
Abstract
This article reports an in-depth characterization and optimization of a fermentation process with the aim of increasing the production of high cell densities as a prerequisite for high productivity of lipids. The freshwater microalgae Chlorella protothecoides was grown heterotrophically in a bioreactor on aerated and stirred cultivation medium with glucose as a carbon source and yeast extract as a nitrogen source. The detailed effect of carbon to nitrogen (C/N) ratio on growth and lipid content of C. protothecoides was studied. Results show that with a C/N ratio lower than 12, the medium was carbon-limited, favoring cell proliferation, and under nitrogen-starvation conditions, (higher C/N ratios), lipid accumulation was enhanced. A two-stage, fed-batch process was developed and achieved a maximum of 255.3 g/L of biomass over 228 h of fermentation, from a low initial cell density (< 2 g/L). The specific growth rate, measured when C/N ratio was low, was 0.078 h−1. The lipid productivity was 16.7 g/L/day, and the maximum lipid content was 58 % (w/w) of the dry cell weight. This is the first time that such a very high cell density and lipid productivity have been achieved. The results offer potential for the production of important algae-derived commercial products.
Introduction
Although microalgae have been known for centuries, commercial, large-scale production only found applications in different fields of industry a few decades ago. To date, their potential in biotechnology is underexploited. The final cost of algal bioproducts depends mainly on the cultivation conditions and strain productivity. 1 High-cell-density cultivations are essential to increase the productivity of bioproducts. 2 Therefore, the ultimate objective for any bioprocess is to achieve the highest product concentration in the shortest time. 3 In this article, the development of an appropriate strategy to enhance biomass and target bioproduct by understanding the effect of different culture conditions and by altering the elemental composition of the cultivation medium has been studied.
The present study focuses on the heterotrophic cultivation of Chlorella protothecoides, a freshwater microalgal strain, to produce lipids. Microalgae do not accumulate lipids during “normal” growth, but overproduction can be induced in some strains by manipulating the physical and/or chemical conditions of the culture medium. 4,5 Nitrogen limitation is one of the most effective and commonly used mechanisms to stimulate lipid accumulation by microalgae, with up to more than 50% lipid accumulated by weight of dry biomass. 4 –8 Traditional methods to cultivate microalgae are open pond systems or closed photobioreactors. However, heterotrophic cultivation in conventional bioreactors is a feasible alternative. Organic substrates are used both as energy and as carbon sources. 4,10 There are several advantages to this method. Because growth is totally independent of light, high cell densities and higher productivities—at an improved growth rate—can be achieved. 11
In this study, a strategy to understand and optimize heterotrophic growth of C. protothecoides is presented. This involved the examination of the influence of carbon-to-nitrogen ratio in the medium and the determination of the optimal conditions to achieve high cell densities and enhance lipid accumulation. Finally, a two-stage fed-batch cultivation process was developed, with the two stages characterized by the C/N mass ratio of the cultivation medium. The aim of the first step was to achieve very high cell densities under carbon limitation, while the second step sought to enhance lipid accumulation under nitrogen limitation. It is important to specify that the term “nutrient-limited” does not imply a nutrient depletion in the medium, but refers to the nutrient that is depleted first (and whose depletion causes the cessation of growth). In batch mode, the initial limiting substrate concentration is proportional to the biomass reached at the stationary phase, and the biomass yield on substrate YX/S is constant.
Higher biomass and increased lipid productivity improve the potential of microalgae as sustainable sources of oil, biomass, food, feed, and other coproducts.
Materials and Methods
Materials
All chemicals were purchased from either Sigma Aldrich (St. Louis, MO) or Thermo Fisher Scientific (Waltham, MA) if not stated otherwise.
Microorganism and Medium
C. protothecoides (ATCC 30411) was obtained from the ATCC (Manassas, VA) culture collection. The components of standard cultivation medium were described by Chen et al. and are as follows (per L of distilled water): KH2PO4 0.7 g/L, K2HPO4 0.3 g/L, MgSO4·7 H2O 0.3 g/L, CaCl2 25 mg/L, NaCl 25 mg/L, FeSO4·7 H2O 3 mg/L, vitamin B1 0.01 mg/L, A5 Trace Solution 1 ml/L, glucose 30 g/L, yeast extract 4 g/L. 12 The pH of the medium was adjusted to 6.8 with 1 M NaOH followed by sterilization through a 0.22-μm filter (Steritop, Millipore, Billerica, MA).
