
Abstract
For decades, conventional plastics obtained from fossil-based sources have been used indiscriminately due to their durability and resistance. However, their use is problematic because of their rapid disposability and slow degradation. Growing scientific interest in the environmental issues associated with rising plastics consumption has become of increasing importance in the search for biodegradable substitutes. Poly (3-hydroxybutyrate) (P(3HB)) is one of the most studied and characterized microbial biopolymers from the family of polyhydroxyalkanoates (PHAs). Its main features are rapid biodegradability, low toxicity, and biocompatibility. Global production of PHAs is approximately 100 tons/year, and is expected to grow to 500,000 tons/year by 2020. Researchers have sought to increase the intracellular accumulation and polymer yield of PHAs by screening strains and improving the cultivation and operating procedures employed during the production processes. Synthesis of P(3HB) normally involves two stages. The first step occurs under growing conditions without nutrient limitation. The second step, polymer production, occurs under conditions in which some of the essential nutrients, such as P, Fe, Mg, and C, are limited and carbon is excessive. This review presents an overview of the metabolic mechanisms of synthesis, production process, recovery and applicability of P(3HB)-related microorganisms as potential producers. It also highlights studies on the optimization of carbon and nitrogen sources in the culture medium and operational parameters that optimize the bioprocess used to obtain P(3HB), both at laboratory- and industrial-scale.
Introduction
The economic exploitation of polymers of biological origin—or biopolymers, as they are also known—has occurred in parallel with human development. For many years, there has been considerable interest in biopolymer polysaccharides produced by microorganisms. These polysaccharides are known as microbial biopolymers and have the ability to form viscous solutions and gels in aqueous media. 1 Recently, however, other types of microbial biopolymers have attracted attention due to their thermoplasticity characteristics and solubility in association to biodegradability: polyhydroxyalkanoates (PHAs), including poly(3-hydroxybutyrate) (P(3HB)) and its copolymers. 2
This review describes the metabolic mechanisms of synthesis, production process, recovery, and applicability of P(3HB) and the potential microorganisms that can produce P(3HB), both at laboratory- and industrial-scale. It also emphasizes the various studies involving carbon and nitrogen sources in the culture medium and the operational parameters by which a higher yield of biopolymer can be obtained.
Biodegradable Plastic Polymers
Three types of biodegradation are of interest when examining biodegradable plastics: completely biodegradable, photo-biodegradable, and semi-biodegradable. The first type includes those plastics that have an intrinsic biodegradation characteristic. For example, the composition of photo-biodegradable polymers includes additives that incorporate polymer chain groups that are sensitive to light. The degradation of these plastics is initiated when they are exposed to ultraviolet radiation and, as a consequence, complete biodegradation may occur. Semi-biodegradable blends incorporate biodegradable polymers, such as starch, and additional polymers that are non-biodegradable. Thus, microorganisms can attack the starch and instigate biodegradation; however, the non-starch portion of the polymer remains non-biodegradable. 3
Relative to the origin, fully biodegradable polymers are classified into four groups: i) agro polymers, which are generally obtained by fractionation of biomass from agro products and are divided into polysaccharides, proteins, and lipids; ii) microbial biopolymers, which are synthesized by the microorganisms from renewable sources, as is the case with PHAs; iii) chemically semi-synthesized polymers, which are derived from monomers of the biotechnological processes, such as poly (lactic acid) (PLA), which can be obtained by polymerizing the lactic acid produced by microbial fermentation; and iv) polymers from non-renewable sources (fossil fuels), which are synthesized by chemical processes such as aromatic co-polyesters (PBAT). 4
The breakdown of biodegradable materials in the environment depends on factors such as how the polymer is discarded; the presence of microspecific enzyme generators that facilitate decomposition; temperature: pH: the oxygen content in the medium; and nutrients. 5 Further, the intrinsic factors of the polymer, such as the degree of crystallinity and molar mass, may influence the rate of biodegradation. 6
PHAs
The discovery of bright, lipophilic granules in Beijeirinckia cells was reported for the first time in 1888. However, it was not until 1923 that Maurice Lemoigne utilized Bacillus megaterium and identified the first compound—P(3HB)—as a probable storage material. Over the next thirty years, interest in P(3HB) was restricted to the description of methods of detection and the determination of the bioplastic content of cells and culture conditions that would lead to their synthesis and degradation by the Bacillus. In 1958, Wilkinson Macre found that the storage granules of B. megaterium increased when the medium proportions of glucose and nitrogen were increased, and he concluded that these compounds were likely a source of carbon and energy for the microorganism studied. 7
In Brazil, the technology associated with the production of PHAs, biodegradable, and biocompatible plastics, was first introduced in the 1990s at the Institute for Research and Technology (IPT), Copersucar (São Paulo, Brazil), and the University of São Paulo, where researchers used carbohydrates as the starting material and sought strategies by which it was possible to use the microorganism derived from Brazilian soil.
The low cost of sugar production in Brazil, combined with the energy surplus of sugar and alcohol, makes Brazil an ideal location for the integrated production of these polymers. Production was inaugurated in September 2000 by PHB Industrial (São Paulo, Brazil), a company that produced and marketed biodegradable plastic type P(3HB) under the name of Biocycle®. 8 The industry operated on a pilot scale until 2015 and even exported products to Japan. However, industrial plant is now defunct.
PHAs represent a class of polyesters that uses many microorganisms as a natural source. These microorganisms accumulate in the form of granules that have an intracellular carbon reserve function for energy. 9 The accumulation of these granules can exceed 80%. 10 These polymers have attracted significant attention from researchers because they are thermoplastic, biodegradable, and elastomeric, and can be produced from renewable sources. 10 –18
An important feature of PHA is that it is biodegradable. In nature, many microorganisms can degrade PHA through hydrolase enzymes and the PHA depolymerase produced within the cell so PHA can be used as a carbon source. However, these enzymes cannot degrade the polymer externally; i.e., after it is removed from the cell, and this is potentially due to the change from the crystalline state to the amorphous state.
Many of the microorganisms found in soils, marine waters, and lakes have an extracellular PHA depolymerase that can degrade the polymer after removal from the cell so it can be used as a carbon source. The degradation ability of PHA is not restricted to bacteria; many fungi can also degrade these biopolymers. 19 -21 The activity of these enzymes may vary and depend on the polymer composition, physical form of the sample dimensions, and the environmental conditions. 9
The rate of the biodegradation of PHAs is influenced by several factors, some of which are related to the environment, such as time, temperature, humidity, pH, and nutrient availability, and others that are related to the biopolymers themselves (composition, crystallinity, additives, thickness, and surface area). Biodegradation starts on the surface through enzymatic hydrolysis. 21,22 Enzymatic degradation involves two steps: first, the adsorption of the enzyme on the polymer surface and second, the hydrolysis of the polymer chains, which occurs at the active site of enzymes. 21,23
Under aerobic conditions, degradation results in the production of carbon dioxide (CO2) and water (H2O). However, under anaerobic conditions, degradation occurs at the end of the process, resulting in the production of CO2 and methane (CH4). 3 Activated sludge and organic soil are among the proposed inoculants to degrade this biopolymer.
