Total days of culture (culture period)
Neutral Lipids Concentration and Productivity
Neutral lipids were extracted from the end culture biomass and quantified using a gravimetric method.
26
Microalgae biomasses from each culture were washed with demineralized water and vacuum dried with a Savant Speedvac System, model SS21 (Thermo Savant, Waltham, MA). 0.05 g per sample of dry biomass was crushed in liquid nitrogen, soaked overnight in 5 mL of hexane, and ultra-sonicated (20hz) using a sonifier with a microprobe (Sonifier 350 Cell Disruptor and A3-561 microprobe from Branson Sonic Power Co., Danbury, CT) in order to break down algal cells. Cell fragments were separated from the liquid phase by centrifugation (5000G, 10 min). Oil was weighed after complete evaporation of the solvent. Every measurement was completed in duplicate. Neutral lipid productivity was calculated as follows (2):
Pl = Neutral lipid productivity (g/L/d)
Pb = Biomass productivity (g/L/d)
L = Neutral lipids in g per g of biomass
Consortium Integrity
Consortium integrity ensured strain stability. It is an important variable to measure because loss of strain stability can affect productivity. Consortium stability may be threatened by an undesired microorganism ratio. Algal cells were counted daily using a Neubauer chamber (hemocytometer) and a phase-contrast microscope (Axio Scope A1 from Zeiss, Toronto, Canada) in accordance with the method described by Guillard and Sieracki. 27 Undesired microorganisms, regarded as biological contaminants, i.e., algae other than Chlorella sp., protists, and rotifers, were also counted. Because the fungal hyphae could not be individually counted, their number was estimated by dividing the surface they covered on the Neubauer chamber by the average Chlorella sp. surface area (20 μm2) to produce an “algal cell equivalent” count. Bacteria level, assessed by a qualitative ranking index, is not reported because it appeared stable throughout the experiment in all trials.
The integrity index was calculated as follows (3):
I = Integrity index
Ci = Total number of biological contaminants in Neubauer chamber
Ai = Total number of consortium algae in Neubauer chamber
T = Total days of culture
i = Day of culture
Fatty Acids Profile
Fatty acids were analyzed with gas chromatography–mass spectrometry (GC-MS) using the method proposed by Li et al. 28 Dry biomasses used for the analysis were stored for one year at −20°C. 2.5 mL of a BF3: MeOH (14% V/V) solution were added to the previously extracted lipids and heated (65°C, 20 min) in closed tubes. One mL of saturated NaCl and 1 mL of hexane were then added to each tube. The tubes were shaken and centrifuged, and the supernatant was collected for analysis. An analysis of fatty acid methyl esters (FAME) was done using an Agilent (Santa Clara, CA) 7820A gas chromatograph with a DB-WAX capillary GC column (30 m, I.D. 0.25mm, film thickness 0.25mm) connected to an MS (Agilent, model 5977E). During mass spectra registration, the injection temperature was 90°C, and helium was used as a carrier (10.5 PSI, 90°C). Electron ionization was 70eV, the frequency 2.5 readings per s, and the range 50–650 m/z. Temperature, initially 100°C, was increased by 10°C/min for 2 min, and by 5°C/min until it reached 250°C. FAME identification and quantification was done using mass spectra standard Supelco® 37 Component FAME Mix (Bellafonte, PA).
Experimental Design and Statistical Analysis
SAS JMP 10.0 software (SAS Institute Inc., Cary, NC) was used to develop a 23 factorial design with 3 central points based on organic C, N, and phosphorus P factors (Table 1). Data for three response variables, i.e., biomass productivity (g/L/d), neutral lipid productivity (g/L/d) and consortium index integrity, were analyzed using multiple linear regressions with the same software. Finally, multiple correlation was used to analyze three more variables: the percent of lipids in biomass (% W/W), the initial pH, and the final pH.
Results and Discussion
Experimental Design
To determine the concentrations of organic C, N, and P that generate maximal productivity, tests with eleven combinations of factors (C/N/P) were run in accordance with a 23 factorial design, including 3 central points (tests 2, 3, and 4; Table 2). These central points were used to measure the experimental error and to validate the models. The models were performed on response variables for biomass productivity, neutral lipid productivity and consortium index integrity, to determine whether ideal concentrations for lipid productivity also favoured biomass production and consortium stability (Table 2). Because the design central points for N and P were not exactly the middle value of the two levels, confusion in terms might have occurred. However, values in the alias matrix were 0 or close to it, showing that there was no confusion among terms (Table 3).
23 Factorial Design Matrix with Observed Values
Aliases Matrix
Int: intercept.