Cultivation of the Microorganism
A cell bank of C. protothecoides was created in 5% (v/v) methanol and stored at −80 degrees. The study of the effect of carbon to nitrogen ratio (C/N) on cell growth was carried out in sterile, 500-mL baffled shake flasks with a working volume of 100 mL. The flasks were incubated in a shaking incubator (GFL, Burgwedel, Germany) with orbital motion of 120 rpm at 25°C.
The standard cultivation medium was prepared by varying the ratio between the initial concentration of glucose and yeast extract. The C/N ratio is calculated according to the formula:
It represents a mass ratio, where 40% is the composition of carbon in glucose and 11.4% that of nitrogen in yeast extract.
Cultures were scaled up to an autoclavable bench-top bioreactor (RALF Advanced, 2015, Bioengineering, Wald, Switzerland) adapted for microbial fermentations (5-L total volume); equipped with hollow baffles for temperature control, a Rushton agitator, a pH probe (Mettler Toledo, Columbus, OH), a temperature probe, and a dissolved oxygen probe (Mettler Toledo); and protected against light. pH was controlled with a conventional proportional–integral–derivative (PID) controller, which automatically added a flow of 1M NaOH or 0.5M H2SO4. A solution of 2% polypropylene glycol 2000 was used to control foam formation. Dissolved oxygen (DO) concentration was maintained ≥50% by increasing the air flow (from 0.75 vvm to 4 vvm) and the stirring rate (from 800 rpm to 1000 rpm). The temperature was kept constant at 25°C. The inoculum was made from a fresh pre-culture grown in a shake flask at exponential phase of growth. The initial working volume was 2,000 mL and the initial OD540nm in experiments carried out in Figs. 1–2 was 0.11 and 1.17, respectively. Dry cell weight (DCW) was less than 1 g/L and as a result could not be determined accurately by sampling small volumes. For the fed-batch mode of culture, sterile feed was composed of 700 g/L glucose and 2-fold concentrations of other medium components—excluding yeast extract—as described herein. Feed was manually added when glucose was completely depleted; the concentration of glucose was maintained under 80 g/L to avoid growth inhibition.

Fed-batch cultivation of C. protothecoides. Results are represented as ♦ DCW in g/L; ■ optical density (OD540nm); and ○ glucose concentration in g/L as a function of the cultivation time in h. The DCW and the OD540nm were measured in duplicate (n = 2), the standard deviation is represented as error bars.

Improved fed-batch cultivation of C. protothecoides. Results are represented as ♦ DCW in g/L; ■ optical density (OD540nm); and ○ glucose concentration in g/L as a function of the cultivation time in h. Total lipids accumulation (▴) is represented as percentage of DCW. The DCW and the OD540nm were measured in duplicate (n = 2) and the total lipid percentage of the DCW in sextuplicate (n = 6). The standard deviation is represented as error bars.
Growth Analysis and Lipid Extraction
Optical density (OD) measurements at 540 nm with UV/VIS spectrophotometer (UniSPEC2, LLG Labware, Meckenheim, Germany) were used to monitor algal growth. Samples were diluted in deionized water to measure absorbance only between the linearity range of 0.1–0.3. Results were calculated as follows:
To determine the DCW, 3 mL of the whole culture broth sample was transferred into a dried, preweighed glass tube and centrifuged at 1,000 rpm for 10 min with a floor centrifuge (FL40R, Thermo Fisher Scientific). The cell pellet was resuspended in 10 mL deionized water, centrifuged, and the supernatant discarded to wash the biomass from medium residues. Glass tubes were dried at 100°C for at least 24 h. The tubes were cooled in a dessicator and weighed. DCW was estimated as follows:
Glucose concentration was analyzed photometrically using a Cedex Bio Analyzer (Roche, Basel, Switzerland). Nitrogen concentration in the medium was not measured in these experiments.