P(3HB)
P(3HB) is the PHA that has been the most frequently studied and characterized to date. P(3HB) is a natural polymer and biodegradable aliphatic homopolymer type that is composed of monomers of four carbon atoms. 24 It is soluble in some organic solvents and insoluble in water. 18 It has the chemical formula (C4H6O2)n, which corresponds to 55.81% carbon, 7.03% hydrogen, and 37.16% oxygen by weight. 25
The industrial interest in the use of P(3HB) emerged during the 1960s when its thermoplastic properties were first described. 26 Its synthesis was first conducted in 1971 via the polymerization of a racemic mixture of β-butyrolactone using a catalyst system of triethylaluminum and water, which resulted in a stereoregular polymer that is partially optically inactive. In additional studies, P(3HB) was biotechnologically obtained from bacteria with a low molar mass (1x104 Da) and crystallinity (29%). 25,27
Because of its interesting thermoplastic properties, in its original form, P(3HB) resists a wide temperature range (from 30°C to 120°C) and can be processed as a classic thermoplastic. 28 P(3HB) is non-toxic, and its degradation produces 3-hydroxybutyric acid, a normal constituent of human blood even at high concentrations. Thus, it can be used both in products that come into contact with human or animal tissue and for human consumption. 29
At present, several companies are dedicated to the production of PHB and its copolymers, both at pilot and industrial scale. The largest number of these companies are located in China. The Tian company produces P(3HB-co-3HV), trade name Enmat®. It had an estimated capacity of 2,000 tons/year in 2007, but this is expected to rise to 50,000 tons/year by 2020. Green Bio-DSM produces KanekaPHBH® [P(3HB-co-3HHx)]. Its capacities were not disclosed in 2007, nor was its outlook for the year 2020. The Tianzhu company produces PHBH®, i.e., the polymer form of P(3HB-co-3HHx), at both pilot and research scales.
Danimer Scientific (Bainbridge, GA) produces Meredian PHA®, which is a non-specific polymer. Production of this polymer is expected to reach 272,000 t/year by 2020. Additionally, in the United States, the TephaFlex® TephElast® polymers are produced at both pilot and research scales. In Japan, Mitsubishi (Tokyo) produces P(3HB) under the trade name Biogreen® at both the pilot and research scales. It did not disclose its production capacity in 2007 or its expected future capacity for 2020. Kaneka (Osaka, Japan) produces P(3HB-co-3HHx), marketed as Kaneka PHBH®, with a capacity in 2007 of 100 tons/year; the forecast for 2020 is 50,000 tons/year. In Ulm, Germany, Biomers produced 10 tons/year in 2007 under the name Biomer™ at both the pilot and research scales. In Canada, the Biomatera (Toronto) company produced the polymer P(3HB-co-3HV), marketed under the name Biomatera®, at both the research and pilot scales. The Bi-on Company (Bologna, Italy) produces PHA-MINERV, which is completely degraded within 10 days in the normal river water, and is considered an alternative to polyethylene terephthalate, polyethylene, polypropylene, high-density polyethylene, and low-density polyethylene. BASF (Ludwigshafen, Germany) produces P(3HB) and poly-(3-hydroxybutyrate-co-3-hydroxyvalerate) (P(3HBV)) biopolymers at pilot scale. The same polymers are produced in the United States by Monsanto (St. Louis, Missouri). In China, TianAn Biopolymer produced P(3HBV) with the commercial name ENMAT® at industrial scale (2,000 tons/year), and until 2007, Tianjin GreenBio produced 1,000 tons/year of the copolymer P(3HB-co-4HB).
While there has been expansion, many industries have ceased production of PHAs. Zenica, located in England, produced Biopol® polymer P(3HB-co-3HV), but suspended production in 1996; Monsanto, which produced the Biopol® brand (P(3HB)/PHV/PHA) in Italy, ceased production in 1998. Procter & Gamble (Cincinnati, OH) produced Nodax® P copolymer (3HB)/HHX, but ended this activity in 2006. However, it passed its production technology to Kaneca and Meredian, now Danimer. The technology Biopol®, which belonged to Zeneka, was transferred initially to Monsanto, which, in turn, transferred it to Metabolix (Cambridge, MA). In 2007, Metabolix and Archer Daniels Midland company (ADM; Chicago, IL) formed the Telles Company, which was dissolved in 2012. The Imperial Chemical Industries (London, England) produced the copolymer P(3HBV) until 2008.
Until recently, P(3HB) was produced in Brazil by the PHB SA company under the name BIOCYCLE®. The company, located in Serrana, São Paulo, operated in partnership with the Institute of Technological Research, Copersucar, and the University of São Paulo. They used the microorganism Ralstonia eutropha, which was genetically modified for the use of sucrose as a carbon source, in a pilot plant. The company projected a capacity of 10,000 tons/year for 2020 but ceased production of P(3HB) in mid-2015. Table 1 shows the main data related to the industrial production of P(3HB).
Global Production of P(3HB)
P(3HB) is similar to polypropylene in that it is insoluble in water; offers equivalent high-melting temperature and shear stress; and has high crystallinity. However, it has less strength at rupture, 30 which results in a brittle material due to its low thermal stability and rigidity. 9,15,21,31 P(3HB) has the ability to hydrolytically degrade, which distinguishes it from the majority of biodegradable polymers that are currently available because they are typically sensitive to light or moisture or are water soluble. It has excellent solvent resistance and exhibits a high resistance to oils, fats, and UV rays, yet exhibits low resistance to acids and alkalis. In comparison to polypropylene, P(3HB) has a higher density and lower oxygen permeability, resulting in an oxygen-sensitive material that is suitable for use in packaging. 28
Microorganisms Producing PHAs
Positive and Gram-negative bacteria have the ability to accumulate PHAs and can be found in several sources in nature including soil, sea water, and wastewater. 10 More than 300 microorganisms capable of synthesizing PHAs have been listed in the literature; however, only a small number are currently used in the development of PHAs, including Ralstonia sp., Azotobacter vinelandii, Alcaligenes latus, Cupriavidus native and recombinant necator, recombinant Escherichia coli, Alcaligenes eutrophus extroquens Protomonas, Chromobacterium violaceum, 15 and Pseudomonas sp. 32 Under normal growth conditions, most bacteria build up a small percentage of PHA (1–15%); however, under special growth conditions and fermentation strategies, the PHA percentage can reach almost 90% of its dried cell mass. 23
As such, the selection of microorganisms for the industrial production of PHAs is based on several factors, such as high-speed growth, the use of different substrates, the relative cost, the ability to accumulate a large amount of polymer (at least 60% of the dry cell mass, DCW), and the high conversion factor of the substrate biopolymer. 15,32 To be viable for industrial production, it is important that the strains of microorganisms that are employed in the process have optimum characteristics associated with production; for example, rapid growth in a low-cost carbon source, a high percentage of PHA efficiency in the transformation of substrate to product, and efficient adaptation and extraction of the polymer into bacterial cells. They also need to be safe for both humans and the environment. Furthermore, the bacterial strain used to produce the polymer should have a low oxygen demand. 33
The microorganisms used for the production of PHAs can be divided into two groups. The first group is characterized by polymer production in the presence of excessive carbon; growth is stopped or reduced by a lack of essential nutrients such as N, P, Mg, K, S, or O. 15 The second accumulates significant amounts of PHA, even during the cell growth phase. 15,32 Thus, both the development and growth strategy are dependent on the type of microorganism studied. The bacteria that belong to the first group typically perform better in a batch fermentation system divided into two stages, while the microorganisms from the second group yield better results when they are produced in a continuous feed system. 34 Table 2 shows the different PHA production levels by microorganisms.