Growth Kinetic
The growth kinetic of the 11 trials is represented in Fig. 1. For most trials, there is a lag phase of 5 days, after which the consortium start its exponential growth, following a weak decrease between the day 5 and 6 for the trials 2, 4, 6, 8, 9. Only trial 10 continued to decrease until day 7 and does not reach its exponential growth phase.
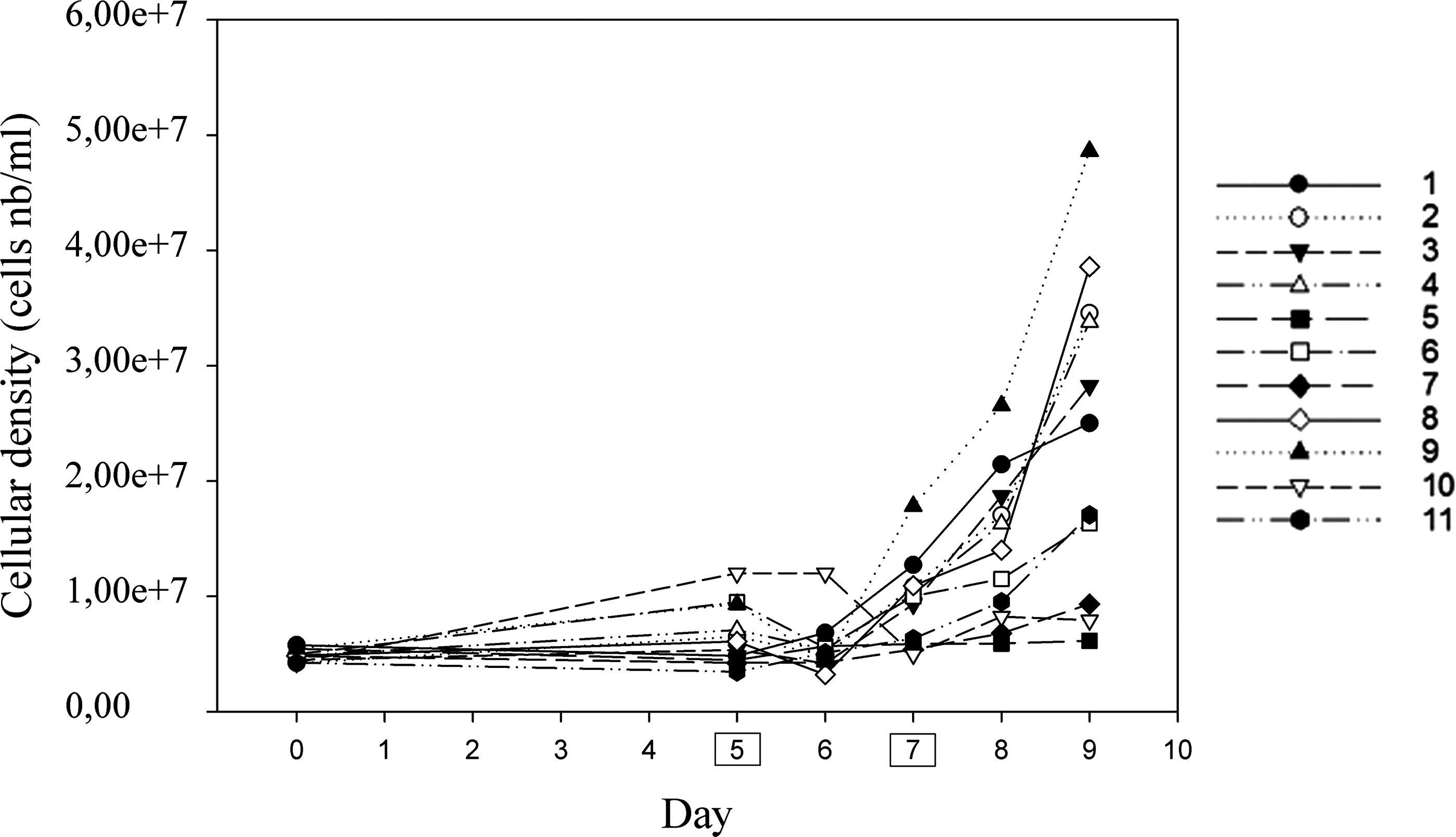
Growth kinetic of the eleven trials, measured on a cellular density basis. Days framed in a box on the x-axis correspond to the glucose fed days.