C. protothecoides cells were pretreated previous to lipid extraction (to disrupt the cell wall) by continuously vortexing a cell suspension with ∅ 1-mm glass beads for 12 min. In a glass test tube, 1 mL of treated cell suspension was regularly vortex-mixed with 2 mL methanol and 4 mL dichloromethane. After 2 h, 1 mL methanol and 2 mL dichloromethane were added and centrifuged. The aqueous upper layer was removed, and the organic layer was filtered through a Whatman (Maidstone, United Kingdom) No. 1 filter paper in a Buechner funnel in a preweighed test tube. Solvents were evaporated in a water bath, and remaining lipids were flushed with nitrogen. The content of total lipids was determined gravimetrically:
In the case of OD540nm and DCW, measurements were repeated twice (n = 2) to check the reproducibility of the results. Lipid extraction was repeated 5 times (n = 5). Experimental results are reported as mean value ± SD and error bars are reported in the figures.
Results and Discussion
Effect of Different C/N Ratios on Algal Growth
Microalgal cells were initially grown in the standard cultivation medium described by Chen et al. 12 This was suitable for the cultivation of C. protothecoides and provided a good base to start the medium optimization. The threshold between carbon-limitation and nitrogen-limitation was defined by cultivating cells under different C/N mass ratios and exploring the effect on biomass yield. It was expected that when the carbon substrate was limiting, the initial concentration would determine the final biomass. Standard cultivation medium was modified by keeping the yeast extract concentration constant at 10 g/L and increasing the concentration of glucose (5; 15; 30; 31.35; 37.05; 42.75; 48.45; 57.15; 60; 90 g/L) according to the formula indicated above.
Figure 3 represents the maximum biomass obtained by cultivating Chlorella in the growth medium as a function of different initial concentrations of glucose. Up to a C/N ratio of 11, the relation between the amount of DCW formed and the initial concentration of glucose was linear (R 2 = 0.99). Analysis confirmed that glucose was completely consumed at the beginning of the stationary phase and thus the culture was glucose limited (results not shown). C/N mass ratio of 11 corresponded to 31.35 g/L glucose and 10 g/L yeast extract, and glucose consumption with C/N ratios higher than 11 was always between 29 g/L and 31 g/L. Furthermore, with ratios higher than 11, DCW remains unchanged, reaching a plateau. The hypothesis is that this was due to the switch to nitrogen limitation. However, it may have been the case that the initial concentration of glucose (higher than 31 g/L) was having an inhibitory effect on growth. Further experiments were needed to clarify the matter. Therefore, in the second part of the work, the culture experiments were repeated, keeping the glucose concentration constant at 20 g/L and varying the yeast extract (1; 2; 4; 6; 8 g/L) (Fig. 4).

Maximum biomass as a function of the initial glucose concentration. Labels indicate the respective C/N ratio. The DCW corresponds to an average of two points (n = 2) measured during the stationary phase, the standard deviation is represented as error bars.