PHA Yield by Different Microorganisms
Among the wide variety of biopolymer known PHAs, P(3HB) is widely distributed. Poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHB-HV) and poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) (PHB-HHX), are also among the most industrially produced and exploited PHAs at commercial scale. 33
Production of P(3HB)
The synthesis of P(3HB) commences with the non-limited growth phase, during which cell growth occurs. Following that, the polymer is then produced and accumulated; however, this commonly takes place in the limited presence of essential nutrients such as P, Fe, Mg, and N, with excess carbon. 23 Under these conditions, the microorganism rapidly grows and multiplies until consumption of the initial total carbon and the depletion of essential nutrients. In the absence of these, cell growth is limited, and the carbon is converted to PHA. 35 Figure 1 presents a flowchart of the P(3HB) production process. Using a pure culture and incubating in an initial volume of medium (V1), the pre-inoculum is obtained. After a certain time, the V1 increases to a larger volume (V2), and this process continues until the V5 volume is reached, which corresponds to the final inoculum. The produced inoculum is then transferred to a bioreactor containing compressed air, and the culture medium is sterilized to initiate the bioplastic manufacturing process. During the polymer recovery step, the culture medium undergoes centrifugation or filtration, and the pellet, in turn, is extracted using an organic solvent. Finally, the cell debris is discarded, and the organic phase is evaporated, resulting in P(3HB). After its formation, P(3HB) can be used in a variety of materials including pharmaceutical capsules, plastic objects, scaffolds, sutures, and dental prostheses.
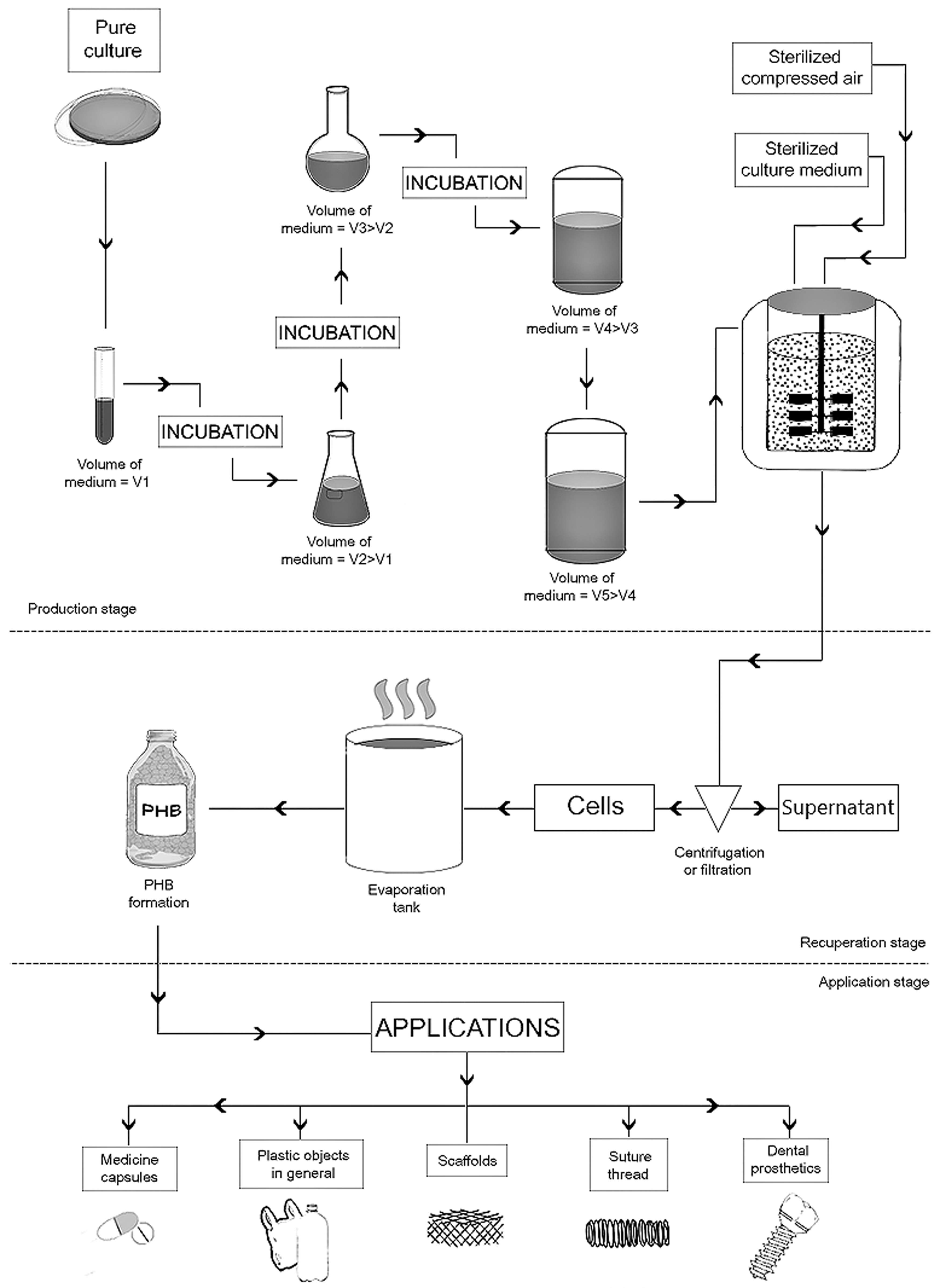
P(3HB) production process, with phases of production, recovery, and applicability.
Bacterial Metabolism
Microorganisms are capable of producing PHA from monomers through the use of various carbon sources. Many pathways have been proposed to produce such monomers. The biosynthetic pathways used to obtain PHA can be based on the monomeric composition. The first group is related to the synthesis of PHAscl, and the second group is represented by the synthesis of PHAmcl. Fig. 2 shows the metabolic pathways for the synthesis of PHA.