Biomass Productivity
Results of the multiple linear regressions performed on the algae-bacteria consortium biomass productivities are shown in Table 4. Main variations are on the N and P factors. The parameter estimated for C was not significant (p = 0.10, data not shown), and this factor was excluded from the model. The highest biomass productivity under our test conditions was 0.93 g/L/d using 0.200 g/L nitrogen and 0.003 g/L phosphorus. Increasing of nitrogen concentration increased biomass productivity, whereas, increasing the phosphorus concentration decreased it (Fig. 2).
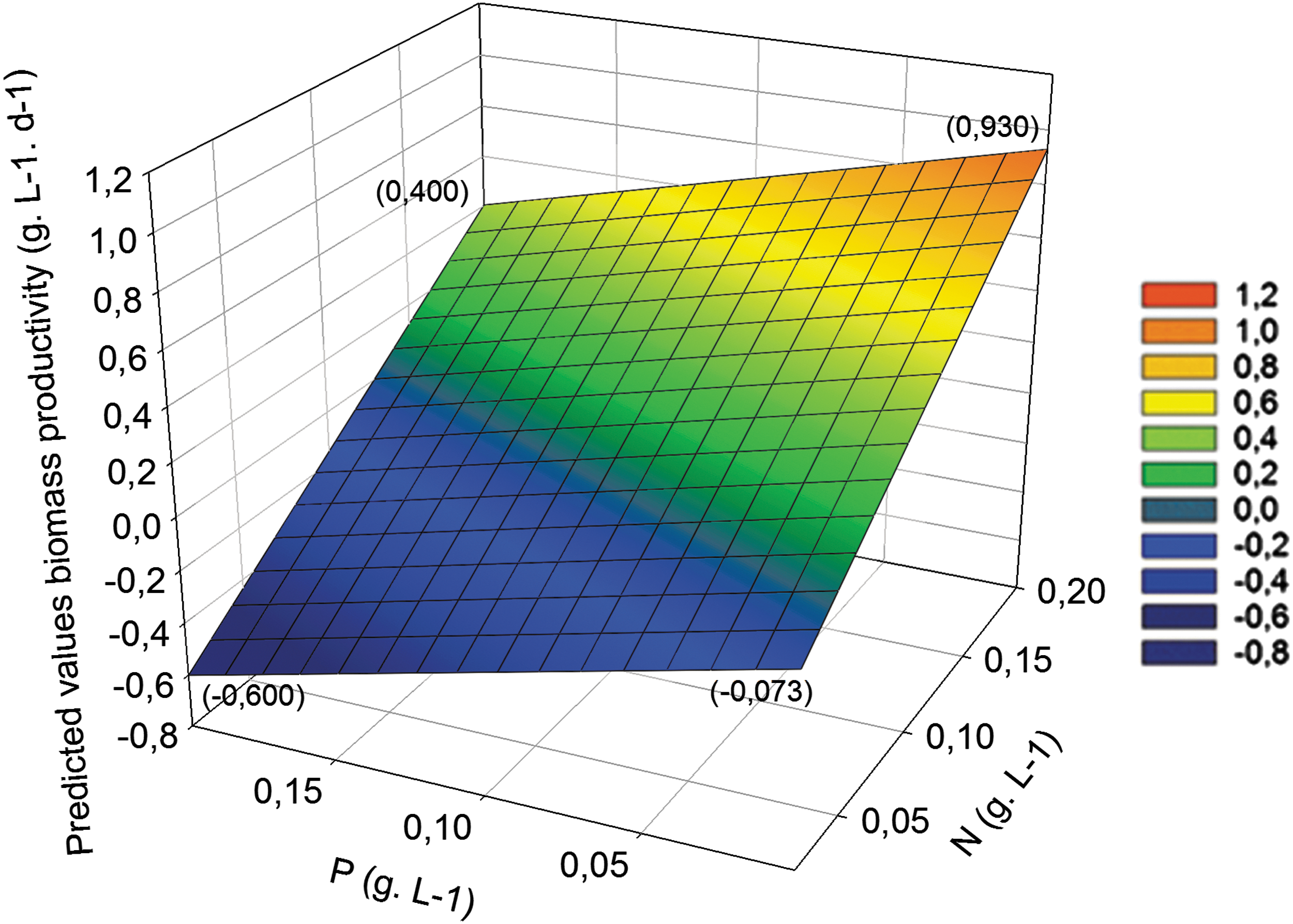
Surface profiler of the model of consortia biomass productivity according to nitrogen (N) and phosphorus (P) concentrations. In parenthesis: predicted values at the surface limits. Legend presents the predicted biomass productivity (g/L/d) values associated to the color of the surface.