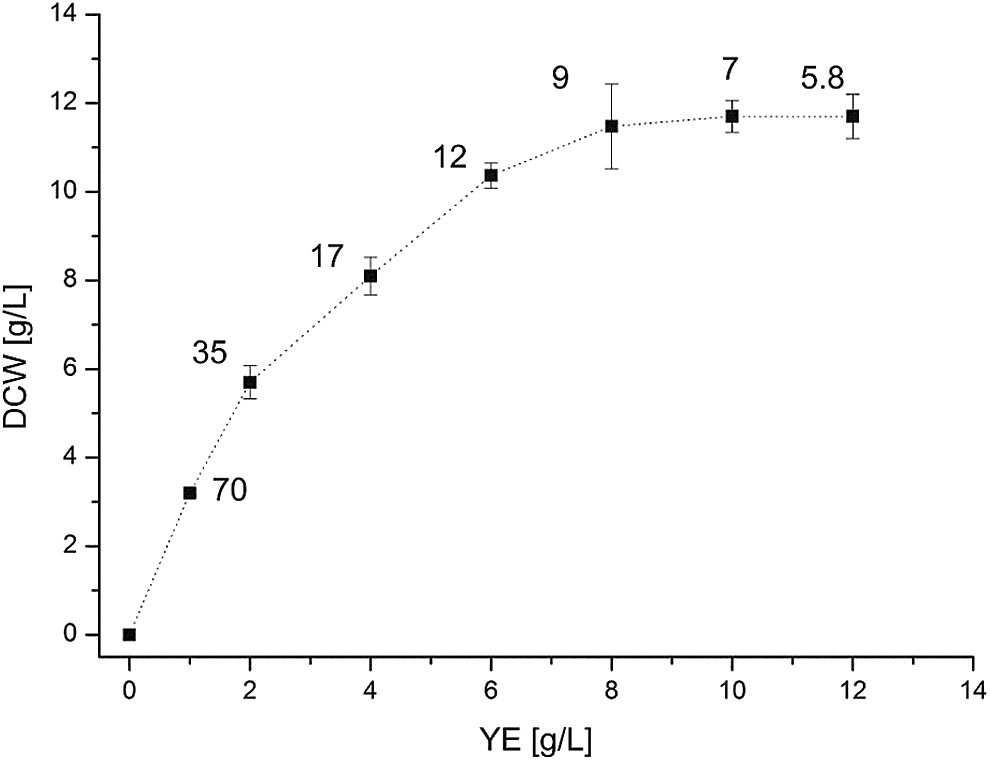
Maximum biomass as a function of the initial yeast extract concentration. Labels indicate the respective C/N ratio. The DCW corresponds to an average of two points (n = 2) measured during the stationary phase, the standard deviation is represented as error bars.
Results in Fig. 4 show that, with a C/N ratio lower than 35, biomass increased proportionally with the amount of yeast extract in the medium. Subsequently, the curve increased slowly until forming a plateau at a C/N ratio between 9–11. In other words, with a lower concentration of nitrogen (higher C/N ratio) the medium became limited in nitrogen and not in glucose. The biomass to yeast extract yield (YX/YE) is 3 g/g and corresponds to the slope of the linear part of the graphic (Fig. 4). The plateau was reached because of the new substrate limitation and not because of the inhibitory effect of the glucose. The conclusion was that the threshold between carbon limitation and nitrogen limitation appears to be at some point between a C/N ratio of 9–11. It was difficult to set a precise limit value because of the slightly heterogeneous nitrogen concentration in yeast extract. According to the literature, for the microalgal strain Chlorella sorokiniana, it was found that the switch from carbon to nitrogen limitation occurred at a C/N ratio of approximately 20. The cell lipid content was at a minimum below this value and increased at higher C/N values. 13 Singhasuwan and collaborators report that the biomass production of Chlorella sp. TISTR 8990 was maximized at a C/N ratio of 29, and with a ratio higher than 63 the accumulation of total fatty acids was enhanced. In order to obtain the target C/N ratio, they used glucose and potassium nitrate. 7 Cheng et al. showed that the maximum oil content was obtained when C/N was 19.8. 14
Batch Cultivations
With a carbon-to-nitrogen ratio lower than 12, medium was carbon limited. The next step was to demonstrate the proportional increase of the biomass as a function of the initial glucose concentration, typical of a carbon limitation. Main objectives were to prove that the biomass yield on glucose is constant; to define the optimal concentration of initial glucose where the biomass is maximal, avoiding any inhibitory effect or metabolic overflow; and to define what the highest biomass attainable in batch culture in bioreactor is.
The slope of the linear part of the graph (R 2 = 0.999) represented in Fig. 5 corresponds to the biomass yield on glucose (YX/S) with a value of 0.66 g/g. Chlorella was grown under glucose limitation (constant C/N 8.77), and the initial substrate concentration was completely consumed, resulting in a proportional increase of the biomass reached at the stationary phase. With 120 g/L of initial glucose, the yield decreased slightly, to 0.57 g/g, and during the fermentation, lactate was produced up to a concentration of 1.7 g/L (results not shown). These values suggested the beginning of a metabolic overflow, and it was suspected that this high concentration was already beyond the optimal point. With higher concentrations of glucose, 167 g/L, cell growth was very low, 0.003 h−1 . Concentrations higher than 80 g/L of glucose have a negative effect on growth lowering the growth rate close to 0 h−1 . These results are in agreement with scientific literature; Xiong et al. defined that glucose concentration higher than 60 g/L had an inhibition on cell growth and, in the study of Doucha and Livansky, it was claimed that Chlorella has good tolerance to glucose—up to 80 g/L obtaining a cell yield on glucose of 0.55–0.69 g/g. 1,15

Maximal DCW of C. protothecoides achieved in batch bioreactor as a function of the initial concentration of glucose. The C/N mass ratio was constant at 8.77. The DCW max corresponds to an average of three points (n = 3) measured during stationary phase, the standard deviation is represented as error bars.