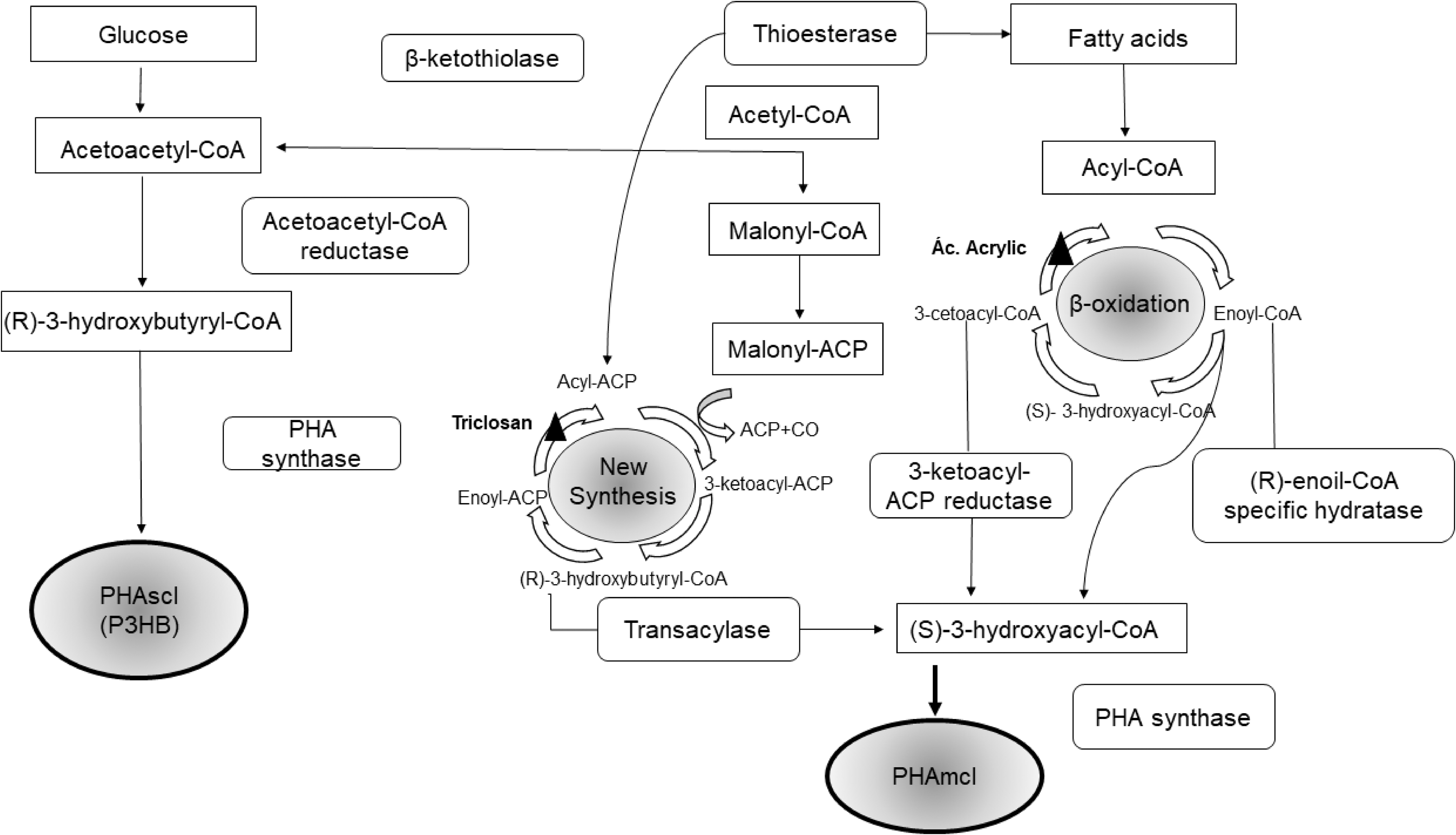
Metabolic routes for the synthesis of PHA.
The synthesis of PHA requires three metabolic phases. In the first phase, a carbon source enables synthesis via a local transmission system through the cytoplasmic membrane of the cell or via the diffusion of the compound into the cell. In the second phase, anabolic or catabolic reactions, or even both, convert the compound into hydroxyacyl coenzyme A, a thioester, which serves as a substrate for PHA synthase, also called PHA polymerase. In the third phase, PHA synthase, which is the key enzyme for PHA synthesis, catalyzes the formation of ester linkages with the release of coenzyme A. 36
The metabolism of the microorganisms is governed by the medium in which they are grown. With balanced growth, the microorganism uses the substrate as an energy source and/or for the maintenance/formation of cellular material. However, when there are both limited essential nutrients and excess carbon in the medium, the carbon substrate is used to form biopolymers that can be stored as energy reserves in the cell. 37
The PHB polymerase is considered the key enzyme for the accumulation of P(3HB) because it performs catalysis, which is the main step involved in the pathway; i.e., the polymerization of D(-)-3-hydroxyacyl-CoA to form the polymer. This enzyme has four classifications that differ according to the substrate specificity and composition of the enzymatic subunit. 9
One of the intermediates for the synthesis of P(3HB) is acetyl-CoA, which is required for the synthesis of the monomer (R)-3-hydroxybutyryl coenzyme A (3HB-CoA). However, this molecule is also a key component for cellular metabolism because it is oxidized by tricarboxylic cycle (TCA), also known as the citric acid cycle or Krebs cycle. Acetyl-CoA—the central intermediary metabolite of all carbon compounds—can be used to generate biologically useful energy or for growth and multiplication. 38 Figure 3 presents a schematic of the possible metabolic routes by which P(3HB) and the biomass, by glycerol or glucose, can be obtained. For glucose metabolism, it is possible to use the Embden-Meyerhof-Parnas (EMP) pathway, also known as glycolysis or the glycolytic pathway, and the Entner-Doudoroff pathway (EDP). The pentose phosphate pathway is another route that can be utilized. 39
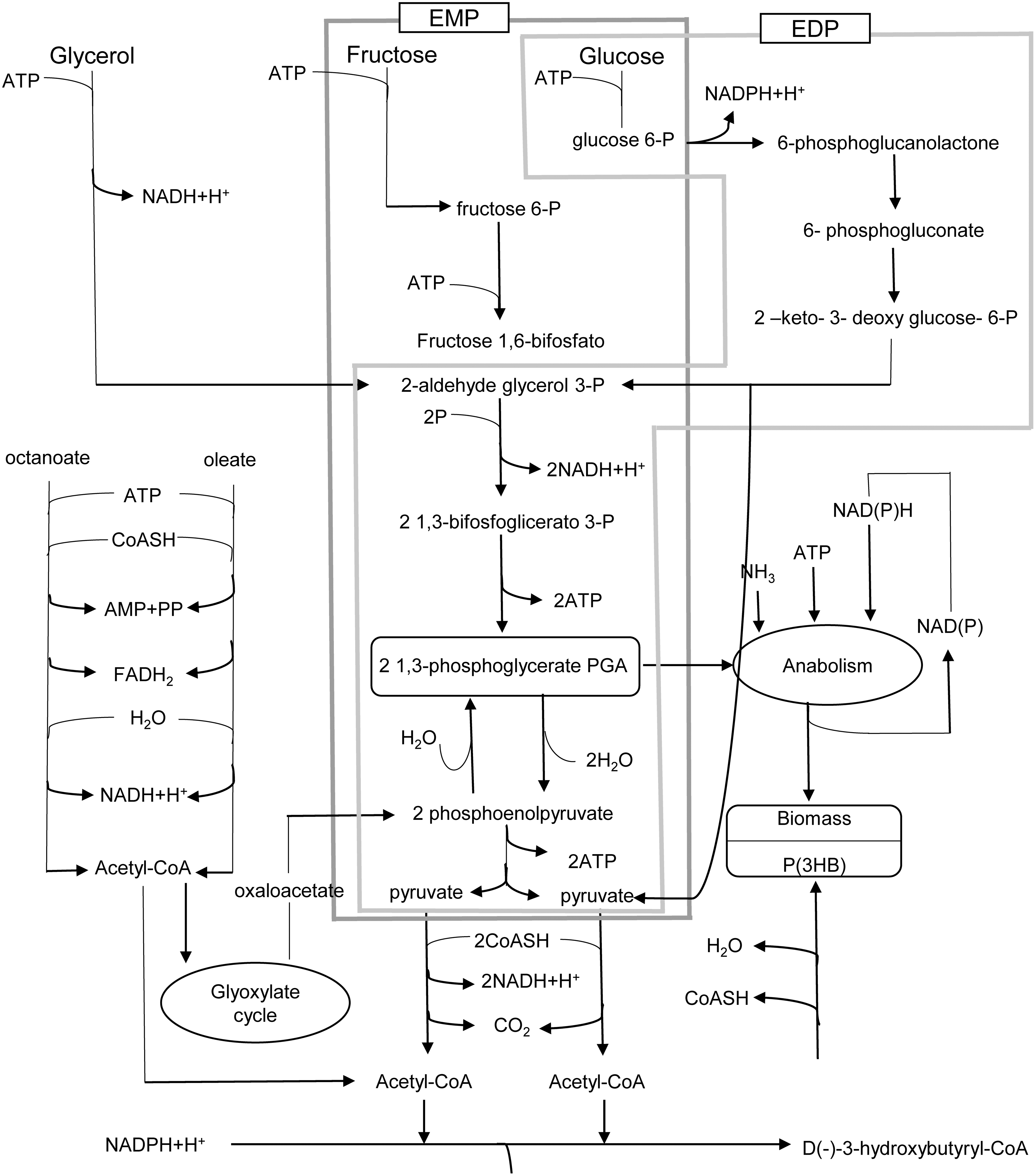
The main metabolic pathways of residual biomass synthesis and P(3HB) from different carbon sources. EMP: Embden-Meyerholf-Parns; EDP: Entner-Doudoroff Pathway. Color images available online at
The synthesis of P(3HB) is favored when there is a high concentration of Acetyl-CoA and NADPH and a low concentration of free Coenzyme A. 25 Tyo et al. showed that, for recombinant E. coli, the enzyme acetoacetyl-coenzyme A reductase NADPH-dependent (Phab) is responsible for controlling the metabolic pathway that produces P(3HB). 39 The NADPH cofactor that is consumed in the reaction of the condensation of 2 acetyl-CoA needs to be regenerated in some other metabolic pathway for the synthesis of P(3HB) to continue. 40
Medium Cultivation
Culture medium containing minerals (mineral resources) is most commonly used when it is intended for the accumulation of P(3HB) in the bacterial cytoplasm. The medium primarily comprises citric acid salts (0.1 to 1.7 g/L), calcium chloride (0.01 to 0.02 g/L) and sodium (0.8 g/L), potassium phosphate (1.5 to 5.5 g/L) and sodium (0.6 to 9.0 g/L), ammonium sulfate (1.0 to 5.0 g/L), and magnesium (0.2 to 1 2 g/L), as well as a solution of trace elements or trace elements. The solution of trace elements may contain boric acid, manganese chloride and copper sulphates, iron, nickel, and zinc. 41 –47 In addition to these nutrients, a source of carbon and nitrogen, which can widely vary, must be supplied to the microorganism.