Model of Consortium Biomass Productivity
Df: degree of freedom.
Lipids Productivity
In the range of nutrient concentrations tested (Table 1), N and P were the main factors that affected lipid productivity as well as the linear interaction between these two factors (Table 5). The parameter estimated for C was again not significant (p = 0.10, data not shown), and C was excluded. The highest lipid productivity in our tests conditions was 0.023 g/L/d using 0.200 g/L nitrogen and 0.003 g/L phosphorus (Fig. 3).
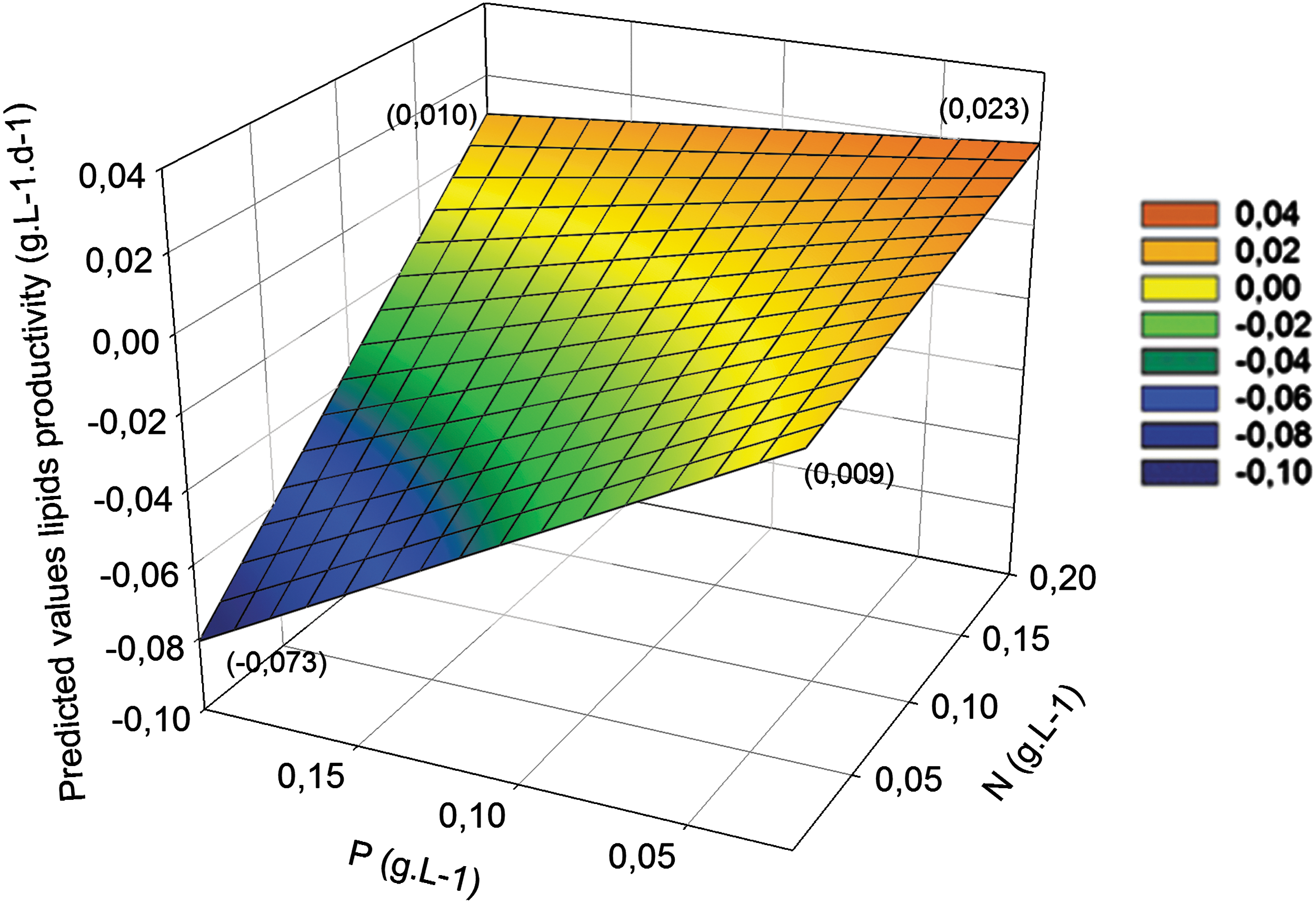
Surface profiler of the model of consortia lipids productivity according to nitrogen (N) and phosphorus (P) concentrations. In parenthesis: predicted values at the surface limits. Legend presents the predicted lipids productivity (g/L/d) values associated to the color of the surface.