The present study shows that when the initial concentration was 20 g/L, the maximum specific growth rate was 0.11 h−1 . With the increase of initial substrate concentration, the growth rate decreased, which may be a result of substrate inhibition. Between the growth rate and substrate concentration exists an hyperbolic relationship, described extensively by Monod. The growth rate rises as a function of the initial substrate up to a maximum value (μmax), however, beyond the maximum value, in case of substrate inhibition, the growth rate decreases. 16,17 Such results are in agreement with previous studies. The maximum growth rate of Chlorella sp., determined by O'Grady et al., was 0.096 h−1 , and the estimated mean specific growth rate for Doucha and Livansky was 0.11 h−1 . 1,18
The highest biomass in batch culture was obtained growing cells in a medium where glucose was the limiting substrate (C/N 8.77); from 120 g/L of initial glucose, 69.83 g/L (SD = 3.535 · 10−3 g/L, n = 2) of DCW was produced. Optimization of the biomass production is an important prerequisite to increase the productivity of lipids. According to the literature, higher cell densities, up to 117 g/L, were achieved with other Chlorella species, i.e., Chlorella vulgaris, improving the cultivation to fed-batch mode. 1 Most biotechnological production platforms of growth-associated products are based on fed-batch cultures because of advantages such as the achievement of high cell densities, the control over the production of byproducts, and improved productivity. 19 Fed-batch operation may be the solution to overcome the contradictory mechanism of cell proliferation and lipid accumulation by combining two different strategies. First, a high cell density may be achieved using a carbon-limited medium, and then, by changing the medium feed addition, lipid accumulation may be enhanced under nitrogen limitation.
Two Stage Fed-Batch Cultivation
Heterotrophic bioreactor cultures of C. protothecoides were optimized to fed-batch operations with the aim of achieving high cell densities under a carbon-limitation medium and enhancing the lipid accumulation under nitrogen limitation. The objective was to improve the overall lipid productivity. Microalgae showed a very high tolerance to glucose, thus a pulsed feed strategy was chosen instead of a complicated regulated system. This strategy consists of adding nutrients to the bioreactor based on the output signal of an indirect measurement of the substrate concentration, in this case glucose. Over-feeding might lead to negative effects on cell growth, therefore the appropriate feeding strategy considered the glucose inhibitory level. The maximum concentration of glucose in the medium after feed addition was kept under 80 g/L (Fig. 5).
A fed-batch fermentation with C. protothecoides was carried out with the cells initially cultivated in batch mode in medium with C/N ratio of 8.77 (Fig. 1). When glucose was completely depleted, feed composed of 700 g/L glucose and concentrated salts was added. A source of nitrogen was contained only in the initial cultivation medium and not in the feed, therefore it was expected that the continuous addition of glucose would increase the C/N ratio until complete nitrogen limitation was achieved—which is expected to gradually enhance the lipid accumulation.