The fed-batch fermentation approach is considered the most effective in achieving high-cell-density cultivation. 48 By using the appropriate substrate feeding technique, the concentrations of key nutrients (generally the carbon source) can be maintained at an optimum level for a high yield and productivity. 49,50
The carbon source varies in relation to the microorganism's needs. In terms of the absorption capacity, it can be a simple sugar, such as a monosaccharide, e.g., glucose or fructose, 42 which can be assimilated by the EDP and EMP pathways, respectively; or disaccharides, such as sucrose, 45 the assimilation of which depends on the sucrose produced by the microorganism or the prehydrolysis of these complex sugars, such as starch and pectins, as a good source of sugars.
However, many bacteria do not produce the necessary hydrolase decomposition commonly required for the hydrolysis step, such that simple sugars are released that are readily metabolizable via acidification or enzymatic methods. 51 Certain microorganisms do not produce amylase or are not capable of using starch as the carbon source. Therefore, prior to the biotechnological process, starch to be hydrolyzed to glucose must be present. 52 Barbosa et al. 42 used fructose (5, 10 and 15 g/L) as a carbon source to obtain better development and growth of the Ralstonia eutropha ATCC 17697 native strain. To do this, different concentrations of fructose were employed in a fed batch bioreactor. When the microorganism depleted the sugar available, the researchers added the sugar equivalent to the initial concentration of the culture medium. For all concentrations tested, 7.5 g of dry biomass/L was obtained; in terms of the accumulation of P(3HB), the best results for the 5 and 15 g/L concentrations were 66 and 57% respectively; and for 10 g/L, an accumulation of 44% was reached. Patwardhan and Srivastava 53 conducted a P(3HB) production study using a Wautersia eutropha strain using fructose as the carbon source and obtained a higher DCW of 14 g/L—with a polymer yield of only 6%—than Barbosa et al. 42 previously cited.
Using a Halomonas TD01 strain with glucose as the carbon source, Tan et al. 54 obtained a cell biomass of 40 g/L and a polymer yield of 60%. Pradella and collaborators 55 used sucrose (700 g/L) in the production of P(3HB) by Burkholderia sacchari TPI 189 and obtained a dry cell biomass of 150 g/L and a polymer yield of 42%. Borah et al. 56 evaluated three types of carbon sources—sucrose, glucose, and fructose—with Bacillus mycoides RLJ B-17 and obtained DCW yields of 3.6 g/L, 3.2 g /L, and 2 g/L, and polymer yields of 69.4%, 56.6%, and 55%, respectively.
Nevertheless, different microorganisms have different requirements and/or tolerance in relation to sugar concentration. Stockdale et al. evaluated the production of P(3HB) by microorganisms from the Azotobacteriaceae family using 1% glucose. 57 They concluded that the strains with the best production were A. beijerinckii NCIB 9067 (70.4%), A. agilis NCIB 8637 (43.3%), and A. chroococcum NCIB 8002 (40.3%). The same authors also used glucose at a concentration of 2%, and the best results were obtained with A. beijerinckii NCIB 9067 (67.7%), A. vinelandii NCIB 8789 (50.3%), and A. agili NCIB 8637 (48.6%).
The carbon source may be responsible for 25–45% of the total production costs. 58 Thus, there is a need to identify lower-cost sources of carbon to decrease the cost of PHA production. The worldwide abundance of available biomass originating from renewable carbon can be used as raw material for production. Many of the wastes and byproducts of agricultural and dairy industries represent attractive candidates because they possess the main characteristics desired, such as low cost and high availability. 52 A wide variety of substrates, such as molasses, 59,60 glycerol, 46,47,61 whey, 58 rice starch, 62 and vegetable oils, 63 have been used to produce PHA from various microorganisms.