Model of Consortium Lipids Productivity
Df: degree of freedom.
Integrity Index
Under test conditions, the algae-bacteria consortium had a higher integrity index when P concentration was low. Phosphorus was the only predictor used because the parameter estimates for C and N were not significant (data not shown) (Table 6). Therefore, consortium integrity decreases or increases with phosphorus concentration (Fig. 4).
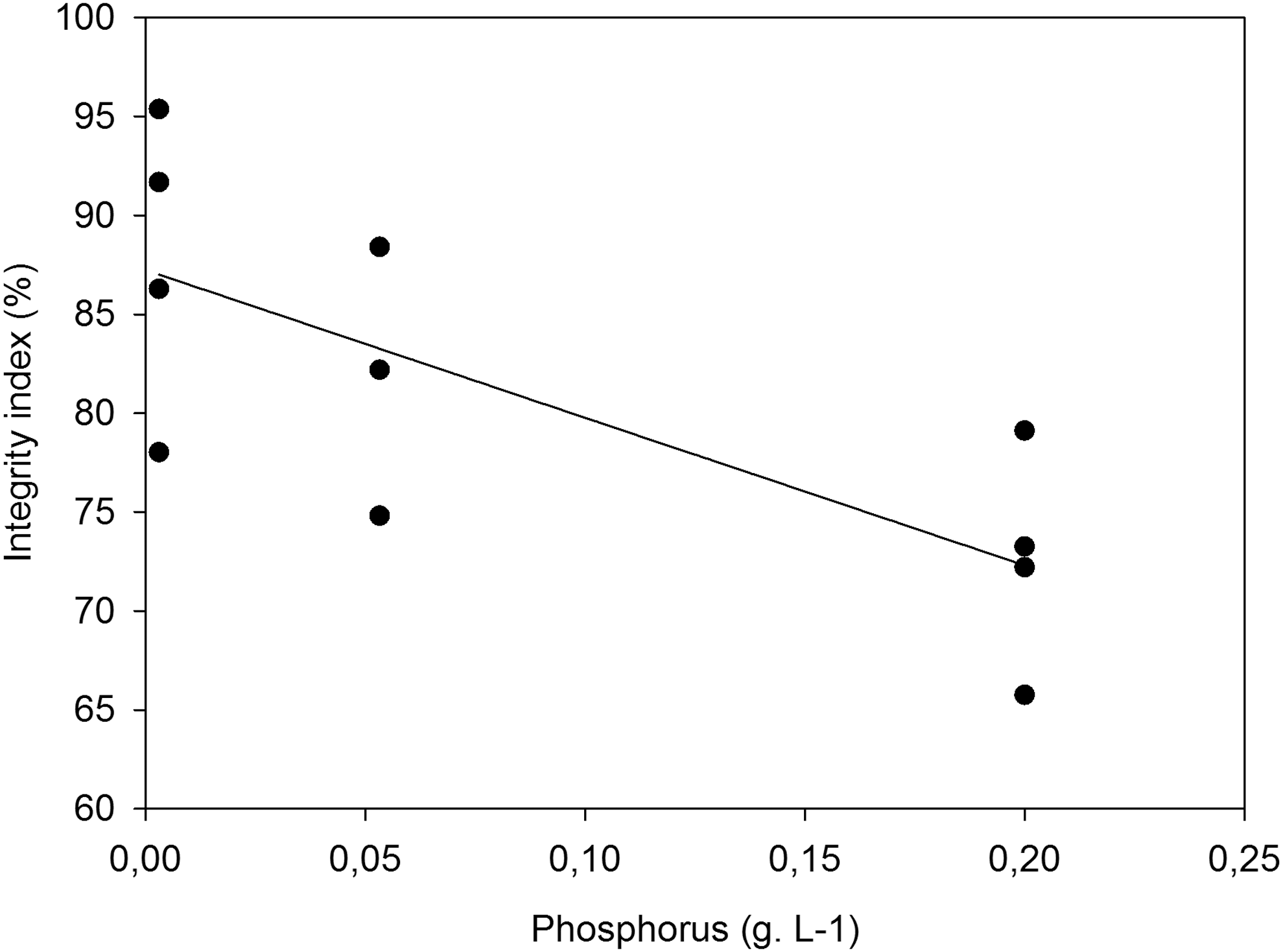
Linear regression of the model of consortia integrity index according to phosphorus concentration
Model of Consortium Integrity Index
Df: degree of freedom.