Yeast extract concentration was 32 g/L, therefore approximately 100 g/L of biomass was expected (YX/YE 3 g/g). According to Fig. 1, such a value corresponds to the point where the optical density measurements achieve a plateau, OD540nm 500 [-]. After 170–190 h, the biomass was 100 g/L, which continued to increase up to about 150 g/L at the end of the culture. The hypothesis was that the difference of 50 g/L does not correspond to an increase in cell number but to an apparent lipid accumulation. The theoretical percentage of total lipid in the biomass corresponds to about 50% DCW. It was suspected that after 170–190 hours, cells stopped proliferating because of nitrogen-limitation. Consequently, under such stressed conditions, lipids were accumulated. Three growth stages can be distinguished based on the growth rate, which was calculated from the DCW data: (1) batch phase 0.069 h−1 (until 129 h); (2) fed-batch 0.01 h−1 (between 130–190 h); (3) lipid accumulation 0.005 h−1 (from 196–260 h). The growth rate during the hypothetical lipid accumulation phase was very low, corresponding to a cell doubling every 140 h. This indicates that in the last stage of cultivation, the biosynthesis of lipids was the major cellular activity, and cell division ceased. Total lipid percentage of biomass was determined only when cells were harvested (after 313 h), and corresponded to 55.18% (SD = 3.275%, n = 5), confirming that lipids were accumulated.
Improved Two-Stage Fed-Batch Fermentation and Lipid Analysis
The aim of the next experiment was to achieve higher cell densities of C. protothecoides by feeding more yeast extract (source of N) after the end of the batch phase. The expectation was that by adding more nitrogen, the carbon-limited phase would be prolonged, providing good conditions for cell proliferation and, consequently, higher cell densities. Based on the same suppositions applied for the previous culture, it was expected that by adding 15 g/L of yeast extract during the feed, 156 g/L of biomass would be produced.
Figure 2 shows that after 134 h, 168 g/L of biomass were achieved. At the same point, optical density measurements plateaued around OD540nm 800 [-]. Hypothetically, cells entered the stationary phase due to the complete consumption of nitrogen. The difference in biomass yield between 134 h and 255 h of fermentation—87 g/L—likely corresponds to total lipid accumulation, corresponding to approximately 52% of total lipids in the biomass. Total lipid extraction was carried out at different intervals of the cultivation. The aim was to define the total lipid percentage of the biomass over time.
Theoretically, cell growth and lipid accumulation can be divided into two stages based on the hypothetical C/N ratio. Under optimal growth conditions, algal cells have a relatively low lipid content, which constitutes about 5–20% of dry cell weight. 5 A similar result is expected when the C/N ratio is lower than 12, analogous to a carbon limitation. Cells will enter into a transition phase were nitrogen is present, but not in excess and continuously decrease. During the so-called transition phase, the C/N ratio is equal or slightly higher than 12. It is expected that under these conditions, cells accumulate lipids gradually, up to a point where nitrogen is not present anymore (C/N >>> 12) and the maximum lipid level is attained.
Figure 2 shows that after about 60 h—at the end of the batch phase—biomass was composed of 9.982% (SD = 0.5046%, n = 5) of total lipids. This value was similar to that defined by the literature, 5 and it was assumed that even at the beginning of the culture the composition was similar. In fact, initial cultivation medium had a C/N of 8.77. Lipid percentage of biomass appeared to increase steadily until 140 h. When the lipid content was equal to 33.75% (SD = 0.9332%, n = 5), the biomass corresponded to 108 g/L, meaning that hypothetically only 15 g/L of nitrogen were still available in the medium. Therefore, the theoretical C/N was about 20, a value situated in the transition phase. Only after 140 h, the percentage of total lipids in the biomass reached a maximum of 58.10% (SD = 2.487%, n = 5). This high percentage of lipids is strongly indicative that nitrogen was completely depleted.
Analysis of results reported in Figs. 1–2 revealed a linear relation between DCW values as a function of the OD540nm measurements, until the beginning of the stationary phase (result not shown). Under low C/N ratios, cell proliferation was enhanced and cell weight increased proportionally. After the achievement of stationary phase, where C/N ratio in the medium was higher, the DCW increased because of an accumulation of lipids and not because of an increase in cell number, therefore the correlation between optical density and DCW is no longer constant.
The results of this experiment showed that under specific culture conditions—high O2 mass transfer due to high agitation rate, leading to dissolved oxygen concentrations higher than 50%; specific C/N ratio in the medium; feeding more nitrogen after the batch phase)—it was possible to achieve higher biomass, 255.3 g/L (SD = 2.828 · 10 −3 g/L, n = 2), and consequently accumulate more lipids. In term of lipid productivity, the maximum was 16.7 g/L/day.