Among the agricultural byproducts mentioned, molasses has been widely used in various commercial fermentation processes because it contain vitamins and nutrients that help promote growth and microbial metabolism. 60 Kulpreecha et al. compared the same concentration of molasses (20 g/L) and sucrose as carbon sources for Bacillus megaterium BA-019. 64 The dried biomass and yield of P(3HB) were significantly improved when molasses, instead of sucrose, was used as a carbon source, regardless of the nitrogen source. At 24 h, values of 7.05 g/L and 55.46% to DCW and P(3HB) accumulation were obtained, respectively, against 2.83 g/L and 30.20% when sugar was used in the same concentration. This result is of great significance because molasses represents a less expensive carbon source that can be readily assimilated. Kanjanachumpol et al., on the other hand, evaluated the influence of different concentrations of sugarcane molasses (46°Brix; 20, 40, 60 and 80 g/L) as a carbon source in a 10-L bioreactor with an initial sugar concentration of 60 g/L. 65 The highest result was obtained for the DCW and P(3HB) accumulation, 32.5 and 8.8 g/L respectively, during just 12 hours of production, proving the effectiveness of using molasses sugarcane.
The effect observed in molasses may be caused by the presence of other nutrients in its composition. In addition to glucose, molasses contains fructose and sucrose, organic acids, minerals, and vitamins such as thiamine, riboflavin, and pyridoxine, which function as growth factors and thereby stimulate cell growth and the production of P(3HB). 64
Glycerol, which is an important agricultural byproduct, is the main residue of biodiesel production, representing approximately 35% of the output. Although pure glycerol is an important industrial raw material that has applications in the food, medicine, cosmetics, and tobacco industries, crude glycerol has a relatively low value due to the presence of impurities. For this byproduct to be truly recycled, it must undergo several purification steps to remove impurities, and these expensive phases are often not economically viable for small- and medium-sized plants. 66 Biological conversions offer an opportunity for glycerol to be used in the synthesis of various products with different features. Glycerol may be used as a carbon source instead of sugars in many microbiological processes. PHA production is an attractive alternative for the biological processing of glycerol. Mothes et al. obtained a P(3HB) polymer yield of 48% and DCW of 50 g/L using glycerol at a concentration of 10 g/L with the microorganism Cupriavidus necator JMP 134 DSMZ 4058. 67 Cavalheiro et al. explored two strategies for improving the production of PHAs: the use of high-density cultures and glycerol residues (GRP) as the primary carbon source for the growth of Cupriavidus necator DSM 545 and synthesis polymers. 61 In this study, the authors evaluated the production of P(3HB) from commercial glycerol (PG), a substrate control, and GRP. Using PG, the accumulated P(3HB) was 62%, with a DCW yield of 82.5 g/L, compared with 50% P(3HB) and dried cell mass of 68.8 g/L using GRP.
Naranjo et al. compared glucose and glycerol as carbon sources and too found a higher accumulation of P(3HB) in Bacillus megaterium when using glycerol (62%); when using glucose, the accumulation was 59% under the same growing conditions. 46 Rodriguez-Contreras et al. considered the same two carbon sources but switched the bioreactor to glucose and glycerol, obtaining good results, with 65% and 68% of P(3HB) concentration and biomass respectively, for Cupriavidus necator, DSM 545. 47 The carbon source used in the study for the production of P(3HB) from strain Zobellella denitrificans MW1 was glycerol, which resulted in a dry biomass of 81.2 g/L and a polymer accumulation of 66.9%. 64 Bormann et al. evaluated the production of the polymer using Ralstonia eutropha DSM 11348 with glycerol and casein hydrolysates and obtained DCW of 32 g/L and a polymer yield of 15.4 g/L (47%). 68
In terms of starch and its derivatives, sources of carbon are available in large quantities in Brazil. They are the main components of the biochemical oxygen demand of the textile industry wastewater, fermentation, and processing of food and beverages. 69 In the food industry, starch may be obtained from grains, such as rice, wheat, and corn, and from tubers such as potatoes and tapioca. 70 Rusendi and Sheppard used the residue that was generated from processing industry potatoes for P(3HB) production and, through enzymatic hydrolysis, obtained glucose as a carbon source. 71 Dalcanton et al. considered the hydrolyzed rice starch cultivation with nutrients added to the mineral medium. 62 Furthermore, they supplemented the medium with soybean oil. They concluded that the use of hydrolyzed rice starch was a viable alternative to the production of P(3HB). It is also worth noting that the hydrolyzed rice contained satisfactory concentrations of metabolizable carbon sources, comprising approximately 10 to 15% proteins. When hydrolyzed, this served as a source of nitrogen and phosphorus to the microorganism, as well as providing a small percentage of lipids, fiber, and minerals (approximately 1%). Supplementation with soy oil, even in small quantities, led to an increase in the production of P(3HB) (43%) compared to media without supplementation (35%). Hafuka et al. studied the production of P(3HB) from Cupriavidus necator H16 (ATCC17699) in continuous fermentation that used liquid food wastes as a carbon source and obtained a polymer yield of 87%. 72 Pandian et al. also used alternative sources, such as a residue from dairy industrialization (150–550 mL/L), rice bran (20–60 g/L), and seawater (150–550 mL/L), to produce the polymer using B. megaterium SRKP-3, and obtained a yield as high as 11.32 g/L of DCW. 73 Yezza et al. produced P(3HB) from Alcaligenes latus using maple sap and obtained 4.2 g of dry biomass/L and a 77% polymer yield. 74
Quillaguaman et al. compared the production of P(3HB) in an orbital shaker incubator to that in a bioreactor using a Halomonas boliviensis strain with a starch hydrolysate that resulted from bacterial recombinant enzymes. 75 They obtained a better dry cell biomass yield (56%) in the shaker than they did in the bioreactor, with the highest yield in the fermentor being 35%. Starch was also used to enhance cell growth with Azotobacter chroococcum 23 in a shaker batch study by Kim and Chang, in which they obtained 8.4 g/L of DCW and 3.7 g/L of P(3HB) with a polymer yield of 44%. 76 In the fermentor, they obtained 54 g/L of DCW and 25 g/L of P(3HB) with a polymer yield of 46%.
Ramadas et al. evaluated various carbon sources (wheat bran, potato starch, sesame oil cake, peanut oil cake, ground tapioca, jackfruit seed powder and corn flour) in the cultivation of Bacillus sphaericus 5149. 44 The culture medium containing wheat bran as a substrate was more beneficial for cell growth, resulting in 15.5 g/L of biomass; however, for the accumulation of P(3HB), the use of jackfruit seed powder and potato starch resulted in higher yields of 46 and 47%, respectively.