Correlations
The correlation matrix (Table 7) presents the relationship with others variables in addition to those included in the model (factors and responses). This analysis gives more information on relationships between responses and the indirect effects of nutrient concentration on the consortium, such as the variation in pH. Higher correlation coefficients are in bold. As expected, lipid accumulation in cells was negatively correlated to nitrogen concentration (−0.80), while biomass productivity was positively correlated (0.77) to nitrogen concentration. The consortium lipid and biomass productivity were highly correlated (0.94). Also, a positive correlation between initial pH and the integrity index was shown (0.71), as well as a negative correlation between final pH and neutral lipids in biomass (0.95).
Pearson Product Moment Correlation of Factors and Other Measured Variables
Organic carbon; bNitrogen; cPhosphorus; dLipids productivity; eBiomass productivity; fIntegrity index; gNeutral lipids accumulation in cells; hInitial pH; iFinal pH.
Fatty Acids Profile
The fatty acid composition of neutral lipids from extracted oil affects the quality of biodiesel produced from this oil. The fatty acid profile from each culture was analyzed to show the effect of different concentrations of organic carbon, nitrogen and phosphorus on the proportion of fatty acids, and consequently the biodiesel conversion potential of the oil produced during each treatment. The main fatty acids found were palmitic (C16:0), stearic (C18:0), and oleic (C18:1n9) acids (Fig. 5). Most fatty acids were saturated (57% to 91%), followed by mono-unsaturated (6% to 25%) and poly-unsaturated (<1% to 31%).
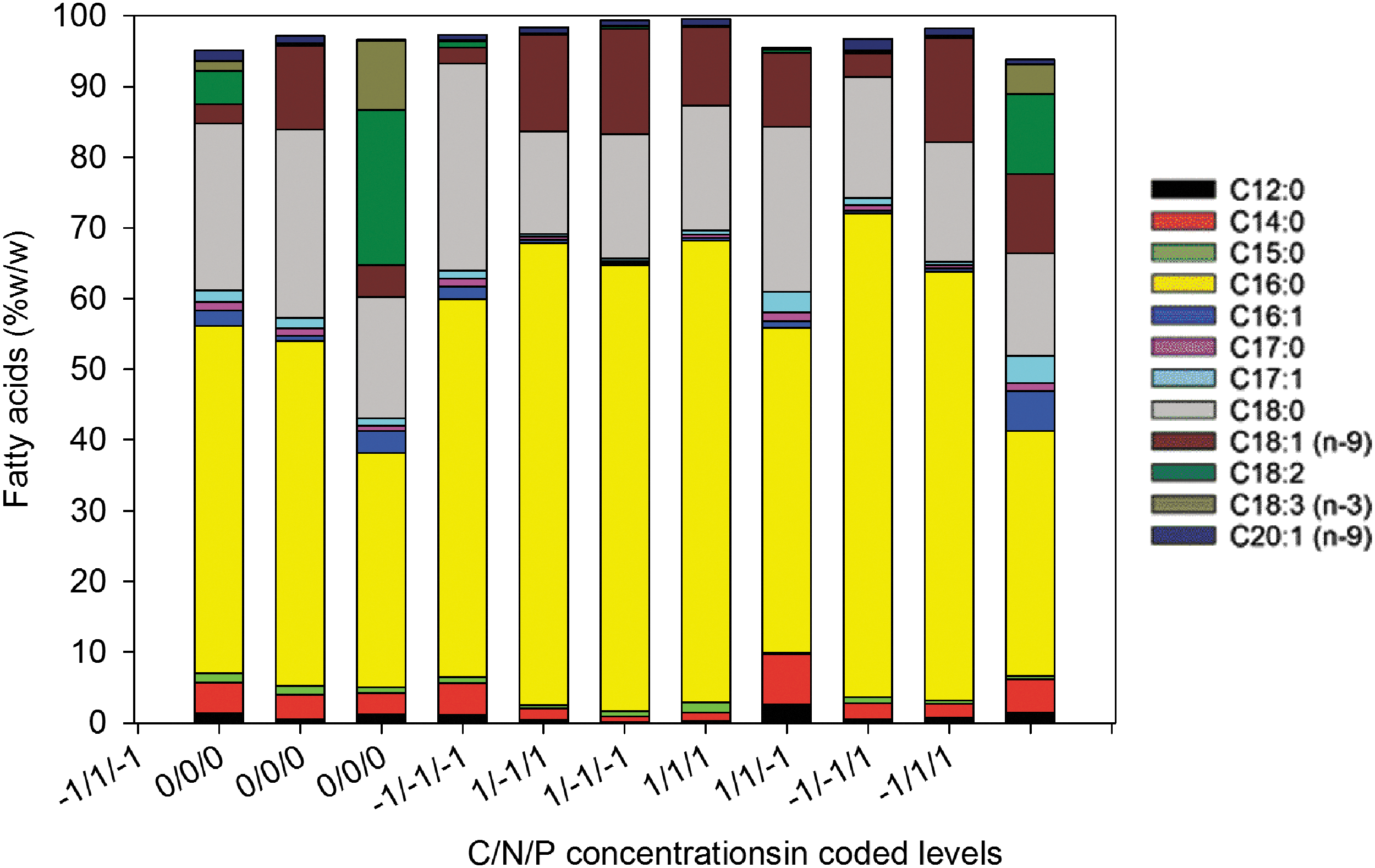
Fatty acids proportions (present in concentration more than 1% in more than one test) in the extracted lipids of design trials
A multiple linear regression analysis of the fatty acids whose concentrations exceeded 1% in more than 1 treatment was performed using 3 factors: C, N and P. Fatty acids in low concentration, except oleic acid, were the most affected (Fig. 5, Table 8). N was the more significant factor; a higher N concentration increased the proportions of lauric (C12 :0), myristic (C14:0), margaric (C17:0), and heptadecenoic (C17:1) acids in total fatty acids and decreased the proportion of oleic (C18:1(n-9)) acid. Interaction between C and N negatively affected the proportions of pentadecylic (C15:0) and heptadecanoic (C17:1) acids. Increasing P concentration decreased the proportion of pentadecylic (C15:0) acid and increased the proportion of heptadecenoic (C17:1) and oleic (C18:1(n-9)) acids. Maximal variation among the treatments was 12% for oleic (C18:1(n-9)) acids.