Discussion
The present study has developed a fermentation strategy to obtain very high cell density cultivation of C. protothecoides, 255.3 g/L, from an initial concentration of less than 2 g/L and with a high productivity of lipids (16.7 g/L/day). The specific growth rate—0.078 h−1 —was calculated under carbon-limited condition in the batch phase (low C/N ratio).
Such results are considerably higher than reported in other studies for high cell density heterotrophic algal cultivation. Table 1 illustrates different examples. Chen et al. reached 45.2 g/L of biomass after 197 h in fed-batch mode of culture, and the maximum lipid content was 54% of DCW. The basal medium was prepared with 30 g/L glycerol as carbon substrate and 4 g/L yeast extract, and the feed solution contained 150 g/L glycerol and 15 g/L yeast extract/L to maintain a certain C/N ratio. 12
Overview of Fed-Batch Strategies for Heterotrophic High Cell Density Culture of Chlorella sp.
Data was estimated from graph observation; bMedium was supplemented also with another C-source, sodium acetate
Xiong et al. achieved cell density of 51.2 g/L in 167 h in a 5-L bioreactor by performing an improved fed-batch culture strategy. However, lipid content was 57.8% in batch culture and only 50.3% in fed-batch culture in a 5-L bioreactor. 15 Zheng et al. developed a two-stage fed-batch fermentation, resulting in 103.8 g/L biomass. In the first stage, the C/N ratio was 29, and during the second stage, only carbon was supplemented (similar to what done in the present study). The lipid content achieved a maximum of 38.7% after 228 h (Table 1). 20
A major limitation of heterotrophic algal cultivation is the sustainability and cost of organic carbon source. Sugar cane juice hydrolysate (SCH) was used by Cheng et al. as an alternative carbon supply for lipid production. In fed-batch cultivation, Cheng et al. reported 48.52 g/L biomass with 54.7% of lipids after 170 h. 14 In another experiment, instead of enhancing the lipid accumulation by exposing cells to nutrient starved conditions, Palabhanvi et al. supplemented the medium with a lipid inducer. 21 According to this study, sodium acetate, being a simply structured molecule, seemed to be better metabolized by cells into acetyl coA, the main metabolic precursor for lipid biosynthesis. As a result, 90.15 g/L biomass was achieved with a lipid productivity of 7.7 g/L/day. Interestingly, the biomass was composed of 70.2% w/w lipids (Table 1). 21
Other studies were focused mainly on producing high yield of biomass and not lipids. Wu et al. obtained a cell density of 116.2 g/L, 22 whereas Doucha and Livansky achieved 117.18 g/L in only 40 hours (Table 1). 1
In all the presented studies, starting cell densities were always lower than 10 g/L making it possible to compare lipid productivities.
Conclusion
In the present study, the effect of C/N ratio on cell growth was explored and enabled the development of a two-stage, fed-batch fermentation process. The high levels of biomass obtained compared with previous studies (Table 1) was due to initially maintaining a low C/N, which provides very good conditions for cell proliferation, followed by a high C/N ratio to limit cell growth and promote lipid accumulation (58% w/w). As a result, a very high lipid productivity of 16.7 g/L/day was achieved. In this study of algal biomass (DCW), over 255 g/L was attained, which is 2–6 fold higher than previously reported. Furthermore, lipid productivity was 2–5 fold higher than previously published data. While the process allowed for only one run, it illustrates the potential for very high outputs. Next steps would be to confirm such improved yields in scale-up experiments. The process may be further optimized by increasing the inoculum concentration, which will reduce fermentation time. This work showed also the importance of an organic nitrogen source, yeast extract. However, it would be beneficial to find cheaper organic sources for commercial development of the work.
The higher biomass and increased lipid productivity obtained in this work improve the potential of algal cultivation, which should find more applications with real-world benefits for uses as food ingredients, aquaculture and animal feed, bioenergy, and products for human health, nutrition and personal care.
Footnotes
Acknowledgments
The authors acknowledge funding from Irish Research Council (IRC) as Employment Based Program in collaboration with Monaghan Bioscience, Ireland
Author Disclosure Statement
No competing financial interests exist.