Crop supplementation studies have found that the addition of oleic and linoleic acids leads to an increase in the production of PHAs. 77 These fatty acids are part of the composition of many vegetable oils, thus, making them a possible low-cost alternative to supplementation crops. Grigull et al. evaluated various oleic acid concentrations (0 to 3.0 g/L) in the culture medium of minerals, linking glucose and fructose as carbon sources for the Ralstonia eutropha strain DSM 545. 43 They concluded that oleic acid was consumed in the exponential phase of growth and that the concentration of this food supplement resulted in an increase in the bacterial growth rate. The yield obtained was 6.96 g/L of biomass with supplementation of 3.0 g/L oleic acid, and a higher accumulation of P(3HB) (16.7%) was obtained at the same concentration. Ng et al. used jatropha oil from India (20 g/L + 10 g/L) as a carbon source for the production of polymer using Cupriavidus necator H16 and varied the nitrogen source—NH4Cl, NH4H2PO4, NH4NO3, (NH4)2SO4, NaNO3 and (NH2)2CO. 78 The best biomass results (8.2 g/L) with a 75.6% polymer yield were obtained using (NH2)2CO, and a better polymer yield (87.9%) with a biomass of 3.3 g/L was obtained using NH4H2PO4. Sangkharak and colleagues used acetate as the carbon source for three strains of Rhodobacter sphaeroides, and, with two of these mutants, they obtained higher polymer yields of 72.94% and 50.03%, compared to only 22.21% using the natural strain. 79 Kim et al. supplemented the culture medium with methanol to produce bioplastics from Methylobacterium organophilum NCIB 11278. 80 They obtained 185 g/L (52%) in 57 h of production and 130 g/L (56%) in 70 hours, highlighting the need to determine the optimum time to complete the process, and, thus, avoid self-degradation of the polymer.
Urea is the best source of nitrogen due to the superior assimilating capacity of B. megaterium BA-019 compared to other nitrogenous compounds. Urea is a small uncharged polar molecule; in contrast to ammonium sulfate, the urea uptake rate through the cell membrane is less pH-dependent and faster. Indeed, the use of this cheap source of nitrogen can reduce the cost of P(3HB) production because the bacterial strain employed can utilize it efficiently. 77 Kulpreecha et al. also assessed urea and ammonia and sulfate (0.8 g/L) as nitrogen sources for B. megaterium BA-019. 64 With respect to the nitrogen source, higher biomass yields (7.05 g/L) and P(3HB) (55.46%) were obtained through the use of urea. They obtained high accumulations of polymer but at a reduced DCW content. The best results were obtained using casamino acids at 3.8 g/L (81.6%).
Sometimes, however, unsatisfactory results are obtained. Koller et al. evaluated the production of PHA by Pseudomonas hydrogenovora DSM 1749 using a whey milk hydrolysate (rich in glucose and galactose) as a carbon source and obtained a low DCW of 1.44 g/L, and 1.44 g/L of P(3HB) and 0.3 g/L of P(3HB-co-3-HV), corresponding to a total yield of 12%. 81
The results obtained by the above-mentioned studies, in terms of the various carbon and nitrogen sources employed, are presented in Table 3.
Dry Cell Mass Yield and Accumulation of P(3HB) Obtained in Cultures of Different Microorganisms, Carbon and Nitrogen Sources
Not included
Growing Conditions: Time, Temperature, pH, and Oxygen
In general, the optimization of fermentation conditions has proven to substantially improve the yield and productivity of many biological processes. 84 Because the synthesis of P(3HB) is known to be influenced by environmental stresses, such as essential nutrients like nitrogen, phosphate or oxygen, 7 fermentation optimization has been widely used. In addition, factors that influence the process by which a substrate is converted into a product, such as time, temperature, and pH, have also been optimized.
The main difference between the fermentative processes in a shaker and a bioreactor is that the oxygen supply and the agitation speed of a shaker cannot be used to control the supply or that the dissolved oxygen (DO) rate and agitation speeds are lower. Most microbial industrial processes are aerobic and conducted mainly in media that contain salts and organic substances. In these processes, oxygen is an important nutrient that affects process performance because the microorganisms use it in the growth, maintenance, and production of metabolites. As such, a lack of oxygen affects process performance. 82 –84 Therefore, it is important to ensure there is an adequate supply of oxygen from the gas stream into the culture medium. Accordingly, the oxygen transfer rate (OTR) varies according to the operating conditions and plays an important role in predicting the metabolic pathway for the growth and production of any metabolite in the culture. Thus, when the DO supply is so low that it is limited and induced stress is required, for example, in B. megaterium, which is strictly aerobic, fermentation is required in the bioreactor to achieve higher P(3HB) production levels by controlling the low DO levels. 85
Kulpreecha et al. evaluated the effect of different values of pH (6.0, 7.0, and 8.0) and DO concentration (40, 60, and 80% saturation) in the culture medium on the fermentation of the B. megaterium BA-019 batch. 64 For the biomass yield, the best result (9.7 g/L) was obtained in fermentation without pH control and with a saturated DO level of 60%; however, the results were not significantly different from those obtained via a combination of pH 8.0 and 80% saturation (9.5 g/L). In contrast to the yield of P(3HB) that accumulated in the cell, a combination of pH 7.0 and 60% saturation resulted in the accumulation of 61.6% the DCW. This may be because pH 7.0 is the optimum value for the key enzymes in the biosynthetic pathway of P(3HB) for that microorganism. Therefore, in the presence of a pH that is higher or lower than the ideal value, the enzymatic activity decreases. It was reported that pH changes can reduce the performance of the culture and that the initial pH can affect the availability of the trace elements in culture medium that are required for bacterial growth. 86
Borah et al. used the three types of sources described above in Bacillus mycoides RLJ B-17 cultures for production in a fermenter using a 20% inoculum (30°C, 650 rpm, 1 vvm, 24 h, pH fixed at 7); they obtained polymer yields of 69.4%, 56.6%, and 55%. 54 Ramadas et al. studied different agro-industrial waste as a source of carbon for the cultivation of the Bacillus sphaericus strain 5149, but they tested different pH values (5.0 to 8.0). 44 They found that an initial pH change in the cultivation medium had a strong influence on the production of P(3HB). Even a small difference in pH from the ideal point caused a sharp reduction in the accumulation. In this work, the medium with an initial pH of 7.5 resulted in the maximum yield of P(3HB) of 25%. The drastic change in production appeared to be due to the effect of the initial pH on the bioavailability of trace elements. Pandian et al. used the nutritional sources described above with Bacillus megaterium SRKP-3 at pH 9 and 30°C, with an agitation of 300 rpm and aeration of 100% at the beginning of the process. 73 They obtained 11.32 g/L of dry cellular biomass; when pH is related to the production of P(3HB) by genera Bacillus, it should be noted that using different pH values resulted in different polymer accumulation.
Ibrahim and Steinbüchel used glycerol to produce P(3HB) by Zobellella denitrificans MW1 at pH 7.3 and 37°C for 24 h with high aeration and stirring (1.67 vvm and 800 rpm) in the fermenter. 87 They obtained 81.2 ± 2.5 g/L of DCW and a polymer concentration of 66.9% ± 7.6%. Bormann et al., as described above, performed fermentation using Ralstonia eutropha DSM 11348 at pH 6.8–7.0, 30°C, with high agitation (800–1000 rpm) for 48 and 92 h, after inoculating the fermenter and following the OD at 420 nm. 68 They obtained 32 g/L of DCW and a polymer yield of 15.4 g /L (47%).
In terms of the OD, at a saturation level of 40%, the productivity decreased dramatically due to the insufficient amount of oxygen available in the culture medium to meet the aerobic anabolic requirements. However, higher OD levels (80%) significantly reduced bacterial growth, probably due to the oxidative stress and shear that resulted from the high stirring speed. 61 Therefore, it would be interesting to maintain a constant stirring rate and adjust the oxygen levels via the addition of pure oxygen-enriched air to prevent stress in the bacterial cell.