Models of Consortia Fatty Acids (Present in Concentration More than 1% in More than One Test) According to the C, N, P Factors
Df: degree of freedom.
Discussion
Lipids and biomass productivities
The productivity of the selected algae-bacteria consortium cultivated in aluminium smelter wastewaters was directly affected by the nutrient concentrations in the culture medium. Results from a 23 factorial design shows that among the concentrations used, the maximal concentration of N (0.200 g/L) and the minimal concentration of P (0.003g/L) resulted in the highest biomass (0.93 g/L/d) and lipid (0.023 g/L/d 1 ) productivity.
Very few studies have been performed on algal mixotrophic production in wastewaters other than agricultural food wastewaters. Bhatnagar et al. reported biomass productivity of 0.215 g/L/d using a Chlorella minutissima strain cultivated in enriched N (0.041 g/L N from NaNO3) and C (0.4 g/L C from glucose) wastewaters from a carpet factory. 21 Although lipid productivity was not reported in this study, another article from same authors reported a lipids productivity of 0.004 g/L/d under the same culture conditions, without enrichment in N and C. 29
As shown in Figures 1, 2 and 3, adding nutrients is essential to ensure consortium productivity in aluminium smelter wastewaters, but the impacts are different. While carbon is the major constituent of cells, its impact on biomass and lipid productivity was not significant (Table 4), despite the p-value for the parameter estimate of this factor (0.10; data not shown) being near to the threshold of 0.05. This result contrasts with the one obtained by Gélinas et al., who have demonstrated a 0.4 g/L gain in biomass, related to a mean glucose uptake of 1 g/L in a medium containing 0.1 g/L of N, with the same algae-bacteria consortium as that cultivated in the present study. 30 Also in the present study, maximal organic C concentration could have been too low, in proportion of the N and P inputs, to have a significant effect on productivity. Adding runs to experimental design, with higher C/N ratio, could provide higher productivity and should allow the detection of effect by the organic carbon concentration, as well as a quadratic relationship. For example, Liang et al. showed an inhibitory effect of high organic carbon concentration. 26 In a medium containing 0.04 g/L of N, the growth rate of a Chlorella vulgaris culture decreased from 0.250 g/L/d to 0.062 g/L/d with 10 g/L and 100 g/L of glucose, respectively.
Negative productivities, due to biomass loss, were seen in two treatments with low N concentrations (Table 2). Lipid productivity depends on two parameters: biomass productivity (favored by N) and lipid accumulation in cells (reduced by N). 31 An equilibrium must be determined; conditions for highest lipid accumulation per cell could reduce total biomass (and the number of cells), leading to reduced total lipid production. In our study, the algae-bacteria consortium reaches a higher lipid productivity in conditions with high nitrogen concentration (Fig. 2), although lipid accumulation in cells (proportion of neutral lipid cells) was negatively correlated to N concentration (Table 7). The correlation between lipid productivity and biomass was higher than the correlation between lipid productivity and lipid accumulation, thereby demonstrating that biomass productivity has a greater impact on lipid productivity than lipid accumulation. Li et al. came to the same conclusion using the strain Chlorella minutissima UTEX2341; 17 lipid productivity was higher when culture conditions were optimized for biomass production, although lipid accumulation per cell was not maximal. Moreover, N has a greater impact on lipid accumulation than organic C (Table 7), as previously noted in the literature; Heredia-Arroyo et al. observed that the organic C concentration had no effect on the proportion of lipids in strain Chlorella vulgaris 2714 cells. 32
In addition to nutrients, pH can have an effect on lipid and biomass productivity. The biomass of Chlorella protothecoides can be increased by 16% if cultivated at pH 6 rather than pH 8. 33 In the present study, pH and biomass production are not strongly correlated (Table 7). Lipid metabolism and accumulation are known to be impacted by pH. High pH stimulated triacylglycerol accumulation in Chlorella CHLOR1 strain cells. 34 However, in this study, the lipid accumulation in cells is inversely proportional to the final pH (R − 0.95), and there is no correlation with the initial pH (Table 7).
Integrity index
Pure cultures of microalgae in open ponds are known to be easily contaminated by predators and competitive microorganisms 35 unlike algal-bacterial consortia that are considered more stable. 36 Consortium ecology is affected by culture conditions, including some nutrient concentrations. 37 The determination of nutrient concentrations that maintain consortium integrity is essential for sustaining a high yield. In this study, consortium integrity is mainly affected by P concentration (Table 6, Fig. 3). The correlation results (Table 7) lead us to assume that this effect could be related to the negative impact of P on pH. Furthermore, consortium integrity has only a slight effect on biomass productivity as demonstrated by the weak correlation with these variables (Table 7). The integrity index was however significantly correlated with lipid productivity (Table 7). Some contaminants such as fungal hyphae could contribute to biomass but less to lipid productivity owing to the fact their lipid content is lower than that of algae.
Fatty acid profile
Biodiesel can be produced from microalgae oil. Its properties depend on the algae fatty acid composition. The degree of unsaturation and chain length of fatty acids are the two main factors to be considered in estimating the quality of a biodiesel made from microalgal oil.38 A large proportion of saturated and mono-unsaturated fatty acids and a small amount of polyunsaturated fatty acids are required to produce a biodiesel that meets European and American standards. Regarding chain lengths, the carbon number has to range from 8 to 18 C. In the present study, carbon chains lengths varied mainly from C12 to C18, and less than 1% were long chains (C20 and more) (Fig. 4). According to Ramos et al., the oil produced by the consortium could have an oxidative stability and combustion properties comparable to conventional diesel. 39
Nutrients such as N and P could support or reduce the production of certain fatty acids. C18:1 (n-9) showed the strongest response to N (estimate = -3.2482) and P (estimate = 2.9538) variations (Table 8). It has been previously noted that a lower N concentration enhanced the production of C18:1 (n-9), 38,40 but the P effect found in this study has not been documented in the literature. The fatty acids affected by N and P were in such a low concentration in cells that they would have no effect on the quality of biodiesel produced.
Conclusion
In this study, a native consortium of algae and bacteria, mainly Chlorella sp, was cultivated in wastewaters from an aluminium smelter using air CO2 and organic C as sources of C (mixotrophy). High N and low P quantities (in comparison with Bold's Basal medium) had to be added to these waters to enhance lipid and biomass productivity. It was shown that the culture conditions that enhanced biomass production resulted in higher lipid productivity than those that enhanced lipid accumulation in cells by reducing N. Consortium integrity was better preserved under these conditions (high N, low P), while high P promoted higher contamination of the consortium by unwanted microorganisms. Despite the variation in the proportion of fatty acids in relation to N and P, nutrient concentrations did not drastically effect algal oil quality and biodiesel production potential.
Footnotes
Acknowledgments
This work was funded by Rio Tinto Alcan, the Natural Sciences and Engineering Research Council of Canada (NSERC), the Québec Research Fund—Nature and Technologies, MITACS and the Consortium for Research and Innovation on Industrial Bioprocesses in Quebec (CRIBIQ). The authors wish to express thanks to all the members of the Industrial Université du Québec à Trois-Rivières-Foundation Research Chair on Environment & Biotechnology as well as to the personnel of the College Center for the Transfer of Technologies integrated to the Cegep of Trois-Rivières for their technical support.
Author Disclosure Statement
No competing financial interests exist.