Faccin et al. evaluated oxygen transfer while subjecting the B. megaterium DSM 32 culture to different agitation levels (100, 200, 300, 400, 500, and 600 rpm). 45 The highest yield of P(3HB), 62%, was obtained by shaking at 200 rpm to generate an oxygen transfer coefficient (kLa) of 0.006 s 1 . The (kL) is an important parameter because it is related to the OTR and is commonly used when an increase in the production scale is desired. 88
Martínez et al. evaluated the effect of pH (6.0, 6.8, and 7.4), agitation (300 and 700 rpm) and aeration (0.25 and 1 vvm) on the microorganism Azospirillum brasilense 7 strain, using tomato extract as a carbon source. 41 A maximum biomass yield of 1.67 g/L and a maximum P(3HB) yield of 0.16 g/L at pH 6.8 were obtained. Agitation of 500 rpm and an aeration value of 0.5 vvm was the best combination, yielding a biomass of 1.2 g/L; however, for an increased yield of P(3HB) (4.9 g/L), the culture was maintained at a greater agitation (700 rpm) and lower aeration (0.25 vvm).
Dalcanton et al. evaluated the effect of different temperatures (30, 35, and 40°C) on the growing culture of Cupriavidus necator DSM 545. 62 Holding the growth temperature at 35C was advantageous for growth (12.4 g/L) and P(3HB) production (43%), compared to those performed at the temperatures of 30°C and 40°C, the former of which is currently used in the production process. Both the enzymes present in this organism and others show the maximum activity when factors such as temperature are close to optimal, which may lead to an increased specific growth rate and, hence, favorable changes in the conversion factor of the substrate product. This increased cultivation temperature may still have a significant influence on the productivity of the process due to the reduction in cultivation time. However, according to Schimdell et al., very high growth temperatures decrease the concentration of oxygen dissolved in the medium, which is an important factor when the P(3HB) production process is aerobic. 88 In this context, studying the influence of culture temperature on the production of P(3HB) is extremely important because the temperature increase still implies a reduced need for cooling during the process.
Crochemore et al. reports that the accumulation of lipophilic inclusions is dependent on the cultivation time for the studied microorganism, Pseudomonas strain CMM43. 32 The cultivation was performed in flasks incubated on a shaker at 28°C and a speed of 150 rpm over periods of 24, 48, and 72 h. The greatest accumulation of P(3HB) was observed at 48 h (29%), with a cell mass yield of 5.8 g/L, while the lower accumulation was observed at 24 h of culture. This latter finding was expected because the production process and accumulation of P(3HB) are usually slow processes. 89 At 72 h of cultivation, the accumulation of the polymer decreased by approximately 4% compared to that obtained after 48 h. It was also found that sugar consumption continued after 48 h of culture, which indicated that the bacteria used the polymer as a source of carbon.
Ng et al. used the conditions specified above for the growth of Cupriavidus necator H16 at pH 7, 30°C, 200 rpm, 6–120 h and obtained biomass that ranged from 3.3 g/L to 8.2 g/L and a polymer yield of 2.9 to 6.5 g/L. 78 Studying the production of P(3HB) using liquid food waste as a nutritional source for Cupriavidus necator H16 (ATCC17699) and using two fermentation regimes for 259 h, Hafuka et al. obtained a high polymer yield—87%—and a productivity of 5 mg/L/h in continuous fermentation. 72 During the production of P(3HB) from Alcaligenes latus using maple sap with a 5% inoculum, pH 7.9, and an aeration of 30% in the fermenter for 33 h, Yezza et al. obtained a low DCW of 4.2 g/L, but a good yield of 77%. 74
Quillaguaman et al. used the Halomonas boliviensis strain in combination with the conditions described above to study the production of the polymer in 1-L shaker flasks with the pH adjusted to 8.5 at the beginning, with an aeration of 310 mL and shaking at 160 rpm. 75 They obtained a DCW of 56%. In a fermenter at pH 8, with an aeration of 0.53 vvm and an agitation rate of 700 rpm, the percentage obtained was lower—35%—probably due to high agitation. At the same time, Tan et al., using Halomonas TD01 with glucose as the carbon source in a shaker flask (pH 9, 200 rpm, 37°C for 48 h) obtained 40 g/L of DCW and a polymer yield of 60%. 54 These values are higher than those obtained by Quillaguaman et al. 75
Using starch to promote the growth of Azotobacter chroococcum 23, Kim and Chang used shakers in batch mode at a pH of 7.2 in the beginning, at 30°C with stirring at 300 rpm for 0–40 h, and 1 vvm for 12 hours, and obtained 8.4 g/L of DCW, 3.7 g/L of P(3HB), and a polymer yield of 44%. 76 In a fed-batch fermenter with a stirring rate of 1,000 rpm and O2 limitation for 37 h, at 70 h 54 g/L of DCW, 25 g/L of P(3HB), and a polymer yield of 46% were achieved.
Patwardhana and Srivastava performed a P(3HB) production study in fermenters using the Wautersia eutropha strain in combination with fructose as the carbon source at a controlled pH for 60 h with 30% aeration, and obtained 14 g/L of DCW but a very low polymer yield of 6%. 53 Koller et al. evaluated the production of PHA by Pseudomonas hydrogenovora DSM 1749 with glucose/galactose whey from hydrolyzed milk as a carbon source with an inoculum stage—pH 6.8–7 and 30°C for 48 h—and production phase—pH 7 and 30°C for 48 h—and obtained a very low DCW of 1.44 g/L and 1.14 g/L of P(3HB) with a total yield of 12%. 81
Using acetate as the carbon source, Sangkharak et al., as described above, used three strains of Rhodobacter sphaeroides, of which two were mutants, for the fermenter production of P(3HB) at pH 7, an agitation rate of 150 rpm for 60 h. 79 The result was a polymer yield of 21% for the natural strain 22, and of 72.94 and 50.03% for the mutant strains.
Kim et al., using the conditions described above with Methylobacterium organophilum NCIB 11278, pH 7, 1,400 rpm, and 30°C for both the inoculum and production phases, obtained 185 g/L (52%) in 57 h of production, and 130 g/L (56%) in 70 h. 76
The results obtained in this work, and others not mentioned, including the operating conditions, are shown in Table 4.
(DCW) Yield and Accumulation of P(3HB) Obtained in Cultures of Different Microorganisms Under Different Conditions in a Shaker and a Bioreactor
Not controlled; bwithout control; ccontrol.
Conclusions
P(3HB) is a lipophilic bacterial polymer whose applicability puts it in a prominent position among other biodegradable polymers from sustainable sources. Factors that should be considered to obtain a higher accumulation and polymer yield, together with improving its quality, are dependent on the producing microorganism. For economically viable production, the adequacy of the means and process operating parameters, including the use of waste, is crucial. Although Brazil is one of the largest producers of sugarcane and biodiesel, and thus generates a large volume of molasses and glycerol—excellent sources of carbon in alternative culture media—there is currently no P(3HB) production in the country.
In this review, we observed extremely variable polymeric yields; unfortunately, some authors have reported high biomass concentrations associated with low yields of P(3HB). This may be related to the low accumulation of polymer bead consumption by the organism due to unfavorable operating parameters or an inefficient extraction method.
Footnotes
Author Disclosure Statement
No competing financial interests exist.